大型蜥腳類恐龍研究
2010-12-21 10:48:20錢邁平胡柏祥詹庚申鄒松梅章其華
地質學刊 2010年4期
錢邁平,胡柏祥,詹庚申,鄒松梅,章其華
(1.南京地質礦產研究所,江蘇 南京 210016;2.江蘇省地質調查研究院,江蘇 南京 210018)
大型蜥腳類恐龍研究
錢邁平1,胡柏祥2,詹庚申2,鄒松梅2,章其華2
(1.南京地質礦產研究所,江蘇 南京 210016;2.江蘇省地質調查研究院,江蘇 南京 210018)
大型蜥腳類恐龍是地球歷史上最大的陸地動物,其中已知體形最大者是梁龍,身長可達 54m,所有已知身長超過 20m的恐龍都屬于蜥腳亞目。蜥腳類恐龍向大型化演化是為了占據更好的生態位空間,取得生存競爭的優勢,其他演化都是圍繞這一演化主線的配套演化。如:①小頭長頸,是為了不用過多移動龐大的身軀就能在大范圍內覓食。②大鼻孔,是為了增大呼吸吞吐量,供應龐大身軀的生命活動,由此可推想其心肺功能的強大,甚至可能除了一個主心臟外,還可能有多個小型輔助“心臟”,以免單個心臟過于龐大。③長形的葉狀牙齒,是為了快速連續地進食,力求每次都能撕扯下一大片鮮嫩的植物枝葉,不經咀嚼直接吞咽。④股骨比脛骨長,說明其雖不善于奔跑,但后肢支撐力很強,有時可站立起來吃高處的植物,或用前肢撲打敵害進行自衛。也說明其不是靠快速奔跑擺脫敵害,而是依仗龐大身軀及強大力量對抗敵害。⑤四足行走,是為了把龐大身體的重量分散到 4個寬大的腳掌上,減小對地面的壓強,提高通過松軟地面的能力。由此可見,生物演化對其自身的生理和身體結構是牽一發而動全身的,演化的主線會帶動一系列配套的演化,以求盡量達到對環境的最佳適應。
蜥腳類恐龍;大型化;中生代
0 引 言
恐龍是中生代地球生物圈的主角之一,它們根據腰帶 (即骨盆)構造的不同分為蜥臀目 (Saurischia)和鳥臀目 (Ornithischia),蜥臀目又進一步分為蜥腳亞目 (Sauropodomorpha)和獸腳亞目 (Theropoda)。蜥腳亞目顧名思義就是有“蜥蜴般的腳”的一類恐龍,迄今發現的化石已超過 100種,包括兩大類——原蜥腳下目 (Prosauropoda)和蜥腳下目 (Sauropoda)。
原蜥腳下目主要有:里奧哈龍科 (Riojasauridae)、大椎龍科 (Massospondylidae)和板龍科 (Plateosauridae)等。
蜥腳下目主要有:鯨龍科 (Cetiosauridae)、火山齒龍科 (Vulcanodontidae)、叉龍科 (Dicraeosauridae)、梁龍科 (Diplodocidae)、雷巴齊斯龍科 (Rebbachisauridae)、腕龍科 (Brachiosauridae)、圓頂龍科(Camarasauridae)、盤足龍科 (Euhelopodidae)和泰坦巨龍類 (Titanosaur)等。
雖然地球陸地上的生命出現已超過 4億年,但除了大型蜥腳類恐龍外,還沒有身長超過 20m的動物,所以它們的演化特別引起科學家們的關注。來自化石的信息顯示,大型蜥腳類恐龍的特征如下:①小頭長頸。②大鼻孔。③長形的葉狀牙齒。④股骨比脛骨長。⑤四足行走。⑥素食。
因以植物為食,食物的熱量和營養較低,需要巨大的食量才能維持日常活動所需的能量,所以它們生活中的大部分時間都用于進食,胃容量很大,腸子也很長,這樣才能有效消化大量的食物。因此,它們身軀特別粗大,以容納龐大的腸胃系統。為了支撐沉重碩大的身軀,它們的四肢粗狀,腳掌寬大以減小壓強,防止陷入松軟的地面。它們通常以群居方式生活,抵御掠食動物襲擊主要依靠龐大的身軀和力量,而不是依靠敏捷和快速的逃避。其覓食、生長、繁衍和遷徙的行為方式和群體結構,類似現代大型哺乳動物,如非洲象 (Loxodonta africana)、斑紋角馬(Connochaetes taurinus)和美洲野牛 (B ison bison)等。
由于陸地動物的遺骸通常不容易被沉積物快速封埋,保存成化石的難度遠超過水里的生物。大型蜥腳類恐龍雖然體形巨大,但一方面為了節省能量和資源,另一方面為了不使行動過于笨拙,它們的骨骼已演化成多孔、空心或薄壁結構,以減輕重量,死亡后很容易在外力作用下破碎,經過漫長的地質作用后,很難保存完整的化石。這就是為什么迄今發現的大型蜥腳類恐龍 95%以上的化石標本是嚴重破碎殘缺的,特別是薄殼結構的頭骨更加難以保存。所以,較完整的大型蜥腳類恐龍化石就顯得格外珍貴,成為科學家探索這些遠古時期巨型陸地動物如何起源演化,繁衍生息以及如何從興旺走向衰亡的最直接的實際材料。
1 化石記錄
雖然迄今已發現的恐龍超過1 000個屬,但身長20m以上的巨型恐龍屬并不多,幾乎都是蜥腳亞目的,根據體形大小排列如下。
1.1 梁龍 (D iplodocus)
1878年命名,屬梁龍科,按以往的資料推算身長可達 27m,體重可達 20t,可能是目前已知最大的恐龍。
1991年命名的地震龍 (Seism osaurus),推算身長37m~54m,體重 30t~70t。但后來的研究表明,這只是長得過于龐大的梁龍而已 (圖 1)。其標本于1979年在美國新墨西哥州沙漠上侏羅統莫里遜組(Morrison For mation)地層發現,已采集到尾椎、腰帶、背椎、肋骨及頸椎骨骼化石。還有一些被認為是胃石的卵石,但也可能是河流沖刷形成的普通卵石。

