東方田鼠胚胎成纖維細(xì)胞永生化細(xì)胞系的建立
2010-09-09 09:20:32成鋼羅賽群蓋楠熊德慧李榮胡維新
成鋼,羅賽群,蓋楠,熊德慧,李榮,胡維新
(中南大學(xué)生物科學(xué)與技術(shù)學(xué)院分子生物學(xué)研究中心,長(zhǎng)沙 410078)
研究報(bào)告
東方田鼠胚胎成纖維細(xì)胞永生化細(xì)胞系的建立
成鋼,羅賽群,蓋楠,熊德慧,李榮,胡維新
(中南大學(xué)生物科學(xué)與技術(shù)學(xué)院分子生物學(xué)研究中心,長(zhǎng)沙 410078)
目的建立東方田鼠胚胎成纖維永生化細(xì)胞系,為全面研究東方田鼠抗日本血吸蟲機(jī)制以及開展不同動(dòng)物成纖維細(xì)胞間比較研究奠定基礎(chǔ)和提供細(xì)胞實(shí)驗(yàn)材料。方法運(yùn)用脂質(zhì)體介導(dǎo)的基因轉(zhuǎn)染法將pSV3 neo質(zhì)粒導(dǎo)入第3代東方田鼠胚胎成纖維細(xì)胞,經(jīng)G418篩選抗性克隆并擴(kuò)大培養(yǎng),建立永生化細(xì)胞系;用PCR檢測(cè)細(xì)胞株中SV40T基因的整合,RT-PCR鑒定SV40T基因在轉(zhuǎn)染細(xì)胞中的表達(dá);繪制東方田鼠胚胎成纖維永生化細(xì)胞生長(zhǎng)曲線。結(jié)果陽(yáng)性細(xì)胞克隆已擴(kuò)大培養(yǎng)并穩(wěn)定傳代50代,經(jīng)鑒定SV40 T抗原已整合到東方田鼠胚胎成纖維細(xì)胞中且穩(wěn)定表達(dá)。結(jié)論成功建立東方田鼠胚胎成纖維永生化細(xì)胞系。
東方田鼠;胚胎成纖維細(xì)胞;永生化;SV40T抗原
東方田鼠(M icrotus fortis,M f)是一種棲居于我國(guó)長(zhǎng)江流域的嚙齒類動(dòng)物,流行病學(xué)調(diào)查和人工感染實(shí)驗(yàn)均證明其對(duì)日本血吸蟲感染具有天然抗性,且這種抗性能穩(wěn)定遺傳[1]。我們前期實(shí)驗(yàn)已對(duì)M f胚胎成纖維細(xì)胞(Microtus fortis embryonic fibroblasts,M fEF)成功培養(yǎng)并進(jìn)行了相關(guān)生物學(xué)特性觀察。建立永生化M f胚胎成纖維細(xì)胞系,是M f實(shí)現(xiàn)實(shí)驗(yàn)動(dòng)物化,標(biāo)準(zhǔn)化,規(guī)范化的重要基礎(chǔ),也是研究M f發(fā)揮抗日本血吸蟲作用的重要試驗(yàn)材料。目前,已建立多種人和小鼠胚胎成纖維細(xì)胞系,M fEF的永生化細(xì)胞系尚為首次報(bào)道。近年來(lái),許多研究應(yīng)用病毒基因的轉(zhuǎn)染使細(xì)胞永生化,為細(xì)胞的長(zhǎng)期培養(yǎng)和功能研究提供了理想的方法和手段[2,3]。本研究采用脂質(zhì)體介導(dǎo)的基因轉(zhuǎn)染法,將質(zhì)粒pSV3 neo(含有SV40 T抗原基因和neo抗性基因)轉(zhuǎn)染第3代M fEF,旨在使其永生化,為M fEF長(zhǎng)期培養(yǎng)和全面研究東方田鼠抗日本血吸蟲機(jī)制以及開展不同動(dòng)物成纖維細(xì)胞間比較研究奠定基礎(chǔ)和提供細(xì)胞實(shí)驗(yàn)材料。
1 材料與方法
1.1 主要材料
20周齡,體質(zhì)量70~80 g的懷孕東方田鼠(長(zhǎng)江亞種人工封閉繁育第3代)由中南大學(xué)湘雅醫(yī)學(xué)院實(shí)驗(yàn)動(dòng)物學(xué)部提供。質(zhì)粒pSV3 neo系中南大學(xué)湘雅醫(yī)學(xué)院病理生理學(xué)教研室惠贈(zèng);Taq DNA聚合酶、Hind III、EcoR I、marker均購(gòu)自日本TaKaRa公司,質(zhì)粒抽提試劑盒購(gòu)自安比奧生物技術(shù)有限公司,新生小牛血清購(gòu)自杭州四季青生物工程有限公司; Lipofectam ineTM2000,購(gòu)自Invitrogen Life Technologies公司;DMEM培養(yǎng)基、胰蛋白酶、G418及Trizol購(gòu)自美國(guó)Gibco公司;其余常用試劑均為國(guó)產(chǎn)分析純。PCR引物由上海英俊生物有限公司合成。
