駝背鱸外周血細胞的形態學研究
2011-03-14 06:06:50王靜香李加兒區又君
海洋科學 2011年4期
王靜香,李加兒,區又君
(1.中國水產科學研究院 南海水產研究所,廣東 廣州 510300;2.上海海洋大學 水產與生命學院,上海201306)
魚類的血液量較少,僅占體質量的 2%左右[1],但在生命活動中卻起著非常重要的作用。目前,國內外已有許多學者對魚類血液進行了研究,Valenzuela[2]、Ballarin[3]、Gao等[4]分別對虹鱒(Oncorhynchus mykiss)、波紋短須石首魚(Umbrina cirrosa)、泥鰍(Misgurnus anguillicaudatus)血液參數方面進行了研究;李長嶺[5]、陳剛[6]、蘇友祿等[7]分別對大彈涂魚(Boleophthalmus pectinirosris)、軍曹魚(Rachycentron canadum)的血細胞的發生進行了研究;還有一些學者對紅鰭笛鯛(Lutjanus erythopterus)[8]、黑鰭冰魚(Chaenocephalus aceratus)[9]、達氏鰉(Huso dauricus)[10]、歐洲鰻鱺(Anguilla anguilla)[11]、條石鯛(Oplegnanathus fasciatus)[12]、鯽
(Carassius auratus)[13]鲹、卵形鯧(Trachinotus ovatus)[14]等魚外周血細胞的形態進行了研究。有關駝背鱸的研究主要集中在形態和生物學性狀[15]、胚胎發育[16]、配合飼料研制[17]、線粒體細胞色素 b基因的序列分析[18]等方面,但在外周血細胞的形態結構方面尚未見過報道。
駝背鱸(Cromileptes altivelis),俗稱老鼠斑,隸屬科(Serranidae)、石斑魚亞科(Epinephelinae)、駝背鱸屬(Cromileptes)[15],是一種高級海產經濟食用魚類,價格昂貴。目前世界上已知的駝背鱸僅 1種,廣布于印度太平洋的暖水海域,東至菲律賓,西至非洲,南至澳大利亞海域,北至日本,中國見于南海。作者就駝背鱸外周血細胞的形態結構進行觀察,旨在為駝背鱸的生理學、繁殖生物學和病理學研究提供參考依據。
1 材料與方法
1.1 材料
試驗所用15尾駝背鱸于2009年9月采自廣州市黃沙水產市場,雌雄兼有,健康無病。體長為(26.48±1.08)cm,體質量為(465.75±97.96) g/尾。
1.2 方法
1.2.1 取材
用0.1 %肝素鈉潤洗過的無菌注射器,魚體尾部血管采血,一部分傾入肝素鈉潤洗過的試管中用作血細胞計數的原液,另一部分用于制作血涂片。
1.2.2 血細胞計數
將試管中的血液分為兩份,一份用 0.65 %NaCl溶液將血液稀釋 100倍,用于紅細胞的計數;另一份用2 mL冰醋酸,98 mL蒸餾水,10 g/L亞甲基藍溶液 3滴配制的溶液將血液稀釋 10倍用于白細胞的計數。用血球計數板在顯微鏡下進行血細胞計數。
1.2.3 光鏡樣品制備
常規方法制作血涂片,每尾魚血涂片 10張,將其中的5張玻片用Wright’s和Giemsa雙重染色法染色,剩余5張入95 %酒精固定10 min,然后浸入蘇木精染液染色5 min,流水沖洗5 min,再入伊紅染液染色 2 min,最后進行常規脫水,二甲苯透明,中性樹膠封片。每尾魚選取 5張血涂片(用 Wright’s和Giemsa雙重染色法),隨機選取100個視野,根據血細胞的形態差異和細胞的著色差異[6]對其上的2 916個白細胞在 Olympus顯微鏡油鏡下進行分類計數,同時進行拍照,測定血細胞(核)大小。
