鞍帶石斑魚工廠化育苗研究
2010-10-23 03:02:24黃宗文郭仁湘陳國華
海洋科學 2010年9期
黃宗文, 駱 劍, 林 彬, 郭仁湘, 楊 薇, 陳國華
(海南大學 海洋學院 熱帶生物資源教育部重點實驗室, 海南 海口 570228)
鞍帶石斑魚工廠化育苗研究
黃宗文, 駱 劍, 林 彬, 郭仁湘, 楊 薇, 陳國華
(海南大學 海洋學院 熱帶生物資源教育部重點實驗室, 海南 海口 570228)
作者設計了一種新的鞍帶石斑魚(Epinephelus lanceolatus)工廠化人工育苗方法。測定了鞍帶石斑魚人工育苗過程中水環境因子和育苗水體浮游生物的種類、數量變化。育苗期間水溫28.0~30.7; ℃鹽度為27.0~32.0; pH值為7.39~7.90; 溶氧量6 mg/L以上; 中午池面平均光照3315.9 lx; 記錄了1 d的光照、水溫、溶解氧和pH的變化; 育苗池前18 d只加水不換水, 氨氮不斷上升, 最低值0.24 mg/L, 最高值 2.53 mg/L, 育苗后期采取換水措施, 氨氮迅速下降; 育苗期間, 水體除正常投喂的輪蟲(Brachionus plicatilis)和橈足類外, 池中出現浮游植物 12種、浮游動物 5種, 浮游植物平均生物量6.92×106個/L, 原生動物平均生物量0.74×106個/L。經過34 d的培養, 得到平均全長2.45 cm魚種, 單位水體育苗密度300尾/m3, 育苗成活率為2.73%。結果表明, 用有效微生物群(effective microorganisms,簡稱EM) +高級蝦片調節水質的模式, 可以保持育苗水質穩定, 成功培育出鞍帶石斑魚幼魚。
鞍帶石斑魚(Epinephelus lanceolatus); 人工育苗; 水環境因子
鞍帶石斑魚(Epinephelus lanceolatus)俗稱龍膽石斑魚, 隸屬鱸形目(Perciformes)、科(Serranidae)、石斑魚屬( Epinephalus), 為暖水性、中下層珊瑚礁魚類, 分布于印度洋非洲東岸至太平洋中部密克羅尼西亞, 南至澳大利亞, 中國產于南海諸島和海南島等海域, 但數量稀少。鞍帶石斑魚是石斑魚類中體型最大的種類[1], 具有生長速度快, 抗病性強, 肉質鮮美等諸多優點, 且肌肉氨基酸組成與人體氨基酸組成較為接近, 營養價值高, 成為養殖前景極大的石斑魚種類, 深受消費者[2]和養殖業者的喜愛, 由于人工繁殖技術尚不成熟, 苗種不足限制了鞍帶石斑魚養殖的發展。
石斑魚魚苗培育被認為是海水魚類育苗中難度較大的一種[2], 而鞍帶石斑魚又有親魚獲取較難、產卵期短等問題, 使人工繁殖更加困難。目前, 學者們[2~7]對于石斑魚育苗存在的問題達成了幾點共識: 首先,育苗水環境因子的變化是育苗成功與否的重要因素之一[3,4]; 第二, 由內源性營養轉為外源性營養的仔魚開口期餌料不適宜[3], 長棘期和收棘期的營養缺乏, 均可導致高的死亡率; 此外, 由于魚苗密度較高產生的局部缺氧和稚、幼魚期的互相殘殺也是育苗成活率低的重要原因。