1.2 方法
1.2.1 東方田鼠胚胎成纖維細(xì)胞的原代培養(yǎng):取妊娠12~13 d的孕鼠,乙醚麻醉后無(wú)菌取出子宮,用PBS漂洗3次,棄除表面血污,剪開子宮,取出帶有胎膜的胚胎,用鑷子撕破胎膜,取出胎鼠,剔除胎盤、胚頭、內(nèi)臟和四肢,無(wú)菌PBS漂洗3次,剔除可能附帶的脂肪組織、被膜結(jié)締組織、壞死組織以及紅細(xì)胞,將鼠胚軀干剪成約1 mm3的碎塊,放入離心管后用培養(yǎng)液反復(fù)吹打,靜置沉淀5 min,吸取上清于培養(yǎng)瓶?jī)?nèi),在含15%FBS、100 U/m L青霉素、100 U/ m L鏈霉素的DMEM,37℃,5%CO2條件下培養(yǎng)[4]。
1.2.2 質(zhì)粒pSV3 neo的擴(kuò)增與轉(zhuǎn)染:質(zhì)粒pSV3 neo提取、鑒定均按試劑盒操作說(shuō)明進(jìn)行。根據(jù)Invitrogenlife technologies公司提供的轉(zhuǎn)染操作說(shuō)明書進(jìn)行基因轉(zhuǎn)染:第3代M fEF細(xì)胞融合率達(dá)到70%~80%時(shí),用無(wú)血清無(wú)抗生素DMEM培養(yǎng)基分別將8~10 μg pSV3 neo質(zhì)粒DNA和15~20 μL脂質(zhì)體試劑稀釋至500 μL;將稀釋的DNA與脂質(zhì)體混合,室溫靜置20 min。M fEF用無(wú)血清DMEM培養(yǎng)基洗滌兩遍后,加入上述1 m L質(zhì)粒與脂質(zhì)體的混合物,于37℃,5%CO2的培養(yǎng)箱孵育6~12 h,即完成轉(zhuǎn)染過(guò)程。
1.2.3 轉(zhuǎn)染陽(yáng)性克隆細(xì)胞的篩選及擴(kuò)大培養(yǎng):轉(zhuǎn)染完成后更換普通培養(yǎng)基,將細(xì)胞接種于六孔板,24~48 h后更換含800 μg/m L G418的培養(yǎng)基開始篩選。陽(yáng)性細(xì)胞克隆生長(zhǎng)至一定數(shù)目后擴(kuò)增培養(yǎng)。細(xì)胞的換液、傳代、凍存及復(fù)蘇按常規(guī)方法進(jìn)行,復(fù)蘇后臺(tái)盼藍(lán)染色,計(jì)算細(xì)胞存活率。
1.2.4 SV40T基因在永生化細(xì)胞中的整合及表達(dá):①PCR擴(kuò)增:收集第5代和第20代細(xì)胞樣品,提取培養(yǎng)細(xì)胞的基因組DNA。根據(jù)SV40大T抗原基因序列設(shè)計(jì)引物,上游,5′-TGTGGTATGGCTGATT ATGA-3′;下游,5′-CGCAGTGAGTTTTTGTTAGA-3′,擴(kuò)增產(chǎn)物為391 bp。反應(yīng)條件:95℃預(yù)變性4 m in,然后95℃30 s,56℃30 s,72℃60 s,30個(gè)循環(huán),72℃延伸10 min。PCR產(chǎn)物用1%瓊脂糖凝膠電泳分析。②RT-PCR分析:按Trizol試劑盒操作法提取培養(yǎng)細(xì)胞的RNA,進(jìn)行逆轉(zhuǎn)錄。SV40大T抗原基因引物序列及大小同上。內(nèi)參引物為β-actin,上游引物:5′-GTGGGAATTCGTCAGAAGGACTCCTATG TG-3′,下游引物:5′-GAAGTCTAGAGCAACATAGC ACAGCTTCTC-3′,擴(kuò)增產(chǎn)物為260 bp,反應(yīng)條件同上,PCR產(chǎn)物以1%瓊脂糖凝膠電泳分析。
