山西長治濕地草本植物優勢種群和群落的空間格局分析
2011-03-28 15:11:40李素清武冬梅王濤上官鐵梁
草業學報 2011年3期
李素清,武冬梅,王濤,上官鐵梁,3
(1.山西大學黃土高原研究所,山西 太原 030006;2.中國科學院地理科學與資源研究所,北京100101;3.山西大學環境與資源學院,山西 太原030006)
植物種群和群落空間格局分析是現代植物生態學的研究熱點之一。自1952年Greig-Smith首次使用方差分析方法研究植物種的格局規模以來,格局分析受到許多生態學家的重視,現已成為研究植物種類空間關系、種類與環境關系的重要手段[1]。在過去的50多年中,許多新的格局分析方法陸續出現,如最近鄰體分析法(nearest neighbor analysis,NAA)[2]、雙項軌跡方差分析法(two-term local quadrat variance,T TLQV)[3]、隨機配對法(random pairing)[4]、譜分析法(spectral analysis)等[5],這些方法現已在種群和群落格局研究中被廣泛接受和應用,促使格局分析理論與方法取得了很大進展。
20世紀80年代以來,我國學者對植物種群分布格局研究較為活躍[6,7],特別是對森林和草地植物群落分布格局的研究報道層出不窮,如張金屯等[8]應用種毗連法和種小區法,對五臺山亞高山草甸小格局的研究表明,在小格局中,大部分種與優勢種苔草(Carex spp.)呈正關聯,小格局規模都較小,各個種趨于交錯分布;不同規模的格局應用不同的分析方法。張金屯[9,10]建立了除趨勢對應分析和TT LQV相結合的二維格局分析方法,并應用該方法對蘆芽山亞高山草甸優勢種群和群落的二維格局進行了研究,結果表明該方法簡單易做,可較好地反映種群和群落的空間特性,草甸主要優勢種格局斑塊的形狀比較規則、面積也較大,群落格局與主要優勢種的格局關系密切。李素清等[11]應用除趨勢對應分析與TT LQV相結合的方法,對云頂山亞高山草甸優勢種群和群落分布格局的研究表明,群落格局與其優勢種的格局關系密切,種群和群落的分布格局不僅受種的生物學特性影響,還受環境因子制約。徐連秀和蒙榮[12]應用方差/均值的T檢驗、聚集指數、平均擁擠度等方法,對內蒙古四子王旗短花針茅(Stipabrevi f lora)荒漠草原植物群落優勢種群分布格局的研究表明,這些植物的分布格局皆屬于聚集分布,但其聚集強度有差異;方差/均值比率法是一種較好的分析種群分布格局的方法。張育新等[13]應用方差/均值比率、叢生指標、負二項式等指標方法,對北京東靈山海拔梯度上遼東櫟(Quercus liaotungensis)種群結構和空間分布的研究表明,東靈山海拔梯度上遼東櫟種群比較穩定,遼東櫟種群結構和空間分布在海拔梯度上的分布特征是種群發育歷史、物種特性、環境、干擾等因素在海拔梯度上綜合作用的結果。魯為華等[14]應用Ripley的K函數對重度和極度退化區域的伊犁絹蒿(Seriphidium transiliense)種群進行了空間格局及關聯分析,表明重度退化區域的絹蒿種群總體上強烈偏離隨機分布呈聚集分布,而極度退化區域內絹蒿種群則在小尺度內呈聚集分布,到達聚集分布的臨界值時則轉入隨機分布。