目前, 鞍帶石斑魚育苗多采用池塘生態育苗方式[6,8,9], 鞍帶石斑魚在海南的主要產卵期是 4~6月和 9~10月, 前一個產卵期內, 海南的池塘育苗中水溫經常達到 32℃以上, 而這個水溫條件在鞍帶石斑魚的育苗早期仔魚很難渡過; 后一個產卵期是海南的臺風季節, 臺風的影響經常導致池塘育苗失敗。可見, 鞍帶石斑魚的池塘育苗受天氣制約, 工廠化育苗可以在一定程度上克服受天氣制約的問題。鞍帶石斑魚的工廠化育苗相關報道較少[2,5],作者曾采用單細胞藻類凈化水質的方法[3]進行鞍帶石斑魚的育苗, 但效果不穩定。為了改進鞍帶石斑魚的工廠化育苗方法, 本實驗是通過向育苗池中定時定量加入 EM 菌和高級蝦片, 在育苗水體中構建一條碎屑食物鏈, 保持育苗水體水質穩定, 完成育苗過程, 現將實驗總結如下。
1 材料與方法
1.1 育苗設施與材料
育苗實驗在萬寧業興水產養殖有限公司石斑魚育苗基地(海南省萬寧市和樂鎮港北港)進行。
水源: 海邊砂濾井過濾海水。
育苗池條件: 室內水泥池, 方形, 規格為5 m×5 m×1 m; 設有上溢下排的排水口和池底排污口; 全池設25只氣石充氣; 育苗車間的房頂部分采用透光材料, 設有遮光網, 可調節室內光照強度, 控制11: 00~15: 00 育苗池水面光照強度 3 000~5 000 lx。
EM菌: 市售東方菌王10 L, 清水20 L, 紅砂糖2 kg, 在50 L聚乙烯桶中混勻, 密封備用。
蝦片: 市售金蝦牌高級蝦片。
生物餌料: 用池塘培育的輪蟲、橈足類。
鞍帶石斑魚受精卵0.2 kg。
1.2 育苗方法與水質管理
1.2.1 育苗前的準備與投放仔魚
培水: 育苗池在使用前用漂白粉消毒, 沖洗至無余氯。加入海水至80 cm深。育苗池進水當天開始, 每天8: 00、15: 00分2次向池中潑灑準備好的EM菌和蝦片。
投放仔魚: 培水至第4天, 購鞍帶石斑魚受精卵0.2 kg, 孵化出仔魚后小心投放到育苗池; 仔魚下池3 h后, 抽樣測定密度為1.1萬尾/m3。
1.2.2 餌料投喂和水質管理
1.2.2.1 餌料投喂
仔魚孵出第2天, 向育苗池中接種輪蟲, 保持育苗水體輪蟲密度 15~20個/mL。第 14天開始, 繼續投喂輪蟲外, 投喂少量經80目篩絹過濾的橈足類幼體。第 20天, 以投喂橈足類為主, 保持育苗水體橈足類密度為5~10個/mL。
1.2.2.2 水質管理
仔魚下池后: 第一周, 不換水; 第二周, 每天向池中加水1~2 cm; 第三周每天換水3~8 cm; 第四周以后, 根據育苗水質狀況, 每天換水20~30 cm。
1.2.3 育苗水體理化條件和浮游生物的測定
1.2.3.1 水溫、pH、鹽度、溶氧、光照強度
每天8: 00、15: 00兩次測定。水溫、溶氧測定使用美國HACH HQ30D flexi METER, 鹽度測定使用手持式折光鹽度計; pH測定用DELTA 320臺式pH計, 光照強度測定用DT-1301 LIGHT METER測定水面照度。