圖1 “地震龍 (Seism osaurus)”(梁龍)骨架復原
特征:長頸、長尾、小頭。頸椎骨 15個,背椎骨 10個,尾椎骨約 70個。頭骨長,嘴前部長有扁平牙齒,側部和后部無牙齒。四肢粗壯,前腿比后腿短,臀部高于肩部。每只腳五趾,前腳內側其一趾長有大而彎曲的利爪 (圖 2)。盡管梁龍科恐龍骨架巨大,但結構較輕,體重比同樣大小的泰坦巨龍類的明顯要輕。
時代:晚侏羅世 Kimmeridgian-Tithonian期,156Ma~145Ma。
標本:在美國科羅拉多、蒙大拿、猶他和懷俄明州上侏羅統陸續發掘出多具不同保存程度的骨架化石,包括珍貴的頭顱骨化石也已找到,研究程度較高。

圖2 梁龍 (D iplodocus)骨架復原
1.2 巨體龍 (B ruhathkayosaurus)
1989年命名,屬泰坦巨龍類,推算身長 40m~44m,重 175t~220t。可能是目前已知最重的恐龍 。
特征:長頸、長尾、小頭,體態類似腕龍 (B rachiosaurus)。其脛骨長 2m,比阿根廷龍 (A rgentinosaurus)的 1.55m長 29%;肱骨長 2.34m,比阿根廷龍(Argentinosaurus)的 1.81m長 30%(圖 3)。
時代 :晚白堊世 Maastrichtian期,約 70Ma。
標本:發現于印度南部的泰米爾納德邦上白堊統卡勒梅德組 (Kallemedu Formation)地層,化石包括腸骨和坐骨的臀部骨骼、1個殘缺的股骨、1個脛骨、1個橈骨以及 1個尾椎骨。最初被鑒定為獸足亞目的食肉恐龍,后重新研究發現是蜥腳亞目的素食恐龍。

圖3 巨體龍 (B ruhathkayosaurus)復原
1.3 普爾塔龍(Puertasaurus)
2005年命名,屬泰坦巨龍類,見于晚白堊世早Maastrichtian阿根廷巴塔哥尼亞 (約 70Ma),正型標本包含 4節頸椎、背椎以及尾椎,其背椎長 1.06m,寬 1.68m,是目前已知最寬的蜥腳類脊椎骨。推測身長達 35m~40m,重約 80t~100t(圖 4)。

圖4 普爾塔龍 (Puertasaurus)背椎化石 (右下)和體態復原圖
1.4 超龍 (Supersaurus)
1985年命名,屬梁龍科,推算身長 27m~40m,體重 40t~70t。
特征:長頸、長尾、小頭,體態類似迷惑龍 (Apatosaurus)。其肩胛骨長 2.4m,肋骨長 3.1m,骨盆寬1.8m,單個頸椎骨可長達 1.5m(圖 5)。
時代:晚侏羅世 K immeridgian-Tithonian期,156Ma~145Ma。
標本:1972年發現于美國科羅拉多上侏羅統莫里遜組 (Morrison Formation)地層,僅 1個肩胛骨和幾個頸椎骨化石。后又在懷俄明州康維斯縣 (Converse County)發現了更完整的標本,被懷俄明恐龍中心 (Wyoming Dinosaur Center)收藏。

圖5 超龍 (Supersaurus)肩胛骨化石 (上)和生態復原圖 (下)
1.5 阿根廷龍(A rgentinasaurus)
1993年命名,屬泰坦巨龍類,推算身長 27m~35m,體重 80t~100t。
特征:其背椎長 1.3m,脛骨長 1.55m,肱骨長1.81m。脊椎有巨大的側翼,適合附著大塊的肌肉(圖6)。

圖6 阿根廷龍 (A rgentinasaurus)骨架復原模型
時代:白堊紀 Albian-Cenomanian期 (112.2 Ma~93.5Ma)。
標本 :發現于阿根廷 Neuquén省 Río Limay組地層,已發掘出一些背椎、脛骨、破碎的肋骨,還有薦骨。
1.6 馬門溪龍 (M am enchisaurus)
1954年命名,屬梁龍科,推算身長 22m~35m,體重 10t~20t,廣泛分布在東亞地區。中國四川、云南、甘肅和新疆都有化石發現,是迄今在中國發現的最大恐龍。
特征:脖子占身長的一半,頸椎長而細,頸部并不靈活,主要由與頭部、肩部的關節來動作、彎曲。馬門溪龍是曾經生活在地球上的脖子最長的動物之一 (圖 7)。

