原核表達載體pET28a-EGFP的構建與表達
2011-01-12 07:22:54季愛加寧喜斌
微生物學雜志 2011年4期
季愛加,寧喜斌
(上海海洋大學食品學院,上海 201306)
綠色熒光蛋白(Green fluorescent protein,GFP)是由日本學者下村修[1]于1962年從多管水母(Aequorea victoria)中發現并分離得到的一種發光蛋白。分子質量為26 ku,是由238個氨基酸構成的“β-桶”型三維立體結構,其中第65~67位氨基酸(絲氨酸-酪氨酸-甘氨酸)形成發光團,為主要發光的位置。GFP的結構賦予了其一些獨特的性質[2]:熒光穩定、檢測方便、無種屬特異性、不需任何反應底物和輔助因子、易于構建載體,并且它不會破壞細胞生長環境,對細胞生理功能無影響等,因此是常用的報告基因。目前,科研人員根據不同需要,利用氨基酸序列修飾等方法對GFP進行改進,獲得了一系列優良的變異體,發光的強度和顏色都有變化,如苯丙氨酸64被亮氨酸取代,絲氨酸65被蘇氨酸取代的EGFP熒光強度就增加了35倍。而且出現了紅色、黃綠色、藍色等多種顏色的熒光蛋白,大大拓寬了GFP研究的領域[3]。近些年,國內外掀起了關于GFP研究的熱潮,已見GFP應用于各人源細胞、動物細胞、植物細胞、真菌等領域中[4-7]。本研究以原核生物E.coliBL21為表達宿主,利用PCR技術從模板EGFP-N3中克隆得到EGFP片段,并在其2端引入酶切位點EcoRⅠ和HindⅢ,對引入酶切位點的EGFP片段和pET28a質粒進行雙酶切處理,連接后得到重組質粒pET28a-EGFP并轉化至E.coli BL21中,通過IPTG誘導可以高效表達。食品安全問題一直是困擾各國的公共衛生問題,食源性病原菌又是引起食品安全問題的主要因素之一。通過本研究,期望能夠為以后EGFP作為熒光標記物標記食源性病原菌,跟蹤監測病原菌的侵染路徑提供理論基礎,從而為食品安全風險評估等提供合理依據。
1 材料與方法
1.1 材料
1.1.1 試驗材料與試劑 PEGFP-N3模板、菌株E.coliDH5α(用于質粒的保存)、E.coliBL21(用于重組質粒的表達)、質粒pET28a(復旦大學遺傳所);限制性內切酶EcoRⅠ,HindⅢ(紐英倫生物技術有限公司);T4DNA連接酶、1 kb DNA ladder(Fermentas公司);DNA凝膠回收試劑盒及質粒小提試劑盒(愛思進生物技術公司);DNA純化試劑盒及IPTG(大連寶生物工程有限公司);PCR用試劑(北京全式金生物技術公司);卡那霉素、瓊脂糖及PCR合成引物等(上海生工生物工程技術服務有限公司);蛋白胨、酵母浸出粉、瓊脂粉等(國藥集團化學試劑有限公司)。