閆海冰等[15]應用點格局分析中的單、雙變量O-ring統計方法,對華北山地典型天然次生林群落的樹種空間分布格局及其關聯性的研究表明,種群分布格局和種群間的空間關聯性隨群落結構、空間尺度和齡級結構不同而有較大變化。李立等[16]應用Ripley的K函數點格局分析方法,對古田山常綠闊葉林優勢樹種甜櫧(Castanopsis eyeri)和木荷(Schima superba)空間格局的研究表明,甜櫧與木荷2種優勢物種在總體上及不同年齡階段主要呈聚集分布,隨年齡階段的增加,其聚集程度有降低的趨勢。這些研究有力地推動了我國森林和草地植物群落空間分布格局研究的發展。但我國對濕地植物群落分布格局的研究報道較少,僅見黃志偉等[17]應用聚集度指標等方法,研究青海湖幾種主要濕地植物的種群分布格局及動態。武玉珍和張峰[18]應用方差/均值的T檢驗等方法,研究山西桑干河流域濕地植被優勢種群分布格局。徐洋等[19]采用幼苗萌發法,研究生態位限制和物種庫限制對湖濱濕地植物群落分布格局的影響。總體看,近30年來我國對濕地植物群落分布格局的研究進展緩慢,目前亟待加強。
濕地作為一種介于水陸之間的獨特生態系統,在生物多樣性保護方面具有極其重要的價值。長治濕地是山西省面積最大和保存最好的水庫濕地,是開展濕地植物物種多樣性研究的理想區域。近年來,一些學者先后開展了長治市濕地主要植被類型調查與研究[20],但對該濕地草本植物優勢種群和群落分布格局的研究尚未見報道。本研究選擇長治濕地典型地段,以草本植物群落為對象,應用除趨勢對應分析(detrended correspondence analysis,DCA)[21-23]與雙項軌跡方差分析[3]相結合的方法,對分布于其西南部海拔900~910 m的濕地草本植物群落空間格局進行分析,揭示其種群分布特征和群落生態特點,以期為該濕地的生態保護和合理利用提供科學依據。
1 材料與方法
1.1 研究區概況
長治濕地是由國家建設部批準設立的山西省第1個國家級濕地公園,位于長治市主城區西北3 km處,屬南運河水系濁漳河南源,面積45.86 km2,東臨長北干線,南至長晉高速下秦村,西通屯留縣境內絳河下游長太高速,北達漳澤湖大壩[24]。濕地屬于暖溫帶大陸性季風氣候,年均溫9.5℃,年降水量621.1 mm,年蒸發量1 070.9 mm,雨量集中在6-9月,無霜期155~184 d。濕地土壤屬于草甸土和沼澤土2個亞類,土壤pH值為8.1,呈微堿性。整個濕地地貌典型完整,景觀獨特幽雅,生態資源豐富。據統計,濕地內有高等植物52科217種,浮游植物7門83種,鳥類16目40科162種,主要水生動物7綱25種,浮游動物58種,有數百公頃蘆葦(Phragmites australis)蕩和200 hm2以上的濕地防護林,是不可多得的重要生態資源,是華北地區少有的野生動植物自然繁育基地,是山西省面積最大和保存最好的濕地生態系統之一。
1.2 研究方法
1.2.1 取樣 采用樣帶法分別在長治濕地西南部與水面平行(海拔889 m)和與水面垂直的方向(906 m)各設1條樣帶(圖1),共2條樣帶。每條樣帶由256個10 cm×10 cm連續的小樣方組成,記錄每個小樣方中所有種的蓋度,用蓋度值作為格局分析的基本數據[8]。