圖7 馬門溪龍 (M am enchisaurus)骨架復原模型
時代:晚侏羅世 Tithonian期 (150Ma~140Ma)。
標本:1952年首次在四川宜賓的馬鳴溪渡口旁的公路建設工地上,發現部分骨骼化石。1954年被楊鐘健描述、命名,由于方言口音被誤作為馬門溪龍。因發現于建設工地,故模式種命名為建設馬門溪龍 (M.constructus)。該標本身長 22m,頸部長度為 11m。另一標本發現 19個頸椎骨,上有長條骨棒。其成年個體身長平均約 24m。
1972年發現第二個種,發現于云南合川,命名合川馬門溪龍(M.hochuanensis),包含 4個不完整骨架 ,頸長達 9.5m。
1994年發現的中加馬門溪龍 (M.sinocanadorum)身長可能達 27m。其頸肋長達 41cm,長于波塞東龍的 34.2cm,是目前蜥腳下目中最長的。
1994年在美國發現的波塞東龍,頸部被證實長達 10.5m~11.5m。波塞東龍屬于腕龍科,有較長的前肢,高度比馬門溪龍高許多。而之前發現的超龍,頸部長度則為 13m~4m。
2006年出土的新疆馬門溪龍,長 35m,頸部長15m,是目前中國最大的恐龍。
1.7 巨酋龍 (Futalognkosaurus)
2007年命名,屬泰坦巨龍類。生存于阿根廷晚白堊世 (約 88Ma),標本為近乎完整的骨架化石,身長 32m~34m(圖 8)。
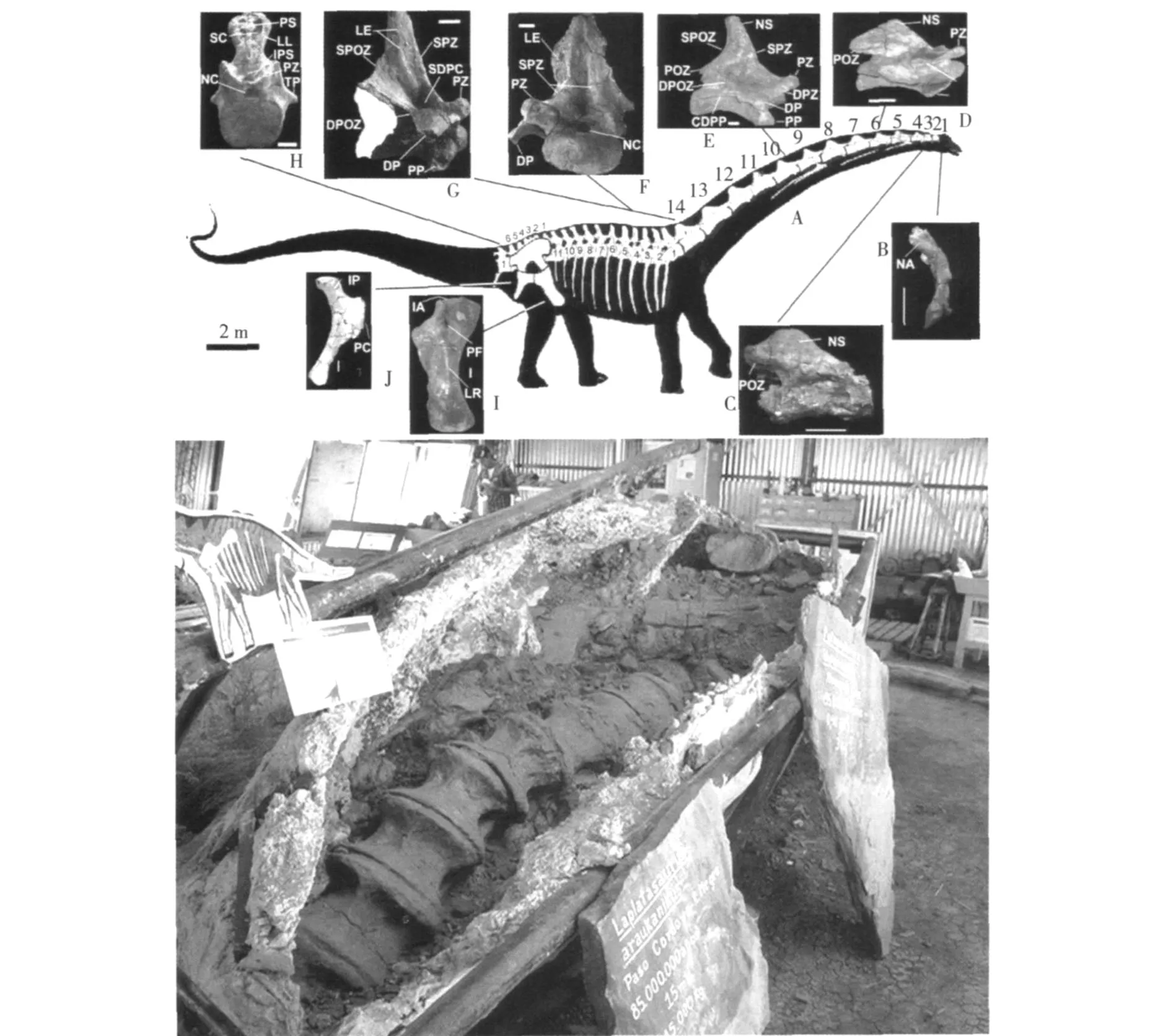
圖8 巨酋龍 (Futalognkosaurus)骨骼化石
1.8 海潮巨龍 (Paralititan)
2001年命名,屬泰坦巨龍類,發現于埃及白堊紀 (約 94Ma)濱海沼澤相沉積中。標本為零散骨骼化石,肱骨長 1.69m,推算身長 24m~32m,體重 65t~80t(圖 9)。

圖9 海潮巨龍 (Paralititan)肩胛骨化石 (上)和生態復原圖 (下)

圖10 重龍 (Barosaurus)化石骨架復原 (上)和生態圖 (下)
1.9 重龍 (B arosaurus)
1890年命名,屬梁龍科。推算身長 23m~27m,體重 10t。
特征:類似梁龍 (D iplodocus),但頸很長尾卻較短,背椎骨也不同 (圖 10)。
時代:晚侏羅世 Kimmeridgian期,154.1Ma~150.7Ma。
標本:最初在美國南達科他州黑山 (Black Hills)上侏羅統莫里遜組 (Morrison Formation)層位,發現一串相聯的尾椎骨化石,此后又陸續找到一些化石;1922年在尤他州的 Carnegie Quarry發現 3個較完整骨架化石。另外,2007年加拿大皇家多倫多博物館在庫存標本中,意外發現 1具 24m長的部分完整骨架化石。在坦桑尼亞至少發現了 3個不完整骨架,包括頭骨碎片及顱后骨骼化石。
1.10 海神龍 (Sauroposeidon)
2000年命名,屬腕龍科,推算身長約 30m,體重10t,身高約 18m,可能是迄今發現的最高恐龍。
特征:類似腕龍 (B rachiosaurus),但更高大,單個頸椎長超過 1.4m(圖 11)。