1.2.3.2 氨氮測定
HACH DREL 2800便攜式完全水質分析實驗室的方法。
1.2.3.3 浮游生物密度測定
育苗期間, 每隔3 d取樣測定1次輪蟲、橈足類以外的浮游生物密度。
1.2.3.4 浮游植物和浮游動物(原生動物)的定性測定
取水樣300 mL, 用150目篩絹過濾去輪蟲、橈足類等小型浮游生物, 用魯哥氏液固定(終濃度為1%), 顯微鏡下觀察、鑒定種類。
1.2.3.5 浮游生物的定量測定
取水樣300 mL, 用150目篩絹過濾去輪蟲、橈足類等小型浮游生物, 用魯哥氏液固定(終濃度為1%), 水樣靜置 24 h, 用虹吸法吸去上清部分, 定容到30 mL。取0.08 mL濃縮水樣到計數板, 顯微鏡下分別計數浮游植物和浮游動物(原生動物), 每個水樣觀察計數2個計數板。每個計數板計數50個視野,記錄每個視野浮游生物個數, 再計算單位原水樣中浮游生物的數量。
1.2.4 魚苗生長的觀察和密度測定
每日兩次觀察記錄魚苗的活動狀況; 每 3天取樣 1次, 每次10~20尾, 經終濃度 4%甲醛固定后測量全長并描述主要形態特征。
石斑魚的密度測定, 在夜間無光條件下, 待魚苗在池中分散均勻, 用500 mL燒杯隨機取樣3~5次,取平均值估算魚苗密度。
2 結果與分析
2.1 育苗期間理化因子和浮游生物的變化
2.1.1 育苗水體理化因子的變化及對育苗的影響
2.1.1.1 水溫和鹽度
育苗期間, 水體溫度范圍在 28.0~30.7℃、鹽度27.0~32.0, 魚苗生長發育正常。在育苗過程中, 遇一次臺風, 室外溫度驟降, 因育苗車間的保溫作用, 室內育苗水溫變化較小, 沒有對育苗造成大的影響。育苗早期不換水, 因蒸發鹽度有小幅上升趨勢, 至育苗的第三周, 因降水影響, 海邊砂濾井的海水鹽度有所下降, 此時對育苗池少量換水, 鹽度有小幅下降, 對魚苗生長沒有明顯影響(圖1)。
2.1.1.2 溶氧
育苗水體采取不間斷充氣, 水體溶氧在 6 mg/L以上, 溶氧條件良好(圖2)。
2.1.1.3 光照
育苗車間的光照變化很大, 主要受天氣變化的影響, 車間設置的日光燈, 對實際光照影響不大, 圖3可以看出, 育苗期間出現過3次低光照情況。
第一次在育苗的第四、五天, 持續時間1 d, 正值仔魚從內源營養向外源營養的過度期, 從育苗的實際效果看, 對魚苗生長沒有產生大的影響; 第二次遇到光照情況是在第15~17天(臺風影響), 持續時間2 d多, 從池中的魚苗密度變化看, 育苗受到一定的影響; 第三次是在育苗的第21~23天, 池中出現一些死苗現象。
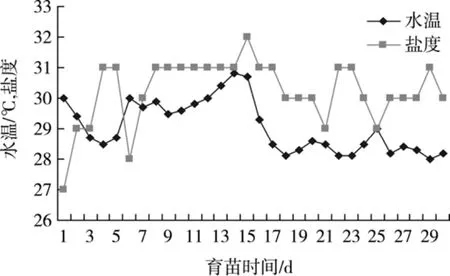
圖1 育苗水體中溫度和鹽度的變化Fig. 1 Variations of temperature and salinity in breeding water
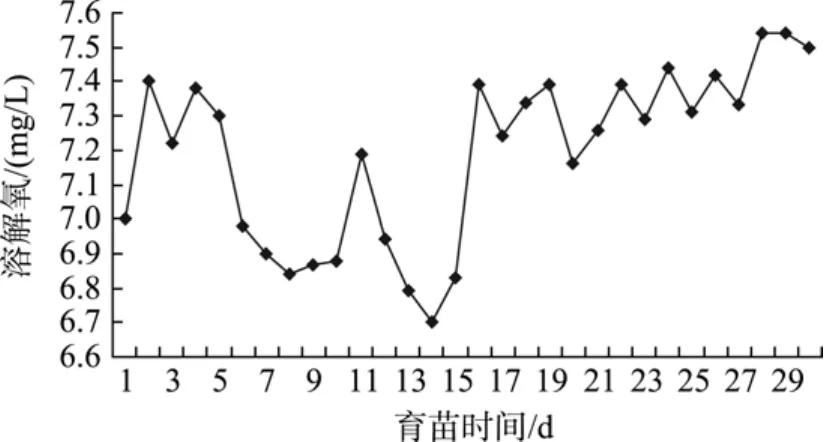