1.2.4 數據處理
試驗結果中數據用平均值±標準差的形式描述,并采用Excel和SPSS軟件對數據進行統計分析。
2 結果
2.1 血細胞的測定及分類計數
通過觀察和測定駝背鱸各類血細胞,結果見表1。從表1中可得出,各類白細胞所占比例依次為:淋巴細胞 37.26%,血栓細胞 30.82%,嗜中性粒細胞23.57%,單核細胞8.35%,嗜酸性粒細胞極少見到。此外,對血細胞的數量統計中紅細胞數 1.746 2×106個/mm3,白細胞數 4.781 7×103個/mm3。
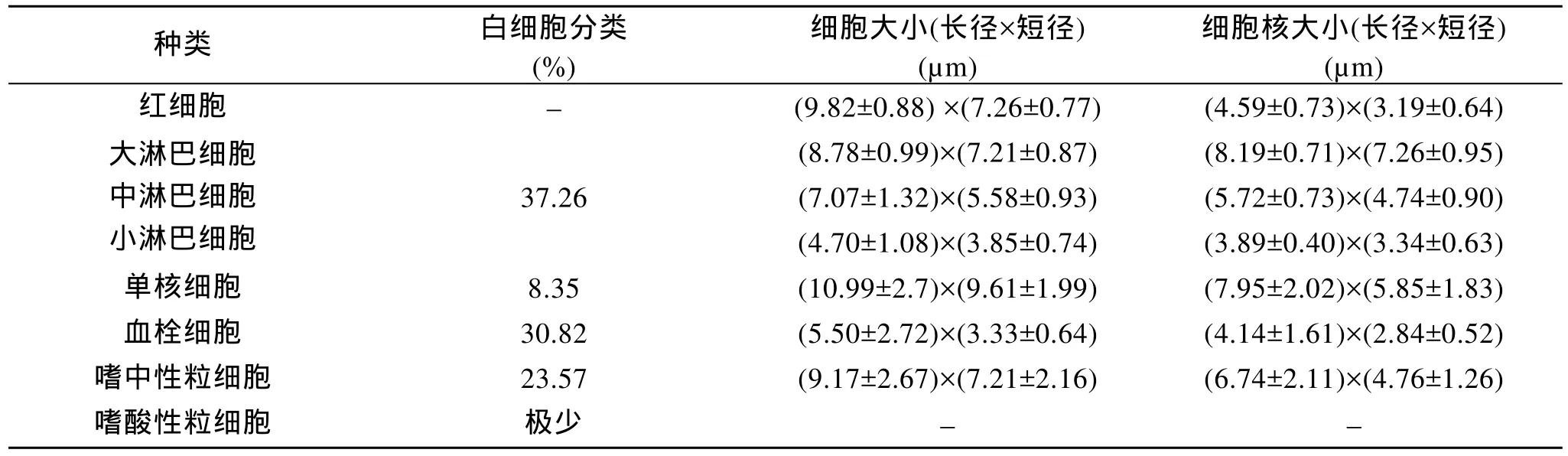
表1 駝背鱸血細胞的大小和白細胞的分類統計值Tab.1 Sizes of the blood cells and classified statistics of white blood cells of Cromileptes altivelis
2.2 紅細胞
正常成熟的紅細胞呈圓形或橢圓形(圖1-1),核呈圓形、橢圓形,外周血中也可看到處于不同發育階段的紅細胞,細胞核呈分葉型(像正在分裂的細胞核)(圖1-2)、也有不規則的形狀。位于細胞中央或偏側。細胞表面光滑,核質比接近1。細胞的胞膜清晰,胞質中血紅蛋白含量豐富且分布均勻。油鏡下觀察,細胞核的核膜清晰可見,核內染色質豐富呈網狀排列或零散。在駝背鱸的血涂片中可以看到幼紅細胞(圖1-3),細胞圓形或不規則形,胞核居中較正常紅細胞大,胞質呈堿性被 Wright’s和Giemsa雙重染色染成灰藍色,被改良的方法染成淺粉色,與淋巴細胞類似。此外,也可看到解體的紅細胞(圖1-4)、細胞膜破裂的紅細胞即過渡型(圖1-5,1-6)以及“核影”紅細胞(圖1-7)。解體的紅細胞,胞質變淺,區域擴大,形狀不規則,胞核也逐漸增大,顏色變淺,圓形或不規則形。而核影紅細胞,沒有細胞質呈現裸核狀態,與解體的紅細胞相比,其細胞核更大,隨著細胞核的擴大,染色質逐漸變淺最終消失。在駝背鱸血涂片中也可以看到正在分裂的紅細胞(圖1-8)。

圖1 駝背鱸的紅細胞Fig.1 Erythrocyte of Cromileptes altivelis
2.3 粒細胞
在駝背鱸的血涂片中,可以看到嗜酸性粒細胞(圖2-1)和嗜中性粒細胞(圖2-2,2-3,2-4),沒發現嗜堿性粒細胞。嗜中性粒細胞數量比較多,細胞呈圓形或不規則。細胞核腎形或分葉型,通常位于細胞的一側,胞質顏色清亮,細胞膜明顯,與淋巴細胞相比粒細胞胞質相對較多。嗜酸性粒細胞數量較少,細胞形狀不規則,細胞核呈圓形。胞質中有被染成橘紅色的顆粒。同時也能看到中性粒細胞的集群和白細胞聚集的現象(圖2-5,2-6)。

圖2 駝背鱸的粒細胞Fig.2 Granulocyte of Cromileptes altivelis
2.4 單核細胞
單核細胞是白細胞中體積較大的一類,與嗜中性粒細胞很難區分。細胞呈圓形、橢圓形或形狀不規則(圖3-1),細胞邊緣粗糙。細胞質豐富呈堿性,被染成深藍色。胞質中含有少量的空泡(圖3-2),且胞質有偽足外伸的現象。細胞核圓形、卵圓形、腎形、菱形、也有不規則的形狀通常偏于一側,也有的居中。細胞核占整個細胞的1/3~1/2。偶爾也可見到細胞核呈啞鈴型或長條形偏于一側,與細胞膜相切,中央有一個很大的空泡,空泡外有一層薄薄的細胞質(圖3-3)。