圖11 海神龍 (Sauroposeidon)與腕龍 (B rachiosaurus)復原對比
時代 :白堊紀 (約 110Ma)。
標本:1994年由美國俄克拉荷馬州東南部的阿托卡縣 (Atoka County)州立監獄一訓狗員發現,目前僅找到第 5—8共 4個相連接的頸椎,并帶有頸肋。
1.11 銀龍 (A rgyrosaurus)
1893年命名,屬泰坦巨龍類,首見于阿根廷晚白堊世 (約 70Ma)。標本為零散骨骼化石,推算身長 20m~30m,體重 45t~55t(圖 12)。

圖12 銀龍 (A rgyrosaurus)骨骼化石 (左下)和體態復原圖
1.12 大夏巨龍 (Daxiatitan)
2008年命名,屬泰坦巨龍類,化石發現于蘭州盆地下白堊統河口群。
特征:股骨遠端髁突為 10°的對稱斜面,髁突的下中—上側部進入股骨骨干,顯示強烈的外八字步態。頸椎極大,推算身長約 28m,脖長 12m,身高8.4m,是中國發現的最大恐龍之一 (圖 13)。
時代:早白堊世。
標本:正型標本包括 10枚頸椎、10枚背椎,2枚近端尾椎、部分頸肋和背肋、1枚脈弓、右肩胛骨、右烏喙骨和右股骨。

圖13 大夏巨龍 (Daxiatitan)頸椎化石 (上)和生態復原圖 (下)
1.13 腕龍 (B rachiosaurus)
1903年命名,屬腕龍科,生存于美國科羅拉多和猶他晚侏羅世 (150Ma~145Ma)。標本陸續收集完整,包括珍貴的頭顱骨化石也已找到,研究程度較高。身長可達 25m,體重 32t~56t。因其頸椎結構可以向上豎起,使其身高可達 13m,是最高的恐龍之一(圖14)。
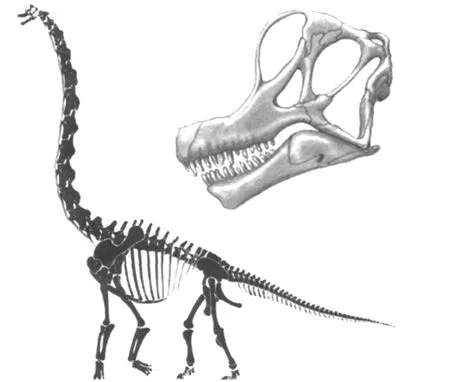
圖14 腕龍 (B rachiosaurus)骨架與頭骨復原圖
1.14 迷惑龍 (Apatosaurus)
1877年命名,屬梁龍科,曾誤名“雷龍 (B rontosaurus)”,見于美國懷俄明、科羅拉多、奧克拉荷馬和猶他晚侏羅世 (約 150Ma)。標本陸續收集完整,包括珍貴的頭顱骨化石也已找到,研究程度較高。身長平均 23m,體重至少 23t(圖 15)。

圖15 迷惑龍 (Apatosaurus)骨架復原 (上)和生態復原圖 (下)
1.15 圓頂龍 (Cam arasaurus)
1877年命名,屬圓頂龍科 (Camarasauridae),見于美國科羅拉多、新墨西哥、猶他和懷俄明晚侏羅世(150Ma~145Ma)。標本陸續收集完整,包括珍貴的頭顱骨化石也已找到,研究程度較高。身長可達23m,體重可達 47t(圖 16)。

圖16 圓頂龍 (Cam arasaurus)頭骨化石 (上)和骨架復原圖 (下)
1.16 江山龍 (Jiangshanosaurus)
2001年命名,屬泰坦巨龍類,化石見于中國浙江江山金華組下部地層 (105Ma),推測身長約 22m。正型標本包括:1個相當完整的左肩胛烏喙骨,5節后部和中部背椎骨,部分左右恥骨和坐骨,3節尾椎骨以及股骨殘片 (圖 17)。
1.17 黃河巨龍 (Huanghetitan)
2006年命名,屬泰坦巨龍類,見于中國甘肅、河南白堊紀。標本為不完整骨架化石,推算身長約20m,其肋骨最長約 3m,顯示體腔龐大,是體形最胖的恐龍 (圖 18)。
1.18 華北龍 (Huabeisaurus)
2000年命名,屬泰坦巨龍類,見于中國河北—山西晚白堊世 (約 75Ma)。標本為不完整骨架化石,推算身長約 20m(圖 19)。