圖2 育苗水體中溶解氧的變化Fig. 2 Variation of DO in breeding water
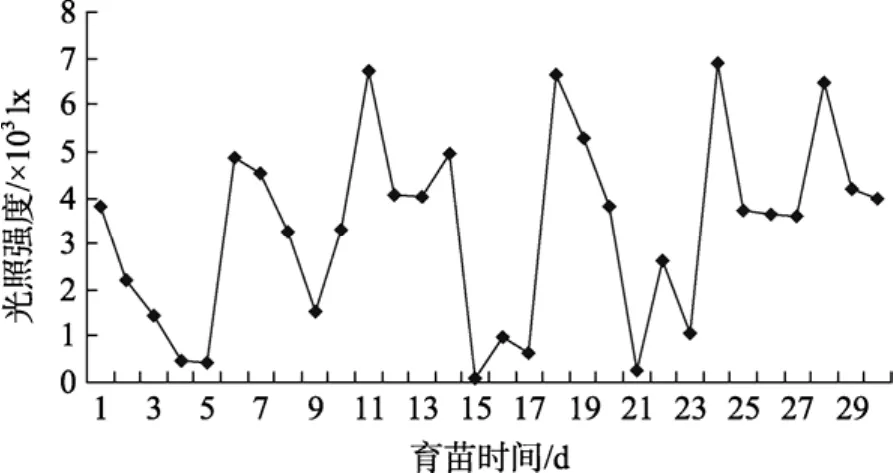
圖3 育苗光照強度的變化Fig. 3 Variation of illumination in whole period
2.1.1.4 酸堿度
從圖 4可以看出, 育苗水體酸堿度在 pH7.39~7.90的范圍變化, 隨著育苗進程, pH值有逐漸降低的趨勢。
2.1.1.5 環境因子的周日變化與魚苗活動規律
選定一個晴天, 即在育苗的第 13天, 每隔 2 h測定一次水溫、溶氧、光照、pH, 以了解其變化規律(表1)。從總體看, 僅光照取自然光, 變化明顯, 水溫、溶氧、pH都相對穩定。此時鞍帶石斑魚苗背鰭和腹鰭都已經長出長棘, 測量理化因子的同時, 觀察了魚苗的活動情況。6:00, 車間內光照為 109.0lx,魚苗胃內食物極少或者沒有, 開始從晚上的隨水漂浮轉為主動游泳, 之后, 出現身體向前竄動的攝食動作; 8: 00, 攝食動作明顯加強, 魚苗腹部飽滿, 以后整個白天, 都有攝食動作, 且腹部飽滿; 至20: 00,車間內光照強度為0, 用燈光照射育苗池發現魚苗隨充氣造成的水流翻滾, 無攝食動作, 偶爾因燈光刺激而活動。

圖4 育苗水體的pH值變化Fig. 4 Variation of pH in breeding water
2.1.2 水體氨氮含量變化及對魚苗生長的影響
育苗期間, 4次測定了水體的氨氮含量, 育苗第1天(水源)、第7天、第18天、第29天的氨氮含量,分別為0.24, 0.47, 2.53, 0.97 mg/L。從結果看, 本方法進行育苗, 水體氨氮含量不斷增高, 在第 18天達到最高(因為此前不換水), 以后隨著換水量增大, 氨氮含量下降。第18天的氨氮含量達2.53 mg/L, 魚苗活動并無明顯異常。
2.1.3 育苗水體浮游生物的測定
2.1.3.1 育苗水體浮游生物的種類組成
育苗第7天開始, 每隔3 d采樣1次, 測定育苗池中輪蟲、橈足類以外的浮游生物種類, 見表2。
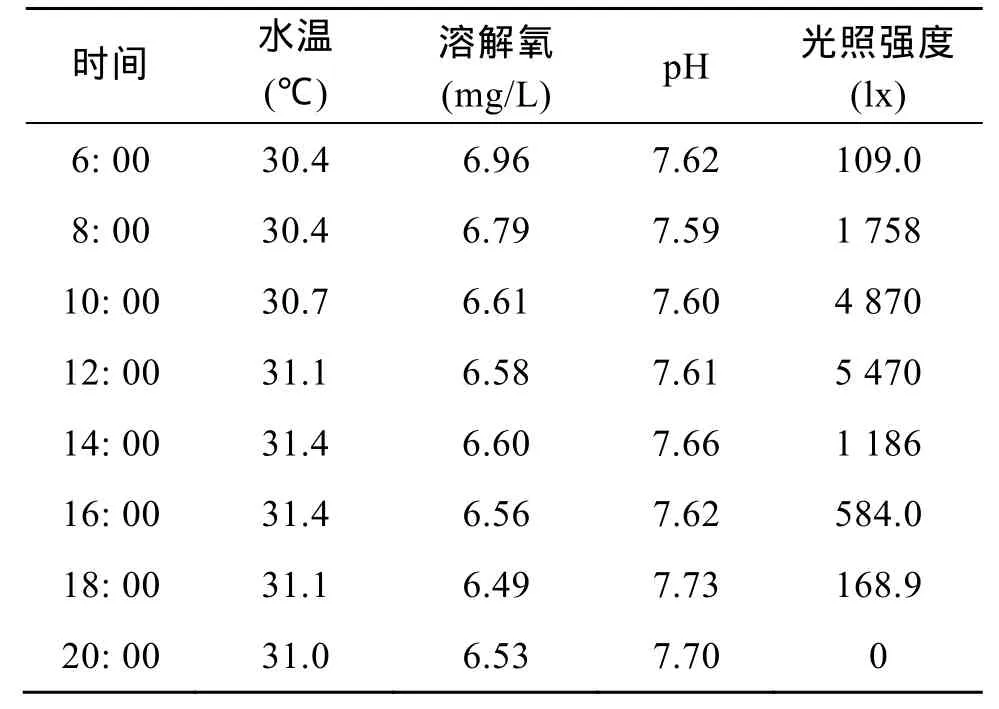
表1 24 h育苗水體環境因子的變化Tab. 1 Daily variations of environment factors
2.1.3.2 育苗水體中微型浮游生物量的變化
育苗池換水之前, 分 4次取樣測定了浮游植物和浮游動物(原生動物)的生物量。浮游植物生物量: 第 7天5.77×106個/L, 第11天4.36×106個/L, 第15天7.71×106個/L, 第19天9.85×106個/L; 浮游原生動物的生物量: 第 7天 1.78×106個/L, 第 11天 0.74×106個/L,第15天0.30×106個/L, 第19天0.15×106個/L。這些浮游生物(浮游植物和浮游原生動物)是育苗過程中自然出現在育苗池中, 在育苗池中生長, 構成了育苗池中生物的組成部分, 在育苗池中發揮作用。