1.2.2 分析方法 格局分析選用雙向軌跡方差法,其計算過程如下:
1)應用除趨勢對應分析法對濕地草本植物群落中所有種的數據進行排序[3],得到256個樣方的DCA排序坐標。分別以小樣方為橫坐標,DCA第1、第2排序坐標值為縱坐標,繪制排序值變化圖。該圖可視為是群落格局變化圖,因為橫坐標實際上代表區組,縱坐標代表變差,圖上峰值所對應的區組即為在該尺度下的格局[10]。這里分別對濕地西南部2個不同部位的草本植物群落DCA第1、第2排序軸進行分析,得到群落格局分析圖,對它們的結果進行比較。
2)以256個樣方的DCA第1排序軸坐標值為依據,采用雙向軌跡方差法分析,以判別群落的格局,其計算區組方差的公式為[9]:

圖1 長治濕地的地理位置及樣帶布置Fig.1 The geographic location and transect layout of Changzhi wetland
式中,V(b)為區組方差,b為區組,j為起點小樣方的序號,xi為小樣方i的觀測值,n為小樣方數。它可以計算1/2n以內任何大小區組的方差。以區組大小為橫坐標,以方差為縱坐標繪制格局分析圖,圖上峰值所對應的區組即為群落在該尺度下的格局。分別對2條樣帶進行分析,就可以得到群落格局斑塊的長度或寬度,也即群落格局的分析結果。
3)采用TT LQV法分別對2條樣帶中的4個主要優勢種進行分析,就可以得到優勢種格局斑塊的長度或寬度,也即群落優勢種格局的分析結果。
全部分析結果用區組-均方圖表示(圖2~5),區組選用區組1,2,4,8,16,32,48,56,64,128和256,這基本能夠反映濕地草本植物群落的分布特征[22]。
1.2.3 群落命名 依據《中國植被》的群落命名原則[25],采用優勢種命名法,結合DCA排序結果,分別將2條樣帶所在的草甸群落命名為扁稈藨草+蘆葦(Scirpusplaniculmis+P.australis)群系和小苜蓿+雙穗雀稗(Medicago minima+Paspalum paspaloides)群系,從樣帶Ⅰ※樣帶Ⅱ,海拔為 889※906 m,坡度為0°※5°,生境由水庫邊緣濕地※與水庫邊緣垂直的平坡濕地,土壤含水量逐漸減少,植物種類由以濕生、中生植物為主轉向以中生、旱中生植物為主[26],放牧等人類活動干擾強度由弱※強,2條樣帶的主要植物種類組成和特征變化如表1所示。
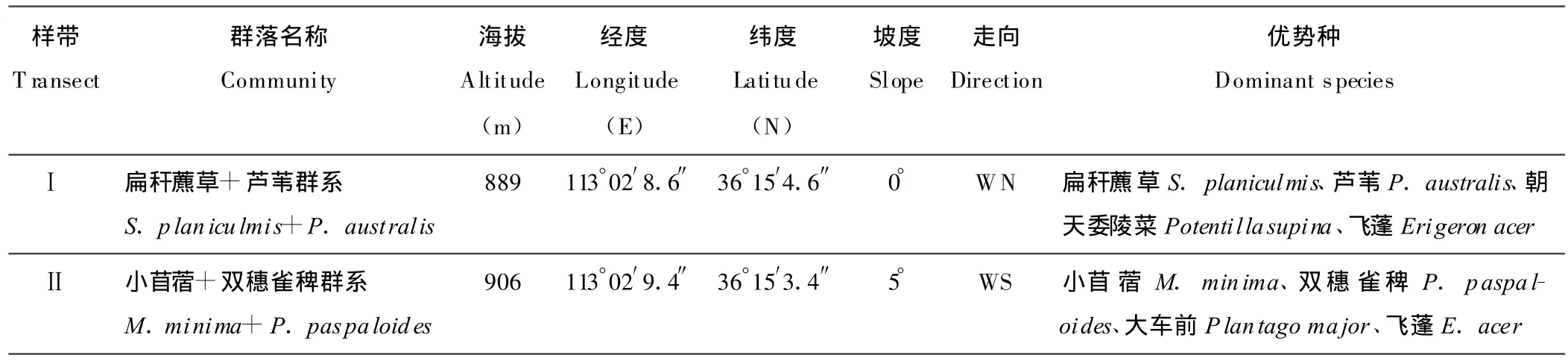
表1 長治濕地2條樣帶概況Table 1 Survey of the two transect of herbaceous communities in Changzhi wetland
2 結果與分析
2.1 優勢種的格局
2條樣帶分別代表2個不同的群落類型,建群種扁稈藨草在區組5和71處有峰值,表明該種的小斑塊和中斑塊的規模分別為50和710 cm(圖2),而大斑塊未能顯示出來;建群種蘆葦在區組6和98處有峰值,表明該種的小斑塊和大斑塊規模分別為60和980 cm,而中斑塊未能顯示出來;優勢種朝天委陵菜的小斑塊規模為20 cm,中斑塊為420 cm,而大斑塊為1 270 cm;優勢種飛蓬的小斑塊,中斑塊和大斑塊規模分別為70,480和900 cm,這可能與朝天委陵菜和飛蓬具有明顯的群集性生長特性有關。與圖6中樣帶Ⅰ的分析結果比較,二者基本一致,可見在群落斑塊的形成中,優勢種扁稈藨草、蘆葦、朝天委陵菜、飛蓬等起著關鍵作用。
