谷氨酸和谷氨酰胺轉運系統的研究進展
2011-03-14 05:33:18王秋菊范明哲
動物營養學報 2011年6期
關鍵詞:系統
王秋菊 許 麗* 范明哲
(1.東北農業大學動物科技學院動物營養與飼料系,哈爾濱 150030;2.圭爾夫大學安大略農業學院動物與家禽科學系,圭爾夫 N 1G 2W 1,加拿大)
谷氨酸(Glu)作為動物黏膜主要的能源物質之一,可以氧化供能,為動物機體提供能量,是腸內能量生成的最大貢獻者[1],是與腸黏膜生長和代謝相關的重要氨基酸之一[2],對幼畜的生長發育尤為重要。谷氨酸可作為氨基酸合成的前體物、其他氨基酸分解代謝或蛋白質從頭合成所需的 α-氨基和氨氮的碳受體,并可維持機體的氮平衡[3]。谷氨酸與半胱氨酸及甘氨酸均為合成谷胱甘肽的底物[4],為動物腸道提供抗氧化劑。另外,谷氨酸還可作為興奮性氨基酸神經遞質在哺乳動物的神經系統中起作用[5]。谷氨酰胺(Gln)是哺乳動物血漿和母豬乳汁中一種含量非常豐富的游離氨基酸,也是動物腸黏膜主要的能源物質。谷氨酰胺對腸黏膜具有保護作用,能增加腸道的血流量和氧耗量,對缺血缺氧造成的腸黏膜物理屏障損傷具有一定的修復功能;在完全胃腸外營養輸液期間,谷氨酰胺可以被磷酸激活的谷氨酰胺酶水解為谷氨酸和氨,是谷氨酸的主要來源,為還原型谷胱甘肽合成提供前體,對機體抗氧化劑的合成起重要作用。谷氨酸和谷氨酰胺的轉運對維持機體內谷氨酸的含量尤其重要,因此,本文就谷氨酸和谷氨酰胺轉運系統的分類及作用機制做一綜述。
1 谷氨酸和谷氨酰胺轉運系統分類
氨基酸轉運系統廣泛存在于機體中,按照轉運氨基酸的性質可以分為中性、堿性和酸性氨基酸轉運載體;按照轉運是否依賴 Na+可以分為Na+依賴氨基酸轉運載體和非 Na+依賴氨基酸轉運載體;按照氨基酸轉運載體的底物特異性、親和力和轉運特點可以分為 XAG-、XC-、y+、A、ASC、B0、B0,+、L、N、b和 y+L轉運系統。 XAG-轉運系統包括 5種高親和力的氨基酸轉運載體,分別為谷氨酸 -天冬氨酸轉運載體(GLAST)或稱為興奮性氨基酸轉運載體 1(EAAT1)、谷氨酸轉運載體 -1(GLT-1)或稱為興奮性氨基酸轉運載體 2(EAAT2)、興奮性氨基酸載體 1(EAAC1)或稱為興奮性氨基酸轉運載體 3(EAAT3)以及興奮性氨基酸轉運載體 4(EAAT4)和興奮性氨基酸轉運載體 5(EAAT5)[6];ASC轉運系統包括 2種中性氨基酸轉運載體,分別為中性氨基酸轉運載體 1(ASCT1)和中性氨基酸轉運載體 2(ASCT2)或稱為谷氨酰胺轉運載體。XAG-和 ASC轉運系統均屬于溶質載體 1(SLC1)家族,GLAST、GLT-1、EAAC1、EAAT4和 EAAT5也可分別表示為SLC1A 1、SLC1A2、SLC1A 3、SLC1A6和 SLC1A7;ASCT1和 ASCT2也可分別表示為 SLC1A 4和SLC1A 5。盡管這些轉運載體之間具有相似的可預測結構(同源性),但轉運機制的不同使它們表現出不同的功能[7]。