1.1.2 主要儀器和設備 PTC-200 PCR儀(美國Bio-Rad公司);GelDoc XR凝膠成像系統(美國Bio-Rad公司);Carl Zeiss Axio-Scope.A1落射熒光顯微鏡(德國Zeiss Jena);DYY-6C電泳槽和電泳儀(北京六一儀器廠);TGL-16G高速臺式離心機(上海安亭科學儀器廠);UV2300紫外可見分光光度計(上海天美科學儀器有限公司)。
1.2 方法
1.2.1 EGFP基因的PCR擴增和回收 以PEGFP-N3中EGFP片段為模板,設計上游引物:5′-ATGGTGAGCAAGGGCGAGGAG-3′,下游引物:5′-GGCTGATTATGATCTAGAGTC-3′。分別取4.0 μL模板、2μL 10μmol/L的上游和下游引物、1.0 μLTransStart FastPfuDNA polymerase(5 U/μL)、10μL 10×Buffer、5.0μL dNTPMixture作為PCR反應體系。用無菌水將反應液調至50μL,于95℃條件下預變性處理2 min,然后分別在94℃變性20 s、65℃退火20 s、72℃延伸30 s、最終72℃終延伸5 min,整個PCR反應體系共進行35個循環。擴增產物經1.0%的瓊脂糖凝膠電泳驗證后,用DNA純化試劑盒回收后待用。
1.2.2 EGFP基因和pET28a表達載體的限制性酶切處理 取上述經DNA純化試劑盒純化后的目的片段作為PCR反應的模板,設計引入EcoRⅠ酶切位點的上游引物(5′-GGAG/AATTCATGGTGAGCAAGGGCGAGGAG-3′)和引入HindⅢ酶切位點的下游引物(5′-CCGA/AGCTTGGCTGATTATGATCTAGAGTC-3′)。PCR反應體系及反應條件同1.2.1。擴增產物經1%瓊脂糖凝膠電泳檢驗后,進行DNA純化。純化后的EGFP片段進行雙酶切[8],反應體系如下:30 μL添加了酶切位點的EGFP片段、5μL 10×buffer2、1μLEcoRⅠ(20 U/μL)、1μLHindⅢ(20 U/μL),用無菌水將反應液調至50μL后,于37℃下保溫1 h,然后在65℃條件下處理20 min進行熱失活。經1.0%瓊脂糖凝膠電泳檢驗后,DNA純化試劑盒純化回收。另一方面,將pET28a質粒載體轉化大腸埃希菌DH5α,培養過夜后提取質粒,并進行雙酶切,經1.0%瓊脂糖凝膠電泳檢驗后,切下線性質粒條帶進行純化回收。
1.2.3 pET28a-EGFP重組表達載體的構建 將上述含酶切位點的EGFP片段與線性pET28a質粒按2◇1比例、在45℃下保溫5 min后,置于冰浴中冷卻,冷卻后分別加入1μL T4DNA連接酶、2μL 10×T4DNA連接酶緩沖液,16℃保溫16 h,如圖1所示。連接液轉化大腸埃希菌感受態細胞DH5α,經37℃培養過夜后,挑選3~5個菌落做PCR驗證,進一步進行酶切鑒定,陽性克隆用于DNA測序。對經測序確證的、含pET28a-EGFP重組質粒的菌落進行質粒提純。