圖17 江山龍 (Jiangshanosaurus)骨架復原模型
1.19 清秀龍 (Q ingxiusaurus)
1991年發現于中國廣西南寧市郊的上白堊統紅層,2008年命名,屬泰坦巨龍類,正型標本為一段較完整的前部尾椎神經棘、左右胸骨板、左右肱骨。推算身長約 20m。
2 研究進展
2.1 關于最大的恐龍
迄今的化石記錄證明,1979年發現于美國新墨西哥州沙漠的 1具不完整的梁龍科恐龍骨架化石,推算身長 37m~52m,是目前發現的最大恐龍。1991年美國南衛理公會大學 David Gillette發表的論文中,命名它為地震龍(Seism osaurus hallorum)。但許多科學家重新研究后認為,該龍骨骼的形態和梁龍屬 (D iplodocus)一致,脊椎錯置的屬征其實是病變或脊椎受傷錯位的結果。2004年美國地質協會年度會議上,地震龍這個屬名被廢棄,保留種名,歸類于梁龍屬,改名梁龍(D iplodocus hallorum)。但2006年發表的文章認為,連這個新種都不存在,坐骨末端鉤狀突起的種征,其實是埋藏時附著的椎弓。這樣看來,它其實屬于以前早已發現的長梁龍(D iplodocus longus)。所以,目前多數科學家認為,最大的恐龍就是梁龍。