圖3 駝背鱸的單核細胞Fig.3 Monocyte of Cromileptes altivelis
2.5 淋巴細胞
在駝背鱸的血涂片中可看到 3種淋巴細胞(圖4-1,4-2,4-3):大淋巴細胞,中淋巴細胞,小淋巴細胞。小淋巴細胞細胞核比紅細胞的細胞核小或接近。中淋巴細胞的細胞核略大于紅細胞細胞核,大淋巴細胞核比紅細胞細胞核大得多。淋巴細胞都有類似圓形的細胞核,胞質少,細胞表面不光滑,通常有偽足樣的突起。有時淋巴細胞的細胞質呈鐮刀狀,緊貼在胞核的一側(圖4-4)。
2.6 血栓細胞
血栓細胞的細胞核呈長桿狀、橢圓形、圓形、紡錘形,胞質少,核質比大,細胞表面光滑,體積小,外形表現多樣,有蝌蚪形(圖4-5)、紡錘形(圖4-6)、水滴形(圖4-7)、短棒形(圖4-8),一般單獨存在,有時可見到正在分裂的血栓細胞(圖4-9)。
3 討論
3.1 幾種魚類血細胞數量的比較
魚類的血液中主要存在紅細胞、血栓細胞、淋巴細胞、粒細胞、單核細胞5種血細胞,駝背鱸的血細胞與大多數魚類相似,形態差異不明顯,僅白細胞中各細胞所占的比例有所差異。有的學者[19]研究表明血栓細胞數目最多,嗜酸性細胞的數量最少(嗜堿性細胞一般很難看到),也有學者[11]發現淋巴細胞的數量最多,嗜酸性細胞最少。駝背鱸的外周血細胞中淋巴細胞數量最多,嗜酸性細胞最少。各白細胞出現數量的差異,作者推測可能有兩方面的原因造成的,一是魚類處于不同的生理狀態;二是在血涂片制作過程中涂片不均勻或計數過程中重復和錯誤判斷。
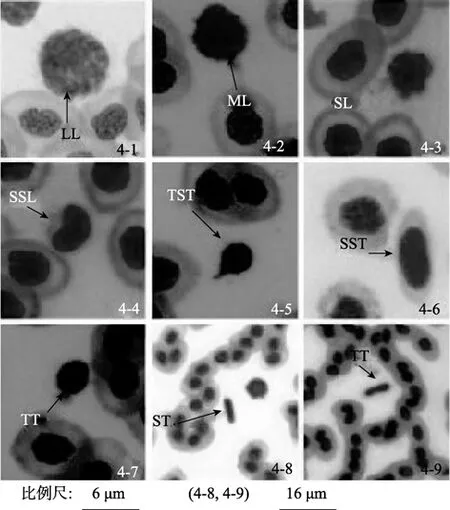
圖4 駝背鱸的淋巴細胞和血栓細胞Fig.4 Lymphocyte and thrombocyte of Cromileptes altivelis
在血細胞的數量上,作者將駝背鱸血細胞的數量與軍曹魚[6]、達氏鰉[10]、卵形鯧鲹[14]、斑點叉尾(Ictalurus punctatus)[20]、鰱(Hypophthalmichys molitrix)[21]等魚血細胞數量進行比較。從表2可見海水魚類如駝背鱸、卵形鯧鲹、軍曹魚以及淡水魚如斑點叉尾等魚類的紅細胞數量均多于達氏鰉,駝背鱸的紅細胞數量略少于鰱;達氏鰉和鰱的白細胞數量均大于駝背鱸、軍曹魚、卵形鯧鲹、斑點叉尾。經方差分析和多重比較,結果表明:在紅細胞數量上,駝背鱸、達氏鰉、鰱、斑點叉尾、卵形鯧鲹、軍曹魚兩兩之間差異均顯著(P<0.05);在白細胞數量方面,駝背鱸與斑點叉尾、斑點叉尾與卵形鯧鲹、卵形鯧鲹與軍曹魚、達氏鰉與鰱之間差異不顯著(P>0.05),其余差異均顯著(P<0.05)。從以上結論可以看出不同魚類的血細胞在數量上有一定的差異,王義強[22]等認為魚類的血細胞數量會因魚類生存環境、生理狀況、種類、年齡、疾病、季節變化、取血部位不同(如從心臟或尾動脈處取血)等各種因素變化而有所不同。
3.2 駝背鱸紅細胞的形態特點及其功能
駝背鱸的血涂片中觀察到了成熟的紅細胞、幼稚紅細胞、解體的紅細胞、正在分裂的紅細胞以及核影。在歐洲鰻鱺[11]、條石鯛[12]、卵形鯧鲹[14]、花尾胡椒鯛(Plectorhinchus cinctus)[23]、史氏鱘(Acipenser schrencki)[24]、海鰻(Muraenesox cinereiu)[19]等硬骨魚類的外周血中也觀察到了直接分裂的紅細胞。這些實驗結果表明魚類的紅細胞除經造血器官生成外,還可通過直接分裂而產生。駝背鱸的血涂片中存在很多分葉核,類似有絲分裂的細胞核,Ellis[25]把紅細胞核產生分葉的現象認為是病態的斷裂增殖或一種紅細胞衰老方式。