高親和力的谷氨酸轉運載體可以介導谷氨酸和胱氨酸的轉運,同時與 3個 Na+和 1個 H+的正向轉運及 1個K+的逆向轉運相偶聯,而 ASC轉運載體可以介導依賴 Na+變化的小分子中性氨基酸的轉運,如谷氨酰胺和天冬酰胺等。谷氨酸轉運載體的偶聯方式允許逆谷氨酸濃度梯度轉運谷氨酸到細胞內[6]。
1.1 谷氨酸轉運系統分類
谷氨酸轉運系統有 XAG-和XC-系統,如圖 1所示。谷氨酸轉運的首要途徑是通過高親和力的XAG-轉運系統[8],依賴 Na+從細胞外轉運谷氨酸入細胞內來完成的;XC-轉運系統為谷氨酸 -胱氨酸交換介導系統,通過將谷氨酸轉運出細胞外置換胱氨酸進入細胞[9],為非 Na+依賴系統。
谷氨酸的攝取除了依靠其轉運載體直接從細胞外轉運外,還可以以谷氨酰胺為前體物,通過谷氨酰胺酶催化谷氨酰胺合成谷氨酸[4]。
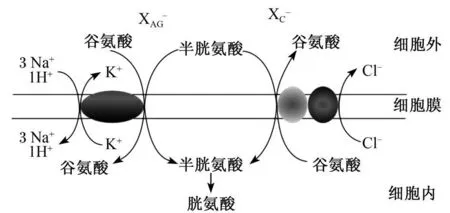
圖 1 谷氨酸的轉運Fig.1 Transportation of g lutamate[10]
1.2 谷氨酰胺轉運系統分類
谷氨酰胺轉運系統包括 A、N、y+L、ASC、B0、L和 b轉運系統。前 5種轉運系統為 Na+依賴轉運系統,轉運過程需要 Na+/K+和 ATP提供能量,進行谷氨酰胺的轉運;后 2種轉運系統轉運谷氨酰胺過程不需要 Na+參與[11]。Na+依賴轉運是谷氨酰胺轉運的主要轉運方式[12],其中 ASC和 B0轉運系統是近些年的研究熱點,而 ASC轉運系統中的 ASCT2是高親和力的轉運谷氨酰胺的載體,表達最強,對谷氨酰胺轉運起重要作用,還可以少量轉運谷氨酸。
表 1歸納了谷氨酸和谷氨酰胺轉運系統載體的種類、組織分布、底物特異性及離子依賴等特點。
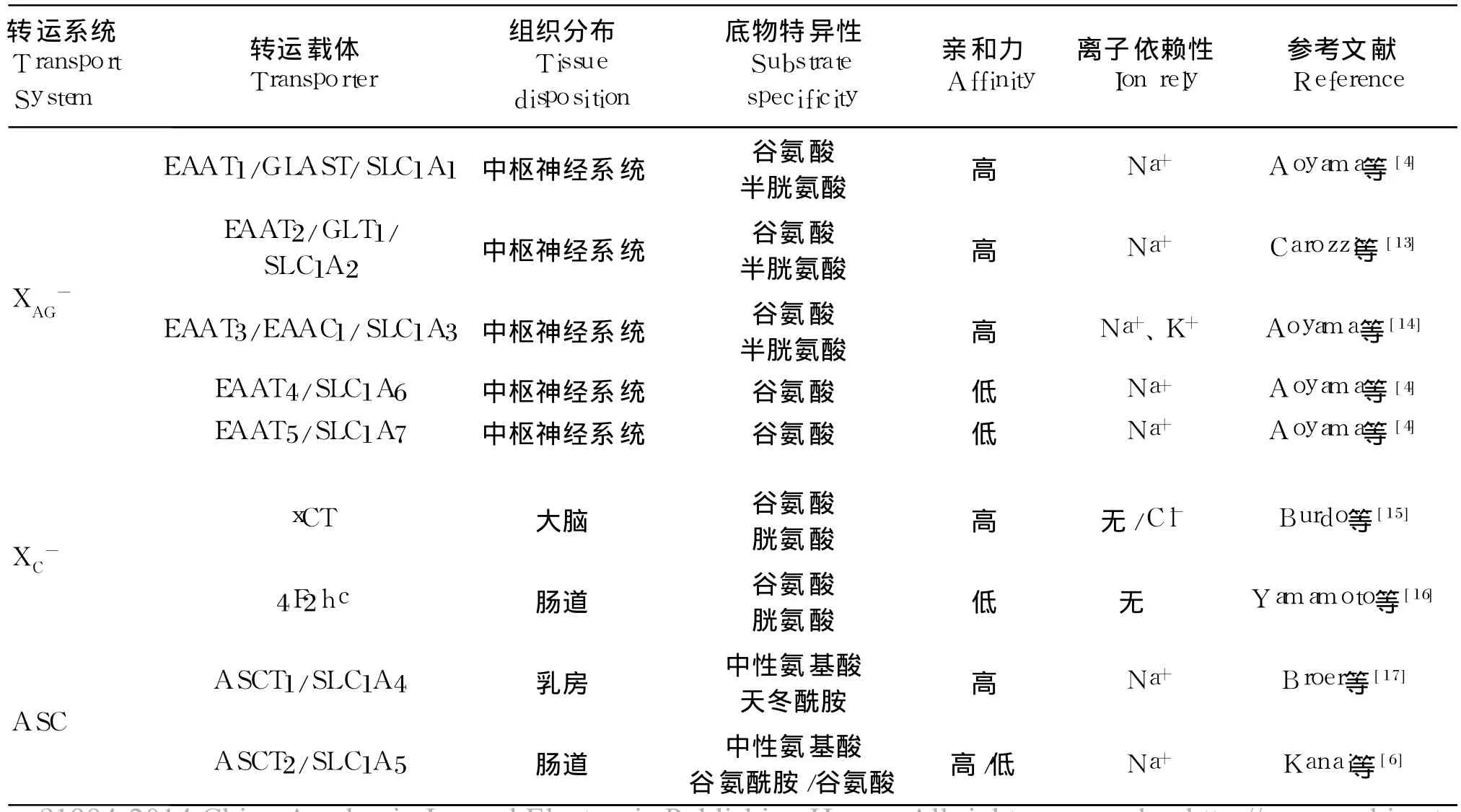
表 1 谷氨酸和谷氨酰胺轉運載體的種類、組織分布、底物特異性、親和力和離子依賴Table 1 Category,tissue disposition,substrate specificity,affinity and ion rely of glutamate and glutam ine transporters
2 谷氨酸和谷氨酰胺轉運系統及主要載體特點
2.1 XAG-轉運系統