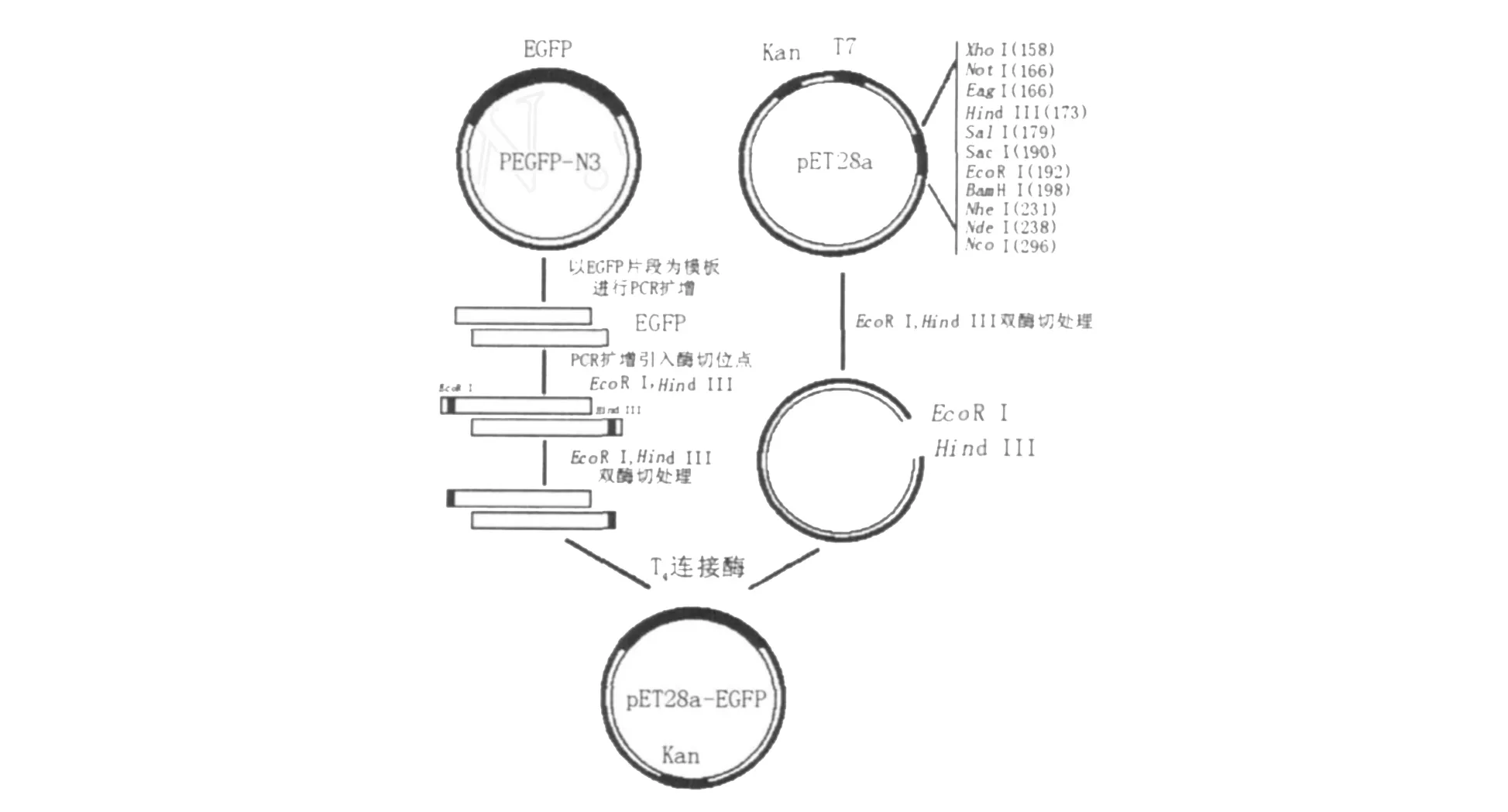
圖1 pET28a-EGFP重組表達載體的構建圖譜Fig.1 Construction of recombinant expression vector pET28a-EGFP
1.2.4 EGFP基因的原核表達及檢測 取1μL“pET28a-EGFP”重組表達質粒,轉化至E.coli BL21感受態細胞,經37℃培養過夜后。隨機挑選10個陽性菌落克隆于含卡那霉素(50μg/mL)的LB液體培養液中,37℃、220 r/min搖床培養至OD600=0.4時,加入誘導劑IPTG,并使其終濃度為1.0 mmol/L。將含有誘導劑的培養液于25℃、110 r/min條件培養12 h后,制成玻片于熒光顯微鏡下觀察,對在熒光顯微鏡下發綠光的大腸埃希菌及時保種,同時,劃線至含卡那霉素(50 μg/mL)及IPTG(1 mmol/L)的LB固體培養基,培養12 h,觀察菌落形態。
2 結果與分析
2.1 pET28a-EGFP重組表達載體的構建及鑒定
將EGFP片段與pET28a表達載體經連接液連接后,通過對挑取的單菌落做菌落PCR檢驗,如圖2所示,PCR產物經1%瓊脂糖凝膠電泳后,發現在約750 bp處有清晰條帶(圖2左),與理論相符,初步確定為陽性克隆。進一步對陽性菌落中的質粒進行提純、用EcoRⅠ和HindⅢ對質粒進行雙酶切,經1.0%瓊脂糖凝膠電泳后,可在約750 bp處看到目的片段的條帶(圖2右)。最終將PCR產物送往上海美吉生物技術公司進行測序,通過與GenBank序列比對,結果顯示序列匹配率為100%,由此可以證明目的片段“EGFP”已正確連接到pET28a表達載體,并且核苷酸序列未發生任何堿基突變。
2.2 重組表達載體pET28a-EGFP在E.coli BL21中的表達與檢測
盡管目的片段EGFP與表達載體pET28a得到了有效重組,但重組載體pET28a-EGFP是否能在原核系統中得到高效表達還需進一步研究,因此通過利用pET28a-EGFP轉化E.coliBL21感受態細胞進行檢測。在含卡那霉素的LB培養基中篩選陽性克隆,LB液體培養基擴大培養至OD600=0.4時,加入IPTG(1 mmol/L)誘導劑誘導表達12 h,取少量菌液涂片,在熒光顯微鏡(10×100)下可以明顯觀察到發綠光的單個細菌,將轉化成功的E.coliBL21命名為E.coliBL21-1,如圖3所示。而轉化菌劃線于含卡那霉素(50μg/mL)與IPTG(1 mmol/L)的LB固體培養基,培養24 h后,在自然光下菌落呈綠色如圖4所示,因此可以說明,重組表達載體pET28a-EGFP能夠在原核系統中得到高效的表達,并且表達效果較好。
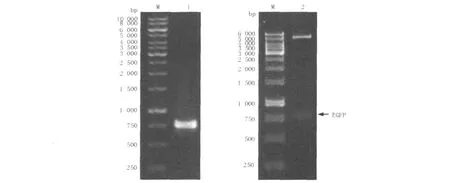