紅細胞作為魚類血液中數量最多的一種血細胞,同時也是魚類體內通過血液運送氧氣的最主要的媒介。在駝背鱸的紅細胞中有一些幼稚紅細胞,呈圓形略大于成熟紅細胞,胞質染成淺藍色。根據紅細胞的發生,由原紅細胞經幼稚紅細胞到成熟紅細胞,胞體及胞核均由大變小,形態由圓形變為橢圓形,核固縮濃染[26],這一點在駝背鱸外周血中得到證實。此外,還觀察到“核影”和解體的紅細胞,“核影”是紅細胞膜破裂,胞質外溢擴散消失,細胞核裸露,染色質疏松,逐漸解體而形成;而解體紅細胞只是正常紅細胞到“核影”形成的一個過渡類型。造成駝背鱸血涂片中這一現象的原因,作者認為首先是細胞衰老達到正常壽命而自動解體,其次可能是在制做血涂片過程中破壞了紅細胞。“核影”在紅鰭笛鯛[8]、海鰻[19]的血涂片中也廣泛存在。

表2 不同魚類血細胞數量的比較Tab.2 Comparison of the blood cell counts among different fish species
3.3 駝背鱸白細胞的形態特點及其功能
3.3.1 有粒白細胞
有粒白細胞有3種類型,嗜中性白血細胞、嗜酸性白血細胞和嗜堿性白血細胞。在駝背鱸的外周血涂片中可觀察到嗜中性白血細胞、嗜酸性白血細胞兩種細胞。嗜中性白血細胞具有吞噬作用,能做變形運動[27]。駝背鱸血涂片中嗜中性白血細胞數目最多,有的形狀不規則,細胞比較大,內含空泡,證實了嗜中性白血細胞吞噬機能。嗜堿性白血細胞和嗜酸性白血細胞一般在魚類的血涂片中很少見到,Watson等[28]提出這兩種細胞的顆粒易溶于水,在制片中被溶解掉而不易被觀察。林光華等[29]在 100張革胡子鲇(Claris lareza)的血涂片上僅發現一個嗜堿性粒細胞。金魚(Carassius auratusvar.)[28]外周血涂片中只見少量嗜堿性白血細胞,本實驗中沒發現。花(Hemibarbus maculatus)[30]外周血中嗜酸性粒細胞較多,本實驗中很少發現。
3.3.2 淋巴細胞和單核細胞
魚類淋巴細胞和哺乳動物的淋巴細胞的形態很相似,按照大小可分為大小兩種。但 Ellis等[25]認為不應將淋巴細胞分為大小兩類,因為它們只是不同功能狀態的同一種細胞不同表現形式;Barber[9]等發現許多淋巴細胞的體積是連續變化的,不應該用大小來界定。但也有些學者將其分為大小兩類,認為它們是具有不同功能的兩類細胞[31]。本次實驗中,觀察到的淋巴細胞不僅有大與小的區別,其形狀也有較大差異。作者將介于大小淋巴細胞之間的淋巴細胞稱為中淋巴細胞。駝背鱸的外周血淋巴細胞形態明顯不同,是否在功能上也有明顯不同,有待對其內部結構和生物學意義進一步研究。
單核細胞存在于所有脊椎動物中,擔負著非特異性免疫的重要作用[32]。魚類單核細胞很少,在一些魚的血液中未見單核細胞,如:鯽[13]。駝背鱸血液中存在單核細胞,有的圓形,有的不規則,胞質中含有空泡,說明單核細胞具有吞噬機能。許多學者對紅鰭笛鯛[8]、歐洲鰻鱺[11]、條石鯛[12]的研究中也證實這一點。
3.3.3 血栓細胞