2.1.1 XAG-轉運系統分類及特點Na+依賴的興奮性氨基酸轉運載體或谷氨酸轉運載體可以調節中樞神經系統細胞內外谷氨酸的轉運,并調節細胞外谷氨酸的濃度,屬于 X-AG轉運系統。XAG-轉運系統可以發揮轉運載體和離子通道的作用[18]。Na+依賴谷氨酸轉運是一個生物電傳導的過程[19],包括 2個不同的半循環過程:1個谷氨酸和 3個 Na+、1個 H+結合,結合位點轉向將谷氨酸運送到細胞內;同時伴隨 K+釋放到細胞外[20],這個過程可表示為 3Na+∶1H+∶1K+∶1谷氨酸。
目前已證實有 5種興奮性氨基酸轉運載體,分 別 是 GLAST、GLT-1、EAAC1、EAAT4和EAAT5[14]。星形膠質細胞主要表達 GLAST和GLT-1,神經系統主要表達 EAAC1、EAAT4和EAAT5,EAAT4只能在小腦浦肯野細胞表達,視網膜表達 EAAT5,而 EAAC1可以通過神經系統廣泛表達于神經元及非神經組織中[21],EAAC1轉運谷氨酸的速度約是轉運載體 GLAST和 GLT-1轉運速度的 10倍,是 XAG-系統中最重要的谷氨酸轉運載體。
2.1.2 EAAC1的功能特性
EAAC1首先是一種高親和力的谷氨酸轉運載體,在保持細胞外谷氨酸濃度,防止興奮過度和興奮性中毒方面起關鍵作用[22]。EAAC1與其他興奮性氨基酸載體不同,它不是神經特異性,可以在一些非神經組織表達,比如前胃、肝臟、腎、胰腺[23]。EAAC1除了具有清除細胞外谷氨酸的作用,還具有轉運半胱氨酸的作用[24]。EAAC1對半胱氨酸和谷氨酸轉運的一個重要區別在于它能更有效地遠距離轉運半胱氨酸[14]。
Kiyama等[25]觀察到在小鼠的運動神經元損傷后 EAAC1表達下降,而這種現象未在大鼠中發現。EAAC1表達受到抑制和運動神經元死亡有關,這種現象引發關于 EAAC1潛在的神經保護機制的研究。EAAC1這種獨特的抗凋亡機制可以在搶救運動神經元損傷過程中發揮作用,這表明EAAC1在預防神經元損傷方面具有多重調節機制。
2.1.3 EAAC1的調節機制
EAAC1的表達活動受細胞內蛋白激酶Cα亞型(PKCα)和磷脂酰肌醇 -3-激酶(PI3K)參與的信號通路的高度調節,這些調節過程在翻譯水平或轉錄后水平都可以發生[26]。
血清和糖皮質激素調節蛋白激酶 1(SGK 1)和3-磷酸肌醇依賴性蛋白激酶 1(PDK 1)可以正調控 EAAC1的表達[27],而 δ-阿片受體 (DOR)、谷氨酸轉運聯合蛋白 3-18(GTRAP3-18)和磷脂酰肌醇三羥基激酶抑制劑渥曼青霉素可以負調控EAAC 1的表達[19]。其中 GTRAP3-18是一種含有188個氨基酸,相對分子質量為 22 500的蛋白質[28]。GTRAP3-18可以直接和 EAAC1主鏈的C-末端相連從而負調控 EAAC1對谷氨酸的轉運[19,29]。GTRAP3-18位于內質網內,可以通過限制 EAAC1從內質網流出來阻止 EAAC 1成熟[30]。
EAAC1表達的另外一種調節機制是使羧基端磷酸化。465位的絲氨酸殘基被認為是重要的磷酸化位點,可以控制 EAAC1在細胞內的空間分布[31]。