圖18 黃河巨龍 (Huanghetitan)骨架復原模型

圖19 華北龍 (Huabeisaurus)骨架復原模型
2.2 關于大型蜥腳類恐龍如何進食和消化巨量的食物
梁龍由于化石材料積累較多,對它們的了解也較多。如其牙齒只長在嘴前部,且很細小,牙冠高呈葉狀,橫斷面呈橢圓形。顯然只能吃柔嫩多汁的植物,沒有咀嚼用的牙齒。美國密歇根大學 Paul Upchurch等于 2000年從其牙齒磨損方式看,磨損面在牙齒一側,而不象其他類型恐龍那樣磨損面在牙齒頂端,所以認為是從植物上剝扯下嫩枝葉直接吞咽,沒有咀嚼。中生代繁茂的松柏類和蕨類植物中,哪些才是梁龍的食物?通過電腦模擬發現,松柏類的枝葉,容易卡在其牙縫里;而蕨類植物卻能很順利地剝食,所以推測其主要食物是后者。因在化石中發現可疑似胃石的卵石,所以推測其可能通過胃的蠕動,由胃石研磨稀爛 (也許還通過發酵)后消化。由于這個消化過程較長,估計要好幾天,所以包括梁龍在內的大型蜥腳類恐龍都具有巨大的胃,以存放數以噸計的待消化食物,故而身軀非常粗碩。
德國波恩大學 P Martin Sander等于 2010年的研究發現,動物體形越大,食物需求量就越大。大型蜥腳類恐龍以植物為食,植物是低熱量食物,所以素食恐龍的進食量比肉食恐龍的要大許多倍,每天要花費大量時間進食。非洲象 (Loxodonta africana)是現代最大的陸地動物,每天要花費 18h進食,幾乎沒有時間睡覺。按大型蜥腳類恐龍遠比現代非洲象龐大得多的體形推算,如采用相同的方式進食,即使全天不停地吃,也無法滿足其自身能量的需求。例如,迷惑龍 (Apatosaurus)1天需 30h不停進食才行,這顯然是不可能的。所以,大型蜥腳類恐龍必須有進食效率更高的生理結構。大型蜥腳類的小頭、長頸生理結構具有大大提高進食效率的重要作用。因進食方式采取直接吞咽,就無需花費大量時間咀嚼,提高了單位時間內的進食量。同時,頭部也無需咀嚼用的臼齒和相關的大塊肌肉,實現了頭骨小巧化,從而為演化出超長的脖子創造了條件。長脖子能進一步提高進食效率,蜥腳類恐龍通常體重幾十噸,行動起來并不輕松,靈活的長脖子可使這些龐然大物站在原地環顧四周尋找食物,快速進食。化石證據顯示,蜥腳類恐龍的頸椎多氣囊,瓣膜系統也很發育,既有助于呼吸,也減輕了長脖子的重量,方便了身體的平衡。另外,中生代大量生長的木賊 (Equisetum)植物,營養豐富但卻含有對牙齒有害的硅酸鹽,現生動物很少有以它們為食的。蜥腳類恐龍只吞不嚼的進食方式,可大大減少硅酸鹽對牙齒的損害,從而獲得比其他咀嚼進食動物更大的進化優勢。
2.3 關于大型蜥腳類恐龍體貌特征和生長速率
1992年,美國學者 S A C研究美國懷俄明州發現的梁龍化石標本,發現其尾的背面保留高高突起的三角形棘刺,由此推測梁龍沿脊椎骨直立排列著棘刺。在現生的一些爬行動物,如鬣蜥 (Iguana)和楔齒蜥(Sphenodon),也有類似結構。根據現代爬行動物研究,雄性個體的棘刺大于雌性個體的,在求偶時棘刺會豎起來,有炫耀和求偶的功能。另外,還可能有一定的防御和威脅敵害的功能。
梁龍的足跡化石證明,它們是成群活動,步態緩慢。足跡化石中沒有見到尾巴拖曳的痕跡,說明其長尾有強有力的肌肉和韌帶支撐,行走時是懸在空中的。其強有力的尾不但是鞭擊敵害的自衛武器,而且也是后腿站立的支撐。站立起來可吃到其他類型素食恐龍可望而不可及的高處植物,或可揮舞前肢反擊敵害的攻擊,其前腳內側趾上的大爪,也是重要的自衛武器。
德國波恩大學 P Martin Sander等于 2005年研究梁龍化石骨骼橫切面上的生長紋發現,其生長速率很快,從一個足球大小的蛋里胚胎,長成幾十米長的成年恐龍,僅需 10年。
2.4 關于大型蜥腳類恐龍的脖子是豎起還是前伸
美國俄勒岡大學 KentA Stevens等于 2005年測繪化石骨骼建立電腦模型進行模擬發現,梁龍科恐龍頸椎骨的嚙合方式使其不可能把頭抬得高出水平位置太多,所以其通常的體態是平伸脖子,橫掃地面,吃地上的植物。如要吃高處的植物,就要抬起前腿,用后腿站立才行。但英國樸次茅斯大學Michael P Taylor等于 2009年的研究認為,所有的四足動物都可豎起脖子舉首四顧,警戒或覓食。德國弗倫斯堡大學Andreas Christian 2010年通過對一中型蜥腳類恐龍師氏盤足龍 (Euhelopus.Zdanskyi,體長 10m~15m)的椎間軟骨進行應力測量分析,計算出其行走 100m所耗能量,結果顯示其能使脖子保持垂直向上的姿態達半小時之久——這足以讓它在一棵高樹前美餐一頓,因此認為至少一部分蜥腳類恐龍可較長時間豎起脖子。
2.5 關于大型蜥腳類恐龍的心臟
美國哥倫比亞大學 David Lambert于 1993年對梁龍的頸部結構研究后發現,如果按已知長度最長的頸子計算,需要 1個 1.6t的巨大心臟,才能把血液泵送到頭部,這樣心臟的重量幾乎達到其體重的1/10,顯然這是不可能的。由此提出,梁龍的頸部可能還有多個次級的輔助“心臟”,每個“次級心臟”僅有的用途就是把血液泵送到下一個“次級心臟”,如此接力傳送。
2.6 關于超龍和極龍
超龍(Supersaurus)的零星骨骼化石自 1972年被發現后,最初認為其親緣關系與腕龍(Barosaurus)接近。但懷俄明大學 D M Lovelace等于 2007年對后來找到的較完整骨架化石研究顯示,更接近迷惑龍(Apatosaurus)。超龍的標本最初由美國楊百翰大學 James A Jensen描述并命名為Super-saurus vivianae。同年,他還報道了另一些 1976年發現于猶他州的大型恐龍骨骼化石,包括 1個肩胛骨、一些脊椎骨和臀部骨骼殘片,認為這是已知最大的恐龍,并命名為“極龍”(U ltrasaurus,意思是“極大的恐龍”),在新聞媒體里報道。但當他 1985年在學術刊物里發表其描述,并正式命名為“馬氏極龍”(U ltrasaurusm acintoshi)時,這個屬名已在 1983年被韓國釜山大學的金港墨 (Haang Mook Kim)搶先用于命名韓國發現的一大型蜥腳類恐龍——極龍 (U ltrasaurus tabriensis)。金港墨認為韓國發現的這種恐龍,與 J A Jensen在美國猶他州發現的“極龍”親緣關系相近,但他的描述卻將股骨和肱骨混淆了,而且因骨骼過于殘缺,“極龍”這個屬并未被普遍認可。
由于金港墨的搶先發表,J A Jensen只好把自己的“馬氏極龍”改名為“馬氏極超龍”(U ltrasauros m acintoshi)。但進一步研究發現:“馬氏極超龍”的骨骼根本不屬于一個恐龍,肩胛骨實際上屬于一巨大的腕龍(B rachiosaurus);而脊椎骨則屬于另一巨大的超龍(Supersaurus),于是極超龍這個命名被廢棄。
3 結 語
大型蜥腳類恐龍曾是陸地上最大的動物,為取得生存競爭優勢,對抗肉食恐龍的攻擊,并與其他素食恐龍爭奪食物資源,向大型化演化有利于其占據更大的生態位空間。大型蜥腳類恐龍的一切特征都是演化的結果,其最主要的演化是體形的大型化,其他的演化都是圍繞這個主線的配套,如:①小頭長頸,是為了不用過多移動龐大的身軀就能在大范圍內覓食。②大鼻孔,是為了大吞吐量的呼吸,供應龐大身軀的生命活動之需,由此可推想其心肺功能的強大,甚至可能除了一個主心臟外,還可能有多個小型輔助“心臟”,以免單個心臟負擔過重。③長形的葉狀牙齒,是為了快速連續地進食,力求每次都能撕扯下一大片鮮嫩的植物枝葉,不經咀嚼直接吞咽,實現在單位時間內盡量多吃,以滿足龐大身軀生命活動所需的能量供應。④股骨比脛骨長,說明其雖不善于奔跑,但后肢支撐力很強,有時可站立起來吃高處的植物,或用前肢撲打敵害進行自衛。也說明其不是靠快速奔跑擺脫敵害,而是依仗龐大身軀及強大力量對抗敵害。⑤四足行走,是為了把龐大身體的重量分散到 4個寬大的腳掌上,減小對地面的壓強,提高通過松軟地面的能力。
也就是說,大型蜥腳類恐龍體形增大的同時,其全身的組織器官、身體結構和生理特點也相應地進行了一系列為適應大型化的配套演化。如,巨大的進食量,配套的是獨特的進食方式,牙齒呈高冠細長葉狀,適合大片地撕扯、剝離植物枝葉,直接吞咽。省略咀嚼這一步驟,又加快了進食速度,單位時間的進食量大大增加。頭骨因無需生長咀嚼肌肉,可以向輕小化發展。頭骨輕小,又可以向長頸化發展,演化出比身軀還要長的靈活頸部,從而不必經常移動龐大的身軀,就能上下左右大范圍地快速覓食。龐大的身軀可以容納巨大的胃,存放因未經咀嚼而消化緩慢的食物。也許還要依靠吞下一些卵石,通過胃部肌肉的蠕動來磨爛食物,幫助消化。太長的頸部可能還需要有多個輔助“心臟”來接力泵送血液。
由此可見,生物演化對其自身的生理和身體結構也是牽一發而動全身的,需要相互配套演化以求盡量達到對環境的最佳適應。
[1] G ILLETTE D D.Seism osaurus halli,gen.et sp.nov.,a new sauropod dinosaur from theMorrison For mation(Upper Jurassic/Lower Cretaceous)of New Mexico,USA[J].JVP 11,1991(4):417-433.
[2] LOVELACE D M,HART MAN S A,WAHL W R.Morphology of a spec imen ofSupersaurus(Dinosauria,Sauropoda)from theMorrison Formation ofWyoming,and a re-evaluation of diplodocid phylogeny[J].Arquivos do Museu Nacional,2007,65(4):527-544.
[3] G ILLETTE D D.Sam’s stomach stones.Seism osaurus-The earth shaker[M].Columbia:Columbia Univ.Press.NY.1994:95-114;195-199.