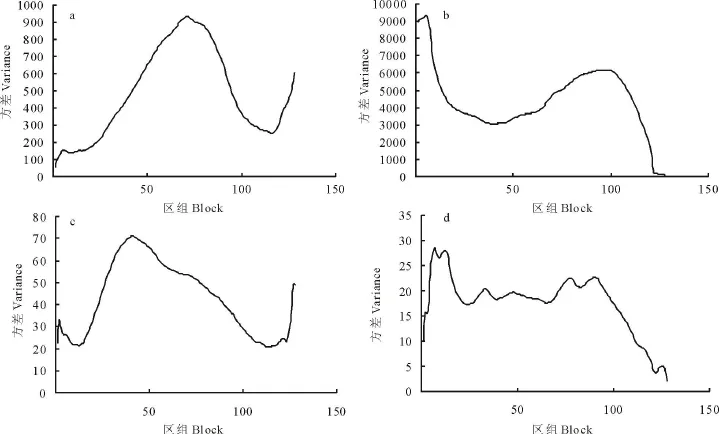
圖2 長治濕地扁稈藨草+蘆葦群系格局分析Fig.2 The pattern analysis of S.planiculmis+P.australis in Changzhi wetland
建群種小苜蓿的小斑塊規模為290 cm,中斑塊規模為750 cm,而大斑塊為990 cm(圖3);建群種雙穗雀稗的中斑塊規模為570 cm,大斑塊規模為890 cm,而小斑塊規模未能顯示出來,這可能與小苜蓿具有明顯的群集性生長特性有關;優勢種大車前的小斑塊規模為220 cm,中斑塊為650 cm,大斑塊規模為950 cm;優勢種飛蓬的小斑塊為240 cm,中斑塊為740 cm,而大斑塊未能顯示出來,與分布于水庫邊緣濕地的飛蓬相比,其群聚程度降低,說明種群的分布格局不僅與種的生物學特性密切相關,也與生境有關,這一結論驗證了王伯蓀[26]、張金屯[10]關于分布格局與植物種的生物學特性和環境因子作用有關,還與植物種在群落中的優勢地位有關的觀點。
2.2 群落的格局規模
圖4反映了2個土壤水分梯度的不同類型濕地群落格局規模斑塊的長度或寬度。扁稈藨草+蘆葦群系的格局規模為小斑塊6區組(60 cm),中斑塊75區組(750 cm),大斑塊150區組(1 500 cm)(圖4a);小苜蓿+雙穗雀稗群系的格局規模為小斑塊3區組(30 cm),中斑塊103區組(1 030 cm),大斑塊191區組(1 910 cm)(圖4b)。可見2個不同類型的濕地群落格局隨土壤水分減少而發生明顯變化,說明群落的分布格局既取決于優勢種或建群種自身的生物學特性,也與它們的環境條件密切相關[26]。

圖3 長治濕地小苜蓿+雙穗雀稗群系格局分析Fig.3 The pattern analysis of M.minima+P.paspaloides in Changzhi wetland
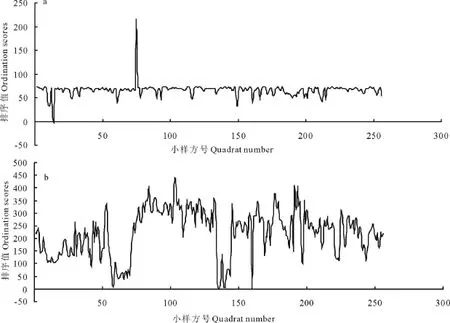
圖4 基于DCA第1軸的格局變化Fig.4 The community pattern based on the first DCA axes
扁稈藨草+蘆葦群系的格局規模為小斑塊5區組(50 cm),中斑塊93區組(930 cm),大斑塊212區組(2 120 cm)(圖5a);小苜蓿+雙穗雀稗群系的格局規模為小斑塊3區組(30 cm),中斑塊110區組(1 100 cm),大斑塊153區組(1 530 cm)(圖5b)。比較基于DCA第1、第2排序軸格局的結果,可以看出二者所反映的群落格局規模是基本一致的,因而在實際濕地植物群落格局分析中,可以任選一軸的排序值進行研究。

圖5 基于DCA第2軸的格局變化Fig.5 The community pattern based on the second DCA axes