血小板源性生長因子(PDGF)可以通過激活蛋白激酶 B(Akt)和 PI3K所組成的信號通路來途徑提高 EAAC1的表達[32],如圖 2所示。激活蛋白激酶 C(PKC),尤其是 PKCα,可以提高細胞膜表面 EAAC1的表達和啟動對谷氨酸的轉運活動。然而,PKCα可以不通過改變細胞膜的構象增強EAAC1的表達[33]。12-十四酸佛波酯 -13-乙酸鹽(PMA)是 PKC的激活因子,可以提高細胞膜表面 EAAC1的表達和活性[34]。
2.2 XC-轉運系統
2.2.1 XC
-轉運系統分類及特點XC
-系統是谷氨酸/胱氨酸轉運載體,1980年,在培養的人體二倍體成纖維細胞中首次被發現[35]。XC
-系統由 2種亞基載體成分組成:表面抗原重鏈 4F2hc(也稱為 CD98)和特異性的輕鏈載體 xCT。在 XC-系統作用下,細胞釋放 1分子的谷氨酸,并攝取 1分子胱氨酸入胞,兩者形成偶聯轉運。胱氨酸的轉運包括 2部分,一部分在胞內迅速被還原成半胱氨酸,參與胞內重要自由基清除劑谷胱甘肽的合成;另一部分則被轉運出細胞外,氧化成胱氨酸,重新參與 XC-系統循環,因而形成了 1個胱氨酸、半胱氨酸氧化還原循環回路。XC-系統轉運功能依賴于谷氨酸及胱氨酸跨膜濃度差高低,是非 Na+依賴轉運系統。
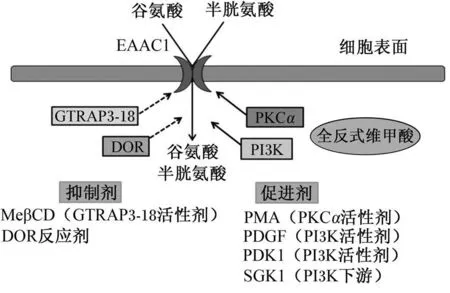
圖 2 EAAC 1的調節機制Fig.2 Regulatorym echanisms of EAAC 1[4]
從結構上來說,XC-轉運系統屬于異側的氨基酸轉運載體(HAT)家族中的一員。這些轉運載體是由一條重鏈(HSHAT)和一條輕鏈(LSHAT)通過二硫鍵偶聯在一起形成的。重鏈參與異二聚體向細胞膜的運輸,而輕鏈則負責轉運和底物特異性。對于XC-轉運系統的重鏈 4F2hc僅有 1個跨膜區,被推測可能不具備轉運活性。因此,xCT才是XC-轉運系統中起關鍵作用的亞單位,負責特異性轉運胱氨酸和谷氨酸。
2.2.2 xCT的轉運功能
xCT轉運載體有 2個主要功能:一方面它介導細胞內外谷氨酸/胱氨酸轉運,通過交換攝取胱氨酸進入細胞內,以滿足細胞內谷胱甘肽合成的需要。這對保持細胞內谷胱甘肽的水平尤為重要,因為谷胱甘肽是保護細胞免受氧化應激和其他化學物質損傷所必需的。另一方面,它可以維持細胞胱氨酸和半胱氨酸的氧化還原平衡。在細胞外的環境中,半胱氨酸迅速被氧化成胱氨酸,因此胱氨酸在循環中尤其在培養基中是主要的氨基酸形式,而在細胞內則以半胱氨酸為主[16]。
2.2.3 xCT的轉運機制