[4] CARPENTER K.Biggest of the big:a critical re-evaluation of the mega-sauropodAm phicoelias fragillim us[C]∥FOSTER J R,LUCAS S G.Paleontology and Geology of the Upper JurassicMorrison For mation.NewMexicoMuseum of Natural History and Science Bulletin 36,2006,131-138.
[5] STEVENS KA,PARR ISH JM.NECK POSTURE.Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs[C]∥CARPENTER KENNETH,T IDS WELL V IRG IN IA. Thunder Lizards:The Sauropodomorph Dino-saurs.Bloomington:Indiana University Press,2005:212-232.
[6] YADAGIR I P,AYYASAM I K.A carnosaurian dinosaur from the Kallamedu Formation(Maestrichtian horizon),Tamilnadu[C]∥M V A SASTRY,V V SASTRY,C G K RAMANUJAM,et al.Symposium on Three Decades of Development in Palaeontology and Stratigraphy in India.Volume 1.Precambrian to Mesozoic.Geological Society of India Special Publication,1989,11(1):523-528.
[7] KRAUSE,D W,O'CONNOR,P M,CURRY ROGERS K,et al.Late Cretaceous terrestrial vertebrates from Madagascar: Implications for Latin American biogeography[J].Annals of the Missouri Botanical Garden,2006,93(2):178-208.
[8] NOVAS F E,SALGADO L,CALVO J,et al.Giant titanosaur(Dinosauria,Sauropoda)from the Late Cretaceous of Patagonia[J].Revisto delMuseo Argentino de CienciasNaturales,n.s.2005,7(1):37-41.
[9] JENSEN J A.Three new sauropod dinosaurs from theUpper Jurassic of Colorado[J].Great Basin Naturalist,1985,45:697-709.
[10]BONAPARTE J,COR IA R.Un nuevo y gigantesco sauropodo titanosaurio de la Formacion Rio Limay(Albiano-Cenomaniano)de la Provincia del Neuquen,Argentina[J].Ameghiniana,1993,30(3):271-282. (in Spanish)
[11]DAV ID BURN IE.The Kingfisher IllustratedDinosaur Encyclopedia[M].Kingfisher Publications Plc.,2002.
[12]YOUNG C C,ZHAO X J.M am enchisaurus hochuanensis sp.nov[J].Institute ofVertebrate Paleontology and PaleoanthropologyMonographs,A,8,1972:1-30.
[13]WEDEL M J,R ICHARD L.CIFELL IR L,et al.Osteology,paleobiology,and relationshipsof the sauropod dinosaurSauroposeidon[J].Acta Palaeontologica Polonica,2000,45:343-388.
[14]WEDEL M J,CIFELL IR L.Sauroposeidon:Oklahoma’s native giant[J].Oklahoma Geology Notes,2005,65(2):40-57.
[15]CALVO J O,PORFIR I J D,GONZáLEZ-R IGA B J,et al.A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur[J].AnaisAcademiaBrasileira Ciencia,2007,79(3):529-41.
[16]S M ITH J B,LAMANNA MC,LACOVARA K J,et al.A Giant sauropod dinosaur from an Upper Cretaceous mangrove depositin Egypt[J]. Science,2001, 292(5522):1704-1706.
[17]MARSH O C.Description of new dinosaurian reptiles[J].American Journal of Science,1890,3(39):81-86.
[18]RUSSELL D,BELAND P,MC INTOSH J.Paleoecology of the dinosaurs of Tendaguru(Tanzania)[J].M oires de la SociétéGéologique de France,Nouvelle S rie,1980,59:169-175.
[19]WEDEL M J,CIFELL IR L,R ICHARD L,et al.Sauroposeidon proteles,a new sauropod from the Early Cretaceous ofOklahoma[J].JournalofVertebrate Paleontology,2000,29(1):109-114.
[20]YOU H L,L ID Q,ZHOU L Q,et al.Daxiatitan binglingi:a giant sauropod dinosaur from the Early Cretaceous of China[J].Gansu Geology,2008,17(4):1-10.
[21]TAYLOR M P.A Re-evaluation ofB rachiosaurus altithoraxRiggs 1903(Dinosauria,Sauropod)and its generic separation from Giraffatitanbrancai(Janensh 1914)[J].Journal ofVertebrate Paleontology,2009,29(3):787-806.
[22]LOVELACE D M,HART MAN S A,WAHL W R.Morphology of a spec imen ofSupersaurus(Dinosauria,Sauropoda)from theMorrison Formation ofWyoming,and a re-evaluation of diplodocid phylogeny[J].Arquivos do Museu Nacional,2007,65(4):527-544.
[23]FOSTER J,JURASSIC WEST.The Dinosaurs of the Morrison Formation and TheirWorld[M].Bloomington:Indiana University Press,2007.
[24]TANG F,KANG XM,J IN X S,et al.A new sauropod dinosaur of Cretaceous from Jiangshan,Zhejiang Province[J].Vertebrate Palasiatica,2001,39(4):272-281.
[25]YOU H L,L ID Q,ZHOU L Q,et al.Huanghetitan liujiaxiaensis.aNew SauropodDinosaur from theLowerCretaceous Hekou Group ofLanzhou Basin,Gansu Province,China[J]. Geological Review,2006,52(5):668-674.
[26]PANGQ Q,CHENG ZW.A new Family of sauropod dinosaur from the Upper Cretaceous of Tianzhen,Shanxi Province,China[J].Acta Geologica Sinica,2000,74(2):117-125.
[27]MO J Y,HUANG C L,ZHAO Z R,et al.A new titanosaur(Dinosauria:Sauropoda)from the Late Cretaceous of Guangxi,China[J].Vertebrata Palasiatia,2008,46(2):147-156.