圖2 重組質粒的PCR插入檢驗(左)和雙酶切鑒定(右)Fig.2 Identification of recombinant plasmid by PCR amplification(left)and digesting with restriction enzymes(right)
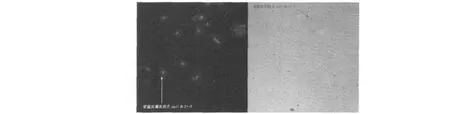
圖3 E.coliBL21-1在藍光激發下(左)和在自然光下(右)觀察的結果Fig.3 Expression of EGFP inE.coliBL21-1 in the blue light(left)and in natural light(right)
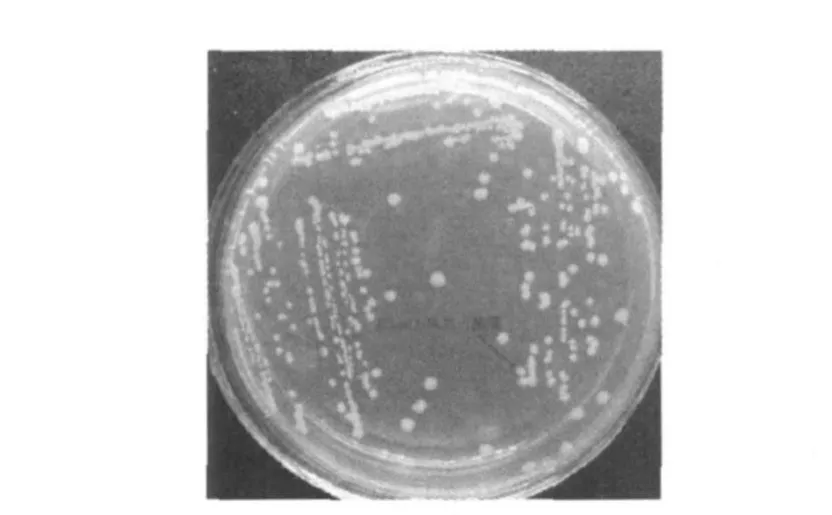
圖4 E.coliBL21-1菌落在自然光下形態Fig.4 Colonies of E.coliBL21-1 in natural light
3 討 論
外源基因的誘導表達常常對宿主造成很強的代謝負擔,表達的目的蛋白有時會對宿主菌有“毒性”作用。而GFP恰恰克服了這一障礙,大部分研究認為作為分子標記物,GFP對細胞無毒性且對細胞生長代謝和細胞功能均無影響,無疑成為眾多科研人員的理想選擇。
本實驗選擇的EGFP是野生型GFP的改造型,其所產生的熒光比野生型GFP亮35倍,可以顯著提高其檢測的靈敏度;同時本研究中選擇pET28a質粒作為表達載體,該質粒含有強啟動子T7啟動子,可以使目的基因得到高效表達。以上表明,pET28a-EGFP重組表達載體能使EGFP基因和pET28a質粒充分發揮各自的優良特性。若GFP直接作為熒光標記物標記食源性病原菌,不僅可以克服熒光染料分布不均勻,易滲漏,隨細胞分裂被稀釋的缺點[9],也易于進行跟蹤監測。
目前,國內關于GFP的研究主要集中在醫學領域[10-12],在食品科學相關領域的研究很少,國外有將GFP標記食源性病原菌對其進行跟蹤監測的報道[13]。本實驗將EGFP片段與pET28a載體連接,獲得重組質粒pET28a-EGFP并轉化至E.coliBL21工程菌中,GFP在自然光及熒光顯微鏡下均得到高效表達,為以后標記其他食源性病原菌并對其進行跟蹤監測提供了理論依據。
[1] Shimomura O,Johnson FH,Saiga Y.Extraction,purification and properties of aequorin,a bioluminescent proteinfrom the luminous hydromedusan,Aequorea[J].J.Cell.Comp.Physiol.,1962,59(3):223-239.
[2] 吳沛橋,巴曉革,胡海,等.綠色熒光蛋白GFP的研究進展及應用[J].生物醫學工程研究,2009,28(1):83-86.