在體內,向細胞內轉運的胱氨酸與向細胞外轉運的谷氨酸按照濃度梯度呈 1∶1的交換(圖 3)。細胞內的胱氨酸很快削減至半胱氨酸,而細胞外的氧化條件則有助于胱氨酸的形成。有報道表明,胱氨酸與谷氨酸交換運輸時依賴 Cl-的存在,不過其可能是在低濃度的胱氨酸(<10μmol),而不是高濃度(>1 mmol)時發生;細胞內如果谷氨酸不足,可以導致 XC-系統活性降低,減少胱氨酸轉運[10]。

圖3 XC-介導的谷氨酸/胱氨酸轉運Fig.3 G lutamate/cystine system transported by X-[10]C
在成纖維細胞中,谷胱甘肽的清除會誘導XC-過程,導致非 Na+依賴谷氨酸轉運的增加,因此增加了細胞內谷胱甘肽的合成。另外,有研究發現氧化應激在某些細胞培養中對 XC-有正向調控功能,即 XC-介導的谷氨酸轉運可由氧化應激引起[8]。XC-過程的正調控會使細胞外聚集更多的谷氨酸,這樣潛在引起了胞外細胞毒性增加的可能[10],同時抑制胞內胱氨酸的含量,導致胞內谷胱甘肽水平的下降。
2.3 ASC轉運系統
2.3.1 ASC轉運系統分類及特點
Na+依賴性氨基酸轉運系統 ASC是經典的中性氨基酸轉運系統[17,36],對谷氨酰胺和天冬酰胺等小分子氨基酸具有很高的親和力。ASC轉運載體系統包括 ASCT1和 ASCT2,已經在人類和小鼠中鑒定出來[36]。與谷氨酸轉運載體需要 K+來使其構象改變相比,ASC轉運載體不需要和 K+偶聯[17]。同樣,ASC轉運載體與 H+的轉運也是非偶聯的[37]。
2.3.2 ASCT2的功能特性
ASCT2可以以很高的親和力轉運谷氨酰胺,為谷氨酸提供前體物,并維持細胞內谷氨酸的動態平衡[6,37],而 ASCT1沒有此功能。ASCT2可以在肺、骨骼肌、大腸、腎、睪丸以及脂肪組織等表達,在腎和腸道,ASCT2分別位于近端小管細胞和腸上皮細胞刷狀緣表面[38]。盡管谷氨酸轉運載體和 ASC在轉運的底物上有明顯的區別,但底物識別方面的共同特性反映出它們結構的相似性。比如谷氨酸轉運載體,尤其是 EAAC1,可以轉運中性氨基酸半胱氨酸。反之亦然。盡管 ASCT2與谷氨酸的親和力很低,但 ASCT2仍然可以轉運谷氨酸[36-37],低 pH可以增強 ASCT2對谷氨酸的轉運。
2.3.3 ASCT2的轉運機制
疏水性分析顯示,ASCT2可能存在 10個跨膜區域[37],在跨膜區 3、4之間的細胞外區域可能存在 2個 N-糖基化位點,細胞內區域可能存在 2個蛋白激酶 C依賴性的磷酸化位點。目前已知,氨基酸的跨膜轉運是由腸上皮細胞刷狀緣的膜轉運系統來完成的。通過這些膜轉運系統,氨基酸底物才能夠從細胞膜的一側轉運至另一側,從而執行其廣泛而又復雜的生理功能。研究已經證實,小腸上皮細胞刷狀緣表達 ASCT2是一種廣譜的中性氨基酸載體,主要吸收谷氨酰胺、丙氨酸、絲氨酸和半胱氨酸等中性氨基酸,為細胞代謝提供重要的營養底物。與 Na+非依賴性氨基酸轉運系統相比,Na+依賴性氨基酸轉運系統由于可利用細胞膜兩側 Na+電勢梯度和逆濃度梯度轉運氨基酸,在腸腔內對氨基酸的吸收起著中心作用[38]。