表2 育苗水體中出現的浮游生物種類Tab. 2 Planktons in breeding pools
2.2 魚苗密度變化和育苗效果
育苗期間, 4次測定了池中的魚苗密度。第1天(仔魚下池)密度為 11×103尾/m3, 第 7 天為 10×103尾/m3, 第 19天為 2×103尾/m3, 第 30天為 0.3×103尾/m3。隨著育苗時間的推移, 魚苗密度逐漸下降,符合石斑魚育苗的一般規律。育苗到第30天, 育苗成活率為2.73%。
2.3 魚苗生長情況
2.3.1 初孵仔魚
剛孵化出膜的仔魚, 身體前端有一個橢圓形半透明的卵黃囊, 卵黃囊后部有一小油球, 初孵仔魚由于油球的作用傾斜浮在水面上, 偶爾垂直向上游動。
2.3.2 開口仔魚
放苗的第 3天, 觀察到卵黃囊明顯變小且接近消失, 此時期仔魚由開始由內源性營養型向外源性營養型轉變, 口部開始形成, 出現胸鰭原基, 在水中觀察可明顯看到仔魚頭部有兩個黑點, 腹部、尾部各一個黑點, 頭朝下浮于水中, 已可在水中平游。用燒杯觀察時, 可見仔魚用吻部用力的啄燒杯底部, 有明顯的攝食行為, 此時應保證水體中有足夠的適合仔魚攝食的餌料。
2.3.3 背棘和腹棘根芽出現
放苗的第7天, 此時仔魚全長為0.250 cm±0.02 cm, 背棘和腹棘根芽出現, 頭部增高, 消化管上部黑色素增大, 在池中有聚群行為, 喜歡在池角和邊緣活動, 有趨光行為。用燒杯打起觀察, 可明顯看到仔魚身體兩側長出一對翅狀鰭條, 振動較快, 黑色素加深, 仔魚攝食能力增強, 活力較好。
2.3.4 背鰭長棘和腹鰭長棘出現
放苗的第 10天, 此時全長 0.350 cm±0.01 cm,背鰭位置上出現一枚長棘, 棘上有黑色素分布, 棘緣呈鋸齒狀, 在身體兩側的腹鰭位置上也長出兩枚長棘, 與背棘相似, 魚體全身布滿黏液。用燒杯觀察,呈“小三叉”狀。魚苗在池中分布面積稍微擴大, 但四角的分布密度仍然比較高, 四壁稍低。主要在水中中上層活動。
2.3.5 背鰭、臀鰭鰭條出現
放苗的第17天, 此時全長0.8 cm±0.1 cm, 背鰭長棘和腹鰭長棘繼續加長, 呈“大三叉”狀。長棘長及尾部, 骨骼和肌肉系統迅速發育, 身體中軸銀色脊索明顯可見, 背鰭、臀鰭、尾鰭鰭條均可觀察到。消化道進一步分化。
2.3.6 背鰭、臀鰭、尾鰭鰭條形成
放苗第23天, 全長1.45 cm, 第二背鰭棘和腹鰭棘絕對長度達到仔稚魚階段的最大值。魚體頭部眼眶斜后上方色斑增多并有向胸鰭附近擴大之勢。第一背鰭和第二背鰭已連在一起, 臀鰭第一鰭條變粗形成鰭棘, 各鰭已基本形成。
放苗第34天, 平均全長2.45 cm, 部分生長快的個體開始長出鱗片。此時, 魚苗可以進入下一階段的標粗培育, 實驗結束。
3 討論
3.1 水環境因子對育苗的影響