[28]LUCAS S G,SPIELMAN J A,R INEHART L A,et al.Taxonomic status ofSeism osaurus hallorum,a Late Jurassic sauropod dinosaur from NewMexico[C]∥FOSTER J R,LUCAS S G.Paleontology and Geology of the Upper Morrison For mation.NewMexicoMuseum ofNaturalHistory and Science(bulletin 36),2006:149-161.
[29]UPCHURCH P,BARRETT PM.The evolution of sauropod feedingmechanism[C]∥Evolution of Herbivory in TerrestrialVertebrates.Cambridge Press,2000,Chapter 4:79-122.
[30]SANDER PM,CHR IST IAN A,CLAUSSM,et al.Biology of the sauropod dinosaurs:the evolution of gigantis m[J].Biological Reviews.(2010-04-29).http://onlinelibrary.wiley.com/doi/10.1111/j.1469.
[31]CZERKAS SA.Discovery of dermal spines reveals a new look for sauropod dinosaurs[J]. Geology,1992,20:1068-1070.
[32]SANDER PM,KLE IN N.Developmentalplasticity in the life history of a prosauropod dinosaur[J]. Science,2005,310:1800-1802.
[33]TAYLOR M P,WEDEL M J,NA ISH D.Head and neck posture in sauropod dinosaurs inferred from extant animals[J].Acta Palaeontologica Polonica,2009,54(2):213-220.
[34]CHR IST IAN A.Some sauropods raised their necks—evidence for high browsing in Euhelopus zdanskyi[J].BiologyLetters,2010,10:37-48.
[35]LAMBERT D.The Ultimate Dinosaur Book[M].London:The Natural HistoryMuseum,1993.
[36]LOVELACE D M,HART MAN S A,WAHL W R.Morphology of a spec imen ofSupersaurus(Dinosauria,Sauropoda)from theMorrison Formation ofWyoming,and a re-evaluation of diplodocid phylogeny[J].Arquivos do Museu Nacional,2007,65(4):527-544.
[37]K IM H M.Cretaceous dinosaurs from South Korea[J].Journal of the Geological Society of Korea,1983,19(3):115-126.
[38]UPCHURCH P,BARRETT P M,DODSON P.Sauropoda.D B WEISHAMPEL,H OS MóLSKA P DODSON.2nd edition.The Dinosauria[C]∥Berkeley:University of California Press.2004:259-322.
[39]CURTICE B,STADT MAN K,CURTI CE L.A re-assessment ofU ltrasauros m acintoshi(Jensen,1985)[C]∥M MORALES.The ContinentalJurassic:Transactionsof the Continental Jurassic Symposium,Museum ofNorthernArizona Bulletin number 60.1996:87-95.
[40]錢邁平,姜楊,蔣嚴根,等.華東白堊紀暴龍類恐龍蛋化石的新發現[J].地質學刊 (原《江蘇地質》),2008,32(2):86-97.
[41]錢邁平,應軍,姜楊,等.淅江白堊紀恐龍化石 [J].地質學刊,2009,33(4):337-345.
[42]錢邁平,章其華,姜楊,等.華東白堊紀鴨嘴龍類恐龍蛋化石的發現[J].地質學刊,2009。33(1):8-27.
Study on gigantic sauropoda dinosaurs
QI ANMa i-ping1,HU Ba i-xiang2,ZHAN Geng-shen2,ZOU Song-mei2,ZHANG Qi-hua2
(1.Nanjing Institute of Geology andMineral Resources,Nanjing 210016,China;2.Geological Survey of Jiangsu Province,Nanjing 210018,China)
Gigantic herbivorous sauropoda dinosaurswere very huge creatures.D iplodocus,among them,was up to 54 meters long and was the biggest terrestrial animal even.Allof the dinosaurs,whose lengthwasover 20 meters,were attached to sauropoda.Theirmain evolution was gigantis m for having advantage of getting ecological niche,and other evolutionswere cooperated with it.The characteristics of sauropoda dinosaurs,for example:① Their long neck with a tiny head would have had the ability to browse atmany levels and orientation needed not to move their gigantic body often.②Their large nareswould have allowed to breathe air in large throughput to support their gigantic body living,and so a sauropoda dinosaur would have had strong blood circulatory system,perhaps had a main heart and rudimentary auxiliary hearts'for fear a single heart too huge.③Their spatulate-shaped teeth would have had the ability to stripped longer portions of plant's stems in a single action,and ingested apace without mastication.④Their femurs were longer than their tibias,and showed theywould have been ineffective runners but their thick legs were powerful,especially hind legs,and could stand up to graze on high plants,or to swat enemies.⑤Theywere quadrupedswith verywide and large paws,so a sauropoda dinosaur would have shared the weight of their titianic bodies by the four legs,and reduced pressure to ground for fear sinking.Thus it can be seen that:in live evolution,what happens to a s mall partmay affect the whole.An organis m'smain evolution would have driven a series of cooperated evolutions in itself for adapt to the environment suitably asmore as possible.
Sauropodomorpha dinosaurs;Gigantis m;Mesozoic;Period
Q915.2+3
A
1674-3636(2010)04-0337-14
10.3969/j.issn.1674-3636.2010.04.337
2010-09-08;編輯:詹庚申
國家自然科學基金(40872006)
錢邁平 (1954—),男,研究員,博士后,地層古生物專業.