3 小 結
維持細胞中谷胱甘肽含量是谷氨酸轉運的動力,而由于半胱氨酸為谷胱甘肽合成中谷氨酸循環限速酶的底物,谷氨酸轉運又受到半胱氨酸的限制。
[1] REEDS JP,BURRIN G D,STOLL B,et al.Intestinal g lutamate metabolism[J].Journal of Nutrition,2000,130:978-982.
[2] FAN M Z,MATTHEW SC J,ETIENNEM P N,et al.Expression of apicalmembrane L-glutamate transporters in neonatal porcine epithelial cells along the small intestinal crypt-villus axis[J].American Journal of Physiology Gastrointestinal Liver Physio logy,2004,287:385-398.
[3] SANTOKH G,PULIDO O.G lutamate receptors in peripheral tissue:excitatory transm ission outside the CNS[M].London:K luwer Academ ic/Plenum Publishers,2005:47-48.
[4] AOYAMA K,W ATABE M,NAKAKI T.Regulation of neuronal glutathione synthesis[J].Journal of Pharmaco logy Science,2008,108:227-238.
[5] HINOI E,TAKARADA T,UNO K,et al.G lutamate suppresses osteoclastogenesis through the cystine/glutamate antiporter[J].American Journal of Pathology,2007,4(170):1 277-1 290.
[6] KANAIY,HEDIGER A M.The glutamate/neutral am ino acid transporter fam ily SLC 1:molecular,physio logical and pharmaco logical aspects[J].European Journal o f Physiology,2004,447:469-479.