圖6 基于DCA和TTLQV的長治濕地草本植物群落格局分析Fig.6 The pattern analysis of herbaceous community in Changzhi wetland by combining DCA and TTLQV
采用TT LQV進行格局分析,扁稈藨草+蘆葦群系的均方值表現出3個峰值,即3個格局規模,分別對應于區組7,41和91,相應的格局規模是:小斑塊(包括間隙)直徑為70 cm,中斑塊直徑為410 cm,大斑塊直徑為910 cm(圖6 a);小苜蓿+雙穗雀稗群系表現出1個格局規模,即區組52(520 cm)(圖6 b)。可見2個不同類型的濕地植物群落格局具有明顯差異,即隨土壤水分含量減少,濕地植物群落的格局規模減少,這種現象說明分布格局并不完全由植物本身特征決定,也與土壤水分等生境有關。扁稈藨草+蘆葦群系位于水庫邊緣,土壤水分含量較多,群落中濕生植物成分較多,扁稈藨草、蘆葦等群落優勢種在群落中占據主導地位,而小苜蓿+雙穗雀稗群系位于與水庫邊緣垂直的地帶,隨著與水庫邊緣距離的逐漸增加,土壤水分含量逐漸減少,群落中耐旱植物成分較多,并且放牧等人為干擾強度逐漸增大[27,28],造成濕地植物退化,群落的異質性較大,因此格局規模減少。
3 討論與結論
在群落格局分析中,應用DCA排序與TT LQV相結合的方法,計算簡單,結果清楚,可成功地揭示群落的格局特征和空間變化。張金屯[9,10]和李素清等[11]曾采用該方法,分析了亞高山草甸優勢種群和群落的格局,成功地揭示了亞高山草甸的格局特征。本研究表明,應用DCA排序與TT LQV相結合的方法,可合理、客觀地揭示長治濕地草本植物優勢種群和群落的格局特征,說明該方法同樣適用于濕地植物群落格局分析。并且基于DCA第1、第2排序軸反映的群落格局結果基本一致,因而,在實際濕地植物群落格局分析中,可以任選一軸的排序值進行研究。
長治濕地草本植物優勢種群的小斑塊規模一般為20~290 cm,大斑塊規模則多為900~1 270 cm,格局規模隨優勢種的生物學特性而變化。并且同一優勢種飛蓬在不同群落中的格局有所不同,隨著土壤水分含量的降低而群聚程度降低,說明種群的分布格局不僅受種的生物學特性影響,還受環境因子制約。該結論驗證了王伯蓀[26]和張金屯[10]的觀點。
長治濕地草本植物群落的分布格局與建群種扁稈藨草、蘆葦、小苜蓿、雙穗雀稗的格局密切相關,扁稈藨草+蘆葦群系的格局規模是小格局70 cm、中格局410 cm、大格局910 cm,小苜蓿+雙穗雀稗群系的格局規模為520 cm,二者的格局差異明顯。即隨著土壤濕度降低,濕地草本植物群落的格局規模增大,說明群落分布格局是主要優勢種格局的綜合反映,但建群種在群落格局的形成中起著決定性作用;隨著距水庫邊緣濕地距離的增加,放牧等人為干擾逐漸增加,群落的異質性增大,格局規模減小,說明群落分布格局并不完全由植物本身特征決定,也與生境條件有關,并且群落的分布格局與生境分布格局吻合。
[1]Greig-Smith P.The use of random and contiguous quadrats in the study of the structure of plant communities[J].Annual Botany,1952,(16):296-316.