魚類的“血小板”即血栓細胞,是白細胞中體積較小的一種細胞,與小淋巴細胞較難區分,具有許多種形態,在駝背鱸的血涂片中觀察到4種形態:蝌蚪形、短棒形、水滴形和紡錘形。軍曹魚[6]中可見多種形態的血栓細胞,歐洲鰻鱺[11]有5種形態,鰈[33]、日本白鯽(Carassius auratus cuvieri)[34]有4種形態的血栓細胞,表明血栓細胞的形態因魚的種類而有差別。許多研究報道表明不同形態的血栓細胞是其凝血功能狀態的反映[35]。Barber等[9]推測不同形態顯示了不同的發育時期或不同的功能狀態,也表明種間存在著差異。同時,在觀察血栓細胞的過程中發現,血涂片中紅血細胞凝血比較多的片子中,血栓細胞比較多。本文將駝背鱸血栓細胞歸為白細胞中的一種,與條石鯛[12]、卵形鯧鲹[14]的血栓細胞的分類一致;也有的將血栓細胞歸為白細胞以外的一種血細胞,如鰱、鳙(Aritichthys nobilis)[36]。
[1]樓允東.組織胚胎學[M].北京:中國農業出版社,2000:111-113.
[2]Valenzuela A E,Silva V M,Klempau A E.Some changes in the haematological parameters of rainbow trout (Oncorhynchus mykiss) exposed to three artificial photoperiod regimes [J].Fish Physio Biochem,2007,33(1):35-48.
[3]Ballarin L,Dall’Oro M,Bertotto D,et al.Haematological parameters inUmbrina cirrosa(Teleostei,Sciaenidae):a comparison between diploid and triploid specimens[J].Comp Biochem Physiol,2004,138A:45-51.
[4]Gao Z,Wang W,Abbas K,et al.Haematological characteri-zation of loachMisgurnus anguillicaudatus:comparison among diploid,triploid and tetraploid specimens [J].Comp Biochem Physiol,2007,147(4):1001-1008.
[5]李長嶺,曹伏君,劉楚吾,等.大彈涂魚血細胞的發生[J].中國水產科學,2002,8(1):10-14.
[6]陳剛,周暉,張東健,等.軍曹魚血液指標及血細胞發生的觀察[J].水生生物學報,2005,29(5):564-570.
[7]蘇友祿,徐力文,馮娟,等.軍曹魚稚魚外周血細胞及其形態學觀察[J].南方水產,2007,3(1):48-52.
[8]曹伏君,朱曉燕.紅鰭笛鯛外周血細胞的顯微結構觀察[J].海洋科學,2006,30(5):32-35.
[9]Barber D L,Mills Westermann J E,White M G.The blood cells of the Antarctic icefishChaenocephalus aceratusL?nnberg:light and electron microscopic observations [J].J Fish Biol,1981,19(1):11-28.