育苗水體溫度是育苗成功與否的關鍵水質指標。很多學者皆對育苗過程中水體適宜溫度進行論述與報道。周仁杰[10]在斜帶石斑魚(Epinephelus coioides)育苗試驗的結果顯示, 水溫范圍在 27.3~30.5℃, 魚苗生長發育正常; 王云新[11]認為斜帶石斑魚最佳培養水溫在 25~27℃; 陳國華[3]報道點帶石斑魚(Epinephelus malabaricus)仔魚期適宜水溫為 24~29℃, 低于或高于這個范圍則會影響仔魚的正常活動, 而稚魚期適溫略寬。本實驗中鞍帶石斑魚育苗期間測得水溫范圍在 28~30.8℃,魚苗的發育正常, 與黃進光[5]報道的鞍帶石斑魚育苗水體溫度為 27~29.5℃接近。龔孟忠[2]在水溫 23.30~27.0℃的條件下也取得了鞍帶石斑魚育苗的成功。
育苗水體鹽度、溶氧量、pH值的變化范圍與斜帶石斑魚[10,11]、點帶石斑魚[3,4]等基本一致。
3.1.1 光照對育苗的影響
本次育苗遇到 3次低光照情況, 持續時間都不長, 從育苗效果看, 沒有造成大的影響。正常情況下,鞍帶石斑魚的育苗車間取自然光, 育苗池水面光照4 000~5 000 lx是適宜的。
3.1.2 氨氮對育苗的影響
非離子氨對水生物有較強的毒性, 中國漁業水質標準規定非離子氨≤0.20 mg/L。周永欣等[12]研究未離解氨對草魚種的毒性, 未離解氨對26、47、125日齡草魚的96 h LC50分別是0.570、1.609和1.683 mg/L, 對 47、60、125日齡的 48 h LC50則分別為1.727、2.050和2.141 mg/L。在亞急性毒性試驗中, 根據氨對草魚生長和鰓組織的影響, 認為未離解氨的最大允許毒物質量濃度為0.054~0.099 mg/L。曲克明等[13]研究氨氮對大菱鲆(Scophthatmus maximus)的急性毒性效應, 過飽和溶氧條件下, 非離子氨對大菱鲆的 48 h LC50值和 96 LC50值分別為 2.4 mg/L 和1.73 mg/L; 正常溶氧條件下, 非離子氨對大菱鲆的48 h LC50值和96 LC50值分別為1.82 mg/L和1.14 mg/L。采用本實驗的方法進行鞍帶石斑魚的育苗,在育苗第18天, 氨氮含量達到2.53 mg/L, 經用“海水水質標準(GB 3097-1997)附錄B”的非離子氨換算方法, 得非離子氨為0.051 mg/L, 并且在育苗過程中立即采取了換水措施, 未見魚苗出現急性中毒現象,但是否存在組織損傷等影響, 值得進一步研究。
3.2 EM菌+高級蝦片的生態育苗模式
用本實驗方法進行鞍帶石斑魚人工育苗, 實際上在水體中建立了一條碎屑食物鏈: 定時定量加入蝦片和微生物制劑, 利用微生物分解蝦片、各類生物的排泄廢物和殘體; 這樣的育苗方法, 育苗池中還會自然出現一些微型浮游生物(如原生動物); 作為餌料投入的輪蟲可攝食微型浮游生物、單細胞藻類等; 育苗后期作為餌料投入的橈足類, 攝食微型浮游生物、輪蟲和藻類等; 作為培育對象的石斑魚苗則攝食輪蟲、橈足類。這種EM菌+高級蝦片的育苗方法解決了石斑魚育苗中幾個不易解決的問題: (1) 提供了石斑魚的開口餌料。石斑魚育苗的難點之一在于如何解決幼苗開口期的營養需求[14]。