[2]Clark P J,Evans F C.Distance to nearest neighbor as a measure of spatial relationships in population[J].Ecology,1954,35:445-453.
[3]Hill M O.The intensity of spatial pattern in plant communities[J].Journal of Ecology,1973,61:225-238.
[4]Greig-Smith P.Quantitative Plant Ecology(3rd ed.)[M].Oxford:Blackwell Scientific Publications,1983:54-104.
[5]張金屯.植被數量生態學方法[M].北京:中國科學技術出版社,1995:268-278.
[6]王本洋,余世孝.種群分布格局的多尺度分析[J].植物生態學報,2005,29(2):235-241.
[7]李先琨,黃玉清,蘇宗明.元寶山南方紅豆杉種群分布格局及動態[J].應用生態學報,2000,11(2):169-172.
[8]張金屯,米湘成,張峰,等.五臺山亞高山草甸小格局分析[J].應用與環境生物學報,1998,4(1):20-23.
[9]張金屯.群落二維格局分析的兩種方法[J].西北植物學報,2004,24(8):1448-1451.
[10]張金屯.蘆芽山亞高山草甸優勢種群和群落的二維格局分析[J].生態學報,2005,25(6):1264-1268.
[11]李素清,楊斌盛,張金屯.山西云頂山亞高山草甸優勢種群和群落的格局分析[J].應用與環境生物學,2007,13(1):9-13.
[12]徐連秀,蒙榮.短花針茅荒漠草原植物群落優勢種群分布格局研究——以內蒙古四子王旗荒漠草原為例[J].內蒙古農業大學學報(自然科學版),2008,29(2):64-67.
[13]張育新,馬克明,祁建,等.北京東靈山海拔梯度上遼東櫟種群結構和空間分布[J].生態學報,2009,29(6):2789-2796.
[14]魯為華,朱進忠,王東江,等.天山北坡兩種退化梯度下伊犁絹蒿種群空間分布的點格局分析[J].草業學報,2009,18(5):142-149.
[15]閆海冰,韓有志,楊秀清,等.華北山地典型天然次生林群落的樹種空間分布格局及其關聯性[J].生態學報,2010,(9):2311-2321.
[16]李立,陳建華,任海保,等.古田山常綠闊葉林優勢樹種甜櫧和木荷的空間格局分析[J].植物生態學報,2010,34(3):241-252.
[17]黃志偉,彭敏,陳桂琛,等.青海湖幾種主要濕地植物的種群分布格局及動態[J].應用與環境生物學報,2001,7(2):113-116.
[18]武玉珍,張峰.山西桑干河流域濕地植被優勢種群分布格局研究[J].植物研究,2006,26(6):737-741.
[19]徐洋,劉文治,劉貴華.生態位限制和物種庫限制對湖濱濕地植物群落分布格局的影響[J].植物生態學報,2009,33(3):546-554.
[20]方金.長治市濕地主要植被類型調查與研究[J].中國林副特產,2009,2:77-78.
[21]Hill M O,Gauch H G.Detrended correspondence analysis:an improved ordination technique[J].Vegetatio,1980,42:47-58.
[22]Braak C J F.Canoco-a Fortran Program for Canonical Community Ordination by[Partial][Detrended][Canonical]Correspondence Analysis,Principle Component Analysis and Redundancy Analysis[M].New York:Cornell University Press,1991.
[23]張金屯.數量生態學[M].北京:科學出版社,2004:257-259.
[24]趙靜敏.試論長治濕地的保護和利用[J].科技情報開發與經濟,2008,18(34):63-64.
[25]吳征鎰.中國植被[M].北京:科學出版社,1982:144-145.
[26]王伯蓀.植物群落學[M].北京:高等教育出版社,1987:41-44.
[27]趙登亮,劉鐘齡,楊桂霞,等.放牧對克氏針茅草原植物群落與種群格局的影響[J].草業學報,2010,19(3):6-13.
[28]桂東偉,雷加強,曾凡江,等.中昆侖山北坡策勒河流域生態因素對植物群落的影響[J].草業學報,2010,19(3):38-46.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