[7] OPPEDISANO F,POCHINIL,GALLUCCIOM,et al.The glutam ine/am ino acid transporter(ASCT2)reconstituted in liposomes:transportmechanism,regulation by ATP and characterization of the glutam ine/glutamate antiport[J].Biochem istry Biophysical A cta,2007,1768:291-298.
[8] SHANKER G,ASCHNER M.Identification and characterisation of uptake systems for cystine and cysteine in cu ltured astrocy tes and neurones:evidence for methylmercury-targeted disruption of astrocyte transport[J].Journal of Neuroscience Research,2001,66:998-1002.
[9] CHO Y,BANNAIS.Uptakeof glutam ate and cysteine in C-6 glioma cells and in cultured astrocy tes[J].Journalo f Neurochem istry,1990,55:2091-2097.
[10] MCBEAC JG.Cerebral cystine uptake:a tale of tw o transporters[J].TRENDS in Pharmacology Science,2002,23(7):299-303.
[11] BODE B P.Recentmo lecular advances in mammalian g lutam ine transport[J].Journal of Nutrition,2001,131(9):2475-2485.
[12] 周濟宏,李幼生,洪志堅,等.腸黏膜上皮細胞的載體分布及功能[J].醫學研究生學報,2009,22(7):677-681.
[13] CAROZZIV A,CANTA A,OGGIONI N,et al.Expression and distribution of'high affinity'glutamate transporters GLT1,GLAST,EAAC 1 and of GCPⅡin the ratperipheralnervous system[J].Journalof A-natomy,2008,213:539-546.
[14] AOYAMA K,SUH S W,HAMBY A M,et al.Neuronal glutathione deficiency and age-dependent neurodegeneration in the EAAC1 deficientmouse[J].National Neuroscience,2006,9:119-126.
[15] BURDO J,DARGUSCH R,SCHUBERT D.D istribution of the cystine/glutamate antiporter system X-Cin the brain,kidney,and duodenum[J].Journal o f Histochem istry and Cytochem istry,2006,54(5):549-557.
[16] YAMAMOTO T,N ISHIZAK I I,FURUYA S,et al.Characterization of rapid and high-affinity uptake of L-serine in neurons and astrocytes in primary culture[J].FEBS Letter,2003,548:69-73.
[17] BROER A,WAGNER C,LANG F,et al.Neutral am ino acid transporter ASCT2 displays substrate-induced Na+exchange and a substrate-gated anion conductance[J].Biochem istry Journal,2000,346:705-710.
[18] CONTI F,DEBIASI S,M INELLI A,et al.EAAC 1,a high-affinity glutamate transporter,is localized to astrocytes and gabaergic neurons besides pyram idal cells in the rat cerebral cortex[J].Cerebral Cortex M ar,1998,8:108-116.
[19] XIA P,PEI G,SCHWARZ W.Regulation of the glutamate transporter EAAC1 by exp ression and activation of delta-opioid receptor[J].European Journal of Neuroscience,2006,24:87-93.
[20] KAVANAUGH M P,BENDAHAN A,ZERANGUE N,et al.Mutation of an am ino acid residue influencing potassium coup ling in the g lutamate transporter GLT-1 induces obligate exchange[J].Journal of Biology Chem istry,1997,272:1703-1708.
[21] ARRIZA J L,ELIASOF S,KAVANAUGH M P,et al.Excitatory am ino acid transporter 5,a retinal glutamate transporter coupled to a ch loride conductance[J].Proceeding of the National Academy Sciences,1997,94:4155-4160.
[22] TANAKA K.Ro le o f glutamate transporters in astrocytes[J].Brain Nerve,2007,59(7):677-688.
[23] HOWELL J A,MATTHEWS A D,SWANSON K C,et al.Molecular identification of high-affinity glutamate transporters in sheep and cattle forestomach,intestine,liver,kidney,and pancreas[J].Journal o f Animal Science,2001,79:1329-1336.