龔孟忠[2]用雙殼類的受精卵、擔輪幼蟲和面盤幼蟲作為鞍帶石斑魚的開口餌料能夠順利渡過此時期, 但投喂受精卵對水質影響較大, 不宜多過投喂。本實驗利用水體中出現的微型浮游生物作為仔魚的開口餌料, 開口效果良好; (2) 石斑魚育苗前二周, 因仔魚太小, 對育苗池換水相當困難。目前, 常用的石斑魚人工育苗的方法, 均以接種單胞藻類來改善水質, 如利用小球藻(Chlorella)、金藻等[3~5]配成“綠水”培育仔魚, 使水體中維持一種生態平衡。本實驗的育苗方法, 實際上是在育苗水體構建了一條碎屑食物鏈, 物質和能量沿著食物鏈傳遞, 微生物和藻類利用水體中溶解的無機鹽, 避免無機鹽在水體中快速積累, 從而保持水質相對穩定, 達到育苗前二周不換水的目的。但需要說明的是, 這樣建立的育苗系統并不是一個能夠自我維持的體系, 只是在一定程度上保持相對穩定, 一些指標還是在不斷上升的, 如 NH3-N, NO2-N, COD等指標實際上也是在不斷上升的, 只是上升的速度有所控制, 并且能夠在 2周左右的時間內滿足石斑魚生長發育的需要, 正好解決石斑魚育苗早期(前2周)難于換水的問題。之后, 隨著魚苗的生長,活動能力增強, 可以通過對育苗池換水, 解決育苗池水中無機鹽不斷上升的問題, 使石斑魚育苗過程得以完成; (3) 輪蟲、橈足類作為石斑魚的餌料投入到育苗水體, 當天不可能全部被魚苗攝食, 總會存留一部分, 本實驗的育苗方法中, 育苗池中存在豐富的微生物、微型浮游生物, 成為了輪蟲、橈足類的餌料, 保證后者不會因饑餓降低其作為石斑魚餌料的價值, 有利于提高育苗成活率。
3.3 微型浮游生物進入育苗池的可能途徑和意義
石斑魚工廠化育苗中, 育苗池會自然出現一些微型浮游生物。分析傳入途徑可能有二: (1) 石斑魚育苗場都建造在海邊, 附近常有大量的養魚、養蝦池分布, 魚蝦養殖池的微型浮游生物可能通過空氣傳播, 養殖池大量使用增氧機, 更增大了微型浮游生物通過空氣傳播的可能性; (2) 育苗過程中必須向育苗池中投入輪蟲、橈足類, 在這些生物餌料中混雜有部分微型浮游生物。盡管這些微型浮游生物不是人為加入, 但按目前的工廠化育苗條件, 育苗池中出現一些微型浮游生物也是不可避免的。這些微型浮游生物在育苗池中生長, 成為育苗水體生物群落的組成部分, 起到保持水質穩定的作用。微型浮游動物在育苗池中的另一個重要作用是作為鞍帶石斑魚仔魚的開口餌料, 尤穎哲[15]曾投喂培育的原生動物作為鞍帶石斑魚仔魚的開口餌料, 取得了比投喂牡蠣受精卵或者S型輪蟲更好的培育效果。本實驗中, 育苗池中保持有較高的原生動物密度, 利用這類生物作為鞍石斑魚仔魚的開口餌料, 取得了較好效果。
[1] 張海發, 王云新, 劉付永忠, 等.鞍帶石斑魚人工繁育及胚胎發育研究[J].廣東海洋大學學報, 2008,28(4): 36-40.
[2] 龔孟忠, 陳慧, 范希軍.龍膽石斑魚引種及人工育苗技術的初步研究[J].福建水產, 2004, 1: 47-50.