[10]周玉,潘風光,李巖松,等.達氏鰉外周血細胞的形態學研究[J].中國水產科學,2006,13(3):480-484.
[11]周玉,郭文場,楊振國,等.歐洲鰻鱺外周血細胞的顯微和超微結構[J].動物學報,2002,48(3):393-401.
[12]胡玲玲,李加兒,區又君,等.條石鯛外周血細胞的顯微結構[J].福建農林大學學報(自然科學版),2009,38(4):384-387.
[13]朱洪文,王浩,秦國強.鯽魚外周血細胞顯微和亞顯微結構的觀察[J].動物學研究,1985,6(2):147-153.
[14]許曉娟,李加兒,區又君,等.深水網箱養殖卵形鯧鲹血液指標[J].動物學雜志,2008,43(6):109-116.
[15]區又君,李加兒,陳福華.駝背鱸的形態和生物學性狀[J].中國水產科學,1999,6(1):24-26.
[16]區又君.駝背鱸的胚胎發育[J].海洋科學,2006,30(8):17-19.
[17]Rossita S,Wing Keong N,Saleem M.Replacement of fish meal with poultry by-product meal in diets formulated for the humpback grouper,Cromileptes altivelis[J].Aquaculture,2007,273(1):118-126.
[18]區又君,李加兒,吳勇,等.駝背鱸線粒體細胞色素b基因的序列分析[J].南方水產,2008,4(5):51-54.
[19]謝嘉華,陳朝陽.海鰻外周血細胞的顯微結構[J].動物學雜志,2003,38(6):14-18.
[20]向建國,周進,金宏.斑點叉尾的生物學與生理生化特性研究[J].湖南農業大學學報(自然科學版),2004,30(4):355-358.
[21]米瑞芙.草魚、鯉和鰱血液學指標的測定[J].淡水漁業,1982,9(4):10-16.
[22]王義強,黃世蕉,趙維信,等.魚類生理學[M].上海:上海科學技術出版社,1990:130-138.
[23]Fujun C,Chuwu L,Changling L.Microstructure of peripheral blood inPlectorhinchus cinctus[J].Acta Laser Biology Sinica,2001,10(3):181-186.
[24]Hongbai L,Yuping H,Qiuzhi Q,et al.Microstructuer and ultrastructure of peripheral blood cells of amur sturgeonAcipenser schrenckibrandt[J].Acta Hydrobiologica Sinica,2006,30(2):214-220.
[25]Ellis A E.Bizarre forms of erythrocytes in a specimen of plaice,Pleuronectes platessaL.[J].J Fish Dis,1984,7(5):411-414.
[26]郭瓊林,盧全章.草魚腎臟和脾臟血細胞發育過程的觀察[J].水生生物學報,1993,17(1):40-45.
[27]林浩然.魚類生理學[M].廣州:廣東高等教育出版社,1999:82-85.
[28]Watson L J,Shechmeister I L,Jackson L L.The haematology of goldfish,Carassius auratus[J].Cytologia,1962,28:118-130.
[29]林光華,張豐旺,洪一江.革胡子鯰血液常數值的周年變化[J].動物學報,1991,37(3):341–342.
[30]趙海鵬,王志堅,楊求華,等.花外周血細胞顯微結構[J].安徽農業科學,2009,37(22):10542-10544,10627.
[31]林光華.成年草魚外周血細胞的超微結構[J].動物學報,1996,42(2):123-128.
[32]Morrow W J W,Pulsford A.Indentification of peripheral blood leucocytes of the dogfish (Scyliorhinus caniculaL.) by electron microscopy[J].J Fish Biol,1980,17(4):461-475.
[33]E1lis A E.The leucocytes of fish:a review [J].J Fish Biol,1977,11(5):453-491.
[34]謝艷霞,林光華.日本白鯽外周血細胞顯微及亞顯微結構的研究[J].動物學雜志,1996,31(11):12-16.
[35]Burrows A S,Fletcher T C,Manning M J.Haematology of the turbot,Psetta maxima(L.):ultrastructural,cytochemical and morphological properties of peripheral blood leucocytes[J].J Appl Ichthyol,2001,17(2):77-84.
[36]林光華,張豐旺,洪一江,等.二齡鰱和鳙血液的比較研究[J].水生生物學報,1988,22(1):9-16.