[3] 馬金石.綠色熒光蛋白[J].化學通報,2009,72(3):243-250.
[4] 代文杰,呂曉穎,周保國,等.人源化綠色熒光蛋白基因真核表達載體的構建與表達檢測[J].肝膽胰外科雜志,2002,14(4):213-214.
[5] Park K W,Cheong HT,LaiL,et al.Production of nuclear transfer derived s wine that express the enhanced green fluorescent protein[J].AniBiotech,2001,12(2):173-181.
[6] Skadsen RW,Hohn T M.Use of Fusarium graminearum transformed with gfp to follow infection patterns in barley and Arabidopsis[J].Physiological and Molecular Plant Pathology,2004,64(1):45-53.
[7] Khodjakov A,Terra SL,Chang F.Laser microsurgery in fission yeast:role of the mitotic spindle midzone in Anaphase B[J].Current Biology,2004,14(15):1330-1340.
[8] Sambrook J,Fritsch EF,Maniatis T.分子克隆實驗指南[M].北京:科學出版社,1992:55-57.
[9] 張國增,李瑞玲,王強.熒光染料在植物細胞Ca2+濃度測定中的應用[J].中國農學通報,2010,26(9):198-201.
[10] 郝鋼躍,張維東,張月英,等.穩定高表達增強型綠色熒光蛋白基因膀胱癌細胞株的構建[J].山東醫藥,2009,49(46):1-3.
[11] 王婧,李昌,李林溪,等.反轉錄病毒介導的穩定表達H IV-1 Gag蛋白和增強型綠色熒光蛋白細胞系的建立[J].畜牧獸醫學報,2011,42(2):203-209.
[12] 金鷹,邢達.腫瘤細胞中表達的GFP蛋白的熒光漂白特性的研究[J].光譜與光譜分析,2004,24(12):1626-1629.
[13] Alicia EC,Romilio TE,Jaime R.TracingVibrio parahae molyticusin oysters(Tiostrea chilensis)using a Green Fluorescent Protein tag[J].J.Exp.Mar.Biol.Ecol.,2005,327(2):157-166.