[3] 陳國華, 張本.點帶石斑魚人工育苗技術[J].海洋科學, 2001, 25(1): 1-3.
[4] 全漢鋒, 劉振勇, 范希軍.點帶石斑魚人工育苗技術的初步研究[J].福建水產, 2004, 3(1): 31-34.
[5] 黃進光.龍膽石斑魚人工育苗技術初探[J].科學養魚,2006, 12: 34-36.
[6] 黎祖福, 陳省平, 莊余謀, 等.鞍帶石斑魚人工繁殖與魚苗培育技術研究[J].海洋水產研究, 2006, 27(3):78-85.
[7] 楊洪志, 梁榮峰.鞍帶石斑魚 Epinephelus lanceolatus(Bloch)繁殖生物學的初步研究[J].現代漁業信息,2002, 17(7): 20-21.
[8] 關則智.鞍帶石斑魚高位池育苗技術探討[J].中國水產, 2009, 2: 44-45.
[9] 蒲利云, 馮永勤, 董楊, 等.室外高位池鞍帶石斑魚人工育苗的初步試驗[J].科學養魚, 2009, 6: 26-27.[10] 周仁杰, 林濤.斜帶石斑魚人工育苗技術試驗[J].臺灣海峽, 2002, 21(1): 57-62.
[11] 王云新, 黃國光, 劉付永忠等.斜帶石斑魚人工育苗試驗[J].漁業現代化, 2003, 6: 14-15.
[12] 周永欣, 張甫英, 周仁珍.氨對草魚的急性和亞急性毒性[J].水生生物學報, 1986, 10(1): 32-38.
[13] 曲克明, 徐勇, 馬紹賽, 等.不同溶解氧條件下亞硝酸鹽和非離子氨對大菱鲆的急性毒性效應[J].海洋水產研究, 2007, 28(4): 83-88.
[14] 何永亮, 區又君, 李加兒, 等.石斑魚人工繁育技術研究進展[J].南方水產, 2008, 4(3): 75-79.
[15] 尤穎哲.龍膽石斑鱸人工育苗仔魚開口及前期餌料轉換技術研究[J].中國水產, 2007, 6: 54-55.
Industrial seed culture of Epinephelus lanceolatus
HUANG Zong-wen, LUO Jian, LIN Bin, GUO Ren-xiang, YANG Wei, CHEN Guo-hua
(College of Ocean, Hainan University, Key Laboratory of Tropic Biological Resources of Ministry of Education,Hainan University, Haikou 570228, China)
Mar., 31, 2010
Epinephelus lanceolatus; artificial breeding; water environment factors
We reported a modified industrial breeding method of Epinephelus lanceolatus. The variations of water environment factors, as well as the type and quantities of micro-planktons were recorded during the whole artificial breeding process. The fertilized eggs of E. lanceolatus could hatch completely and develop into normal larval fishes under the following conditions: 28~30.7℃, 27~32 salinity, pH 7.39~7.90, >6 mg/L DO, and 3315.9 lx average noon illumination intensity. Variations of these environmental factors during the time span of a day were also recorded every two hours. The concentration of Ammonia nitrogen went from 0.24 mg/L up to 2.53 mg/L 18 days after the addition of water, and it then dropped dramatically after water renewing at the later stage. During the whole culture process, 12 types of phytoplanktons and 5 types of zooplanktons were detected in the breeding pool. The average concentrations of plankton and protozoa were 6.92×106/L and 0.74×106/L, respectively. At the end of our 34-day industrial culture experiment, the average length of grouper larvae reached 2.45cm, and the density was 300/m3.The survival rate of grouper larvae was 2.73%. The result of this study indicated that using effective-microorganism(EM) mix shrimp-cracker method, the water quality could remain suitable for E. lanceolatus larvae growth.
S962
A
1000-3096(2010)09-0023-07
2010-03-31;
2010-06-18
國家科技支撐計劃資助項目(2007BAD29B01,2007BAD29B02); 海南大學重點科研資助項目(hd09xm11)
黃宗文(1984-), 男, 廣西北海人, 碩士生, 研究方向為熱帶水產養殖生物生殖調控與繁育, E-mail: wening201@163.com; 陳國華,通信作者, 教授, E-mail: chguh@hainu.edu.cn
(本文編輯: 譚雪靜)