[24] HIM IT,IKEDA M,YASUHARA T,etal.Role of neuronal glutamate transporter in the cysteine uptake and intracellular g lutathione levels in cultured cortical neurons[J].Journal of Neural Transm ission,2003,110:1337-1348.
[25] K IYAMA H,K IRYU-SEO S.Mu ltiple functions of glutamate transporter EAAC 1 in motor neurons[J].Brain Nerve,2007,59(12):1325-1332.
[26] BEART PM,OHEA R D.Transporters for L-glutamate:An updateon theirmolecular pharmaco logy and patho logical involvement[J].British Journal o f Pharmacology,2007,150:510-517.
[27] REXHEPAJ R,GRAHAMMER F,VOLKL H,et al.Reduced intestinal and renal am ino acid transport in PDK 1 hypomorphic m ice[J].FASEB Journal,2006,20:2214-2222.
[28] WATABE M,AOYAMA K,NAKAK I T.Regulation of glutathione synthesis via interaction betw een glutamate transport-associated protein 3-18(GTRAP3-18)and excitatory am ino acid carrier-1(EAAC 1)at plasmam em brane[J].Molecular Pharmacology,2007,72:1103-1110.
[29] BUTCHBACH M E,GUO H,LIN C L.M ethy l-beta-cyclodextrin but not retinoic acid reduces EAAT3-mediated glutamate up take and increases GTRAP3-18 exp ression[J].Journal o f Neurochem istry,2003,84:891-894.
[30] RUGGIERO A M,LIU Y,V IDENSKY S,et al.The endop lasm ic reticulum exitofglutamate transporter is regulated by the inducible mammalian Y ip6b/GTRAP3-18 p rotein[J].Journal of Biology Chem istry,2008,283:6175-6183.
[31] HUANG Y,FENG X,SANDO J J,et al.Critical role of serine 465 in isoflurane-induced increase of cell-surface redistribution and activity of glutamate transporter type 3[J].Journal of Bio logical Chem istry,2006,281:38133-38138.
[32] SHELDON A L,GONZALEZ M I,ROBINSON M B.A carboxy l-term inal determ inant of the neuronal glutamate transporter,EAAC 1,is required for platelet-derived grow th factor-dependent trafficking[J].Journal of Biology Chem istry,2006,281:4876-4886.
[33] GONZALEZM I,KAZANIETZ M G,ROBINSON M B.Regulation of the neuronal glutamate transporter excitatory am ino acid carrier-1(EAAC 1)by different protein kinase C subtypes[J].Molecular Pharmacology,2002,62:901-910.
[34] KALANDADZEA,WU Y,ROBINSON M B.Pro-tein kinase C activation decreases cell surface expression of the GLT-1 subtype of glutamate transporter:requirement of a carboxyl-term inal domain and partial dependence on serine 486[J].Journal of Biology Chem istry,2002,277:45741-45750.
[35] BANNA IS,K ITAMURA E.Transport interaction o f L-cystine and L-glutamate in human dip liod fibroblasts in cu lture[J].Journal of Biological Chem istry,1980,255:2372-2376.
[36] UTSUNOM IYA-TATE N,ENDOU H,KANAI Y.Cloning and functional characterization of a system ASC-like Na+-dependentneutral am ino acid transporter[J].Journal of Biology Chem istry,1996,271:14883-14890.
[37] WANG X,HALD H,ERNST H A,et al.Over-expression,purification and characterization of an Asc-1 homologue from G loeobacter violaceus[J].Protein Expression and Purification,2010,71:179-183.
[38] AVISSAR N E,RYAN C K,GANAPATHY V,et al.Na(+)-dependent neutral am ino acid transporter ATB(0)is a rabbit epithelial cell brush-border p rotein[J].American Journal of Physio logy,2001,281:963-971.
[39] DRINGEN R.Metabo lism and functions o f glutathione in brain[J].Progress in Neurobiology,2000,62:649-671.
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