1.2.5 未轉(zhuǎn)染和轉(zhuǎn)染后M fEF生長(zhǎng)情況比較:取未轉(zhuǎn)染的第3代M fEF和轉(zhuǎn)染后第15代M fEF分別接種至48孔板,每孔接種10 000個(gè)細(xì)胞,每種濃度重復(fù)三孔,用血球計(jì)數(shù)板每天計(jì)數(shù)取平均值,連續(xù)7 d,繪制細(xì)胞生長(zhǎng)標(biāo)準(zhǔn)曲線。
2 結(jié)果
2.1 永生化M fEF的生物學(xué)性狀
倒置顯微鏡觀察:剛傳代接種的單個(gè)M fEF呈球形,懸浮于培養(yǎng)液中,胞質(zhì)透明,1~2 h后開始貼壁,細(xì)胞呈纖維狀生長(zhǎng),表面顆粒少,胞質(zhì)飽滿,輪廓清晰且細(xì)胞增殖較快,培養(yǎng)24~48 h可長(zhǎng)滿瓶底(圖1,彩插3);永生化M fEF與原代M fEF相比,可見較多的細(xì)胞分裂相和增殖細(xì)胞,在形態(tài)結(jié)構(gòu)上與原代細(xì)胞無(wú)明顯差異,細(xì)胞傳代后貼壁速度較原代細(xì)胞稍慢。
2.2 質(zhì)粒pSV3 neo酶切鑒定
質(zhì)粒pSV3 neo經(jīng)EcoR I酶切,可見8.8 kb片段,經(jīng)Hind III酶切,可見4.6、3.1、1.4、0.64 kb幾個(gè)片段(圖2)。
2.3 永生化M f胚胎成纖維細(xì)胞系的建立

注:M.DNA marker;1.質(zhì)粒pSV3neo經(jīng)Hind III酶切;2.質(zhì)粒pSV3neo經(jīng)EcoR I酶切;3.pSV3neo質(zhì)粒。圖2質(zhì)粒pSV3 neo酶切鑒定結(jié)果Note:M.DNA marker;1.pSV3neo plasmid digested with Hind III;2.pSV3neo plasmid digested with EcoR I;3.pSV3neo p lasmidFig.2 Restriction endonuclease digestion analysis of the p lasmids
利用脂質(zhì)體轉(zhuǎn)染試劑LipofectamineTM2000將質(zhì)粒pSV3 neo導(dǎo)入第3代M fEF內(nèi),經(jīng)G418篩選,第6天即長(zhǎng)出M fEF陽(yáng)性克隆(圖3,彩插3)。陽(yáng)性克隆經(jīng)擴(kuò)大培養(yǎng)目前已穩(wěn)定傳代4個(gè)月,經(jīng)鑒定SV40 T抗原已整合到M fEF中且穩(wěn)定表達(dá),細(xì)胞系命名為M fEF永生化細(xì)胞系。細(xì)胞按常規(guī)方法凍存,液氮凍存3個(gè)月后解凍復(fù)蘇細(xì)胞接種存活率達(dá)80%以上。M fEF永生化細(xì)胞系目前已連續(xù)傳代50代。
2.4 SV40 T基因在永生化細(xì)胞系中的整合及表達(dá)
對(duì)第5代和第20代M fEF永生化細(xì)胞系基因組DNA進(jìn)行PCR,質(zhì)粒pSV3 neo作陽(yáng)性對(duì)照,產(chǎn)物進(jìn)行1%瓊脂糖凝膠電泳分析。結(jié)果在391 bp處有一特異擴(kuò)增條帶,與陽(yáng)性對(duì)照條帶位置相同,而未轉(zhuǎn)染的細(xì)胞基因組DNA中無(wú)擴(kuò)增條帶(圖4)。
2.5 RT-PCR結(jié)果
陽(yáng)性細(xì)胞在391 bp處的特異擴(kuò)增條帶與陽(yáng)性對(duì)照條帶位置相同,而未轉(zhuǎn)染細(xì)胞未見該條帶,內(nèi)參引物為β-actin,擴(kuò)增產(chǎn)物為260 bp(圖5)。
2.6 未轉(zhuǎn)染和轉(zhuǎn)染后M fEF生長(zhǎng)情況比較
進(jìn)行了永生化M fEF與非永生化M fEF生長(zhǎng)速率間的比較,在15%血清濃度培養(yǎng)基中,前者在培養(yǎng)第2天即進(jìn)入指數(shù)增長(zhǎng)期,培養(yǎng)第2~4天達(dá)到生長(zhǎng)高峰,隨培養(yǎng)時(shí)間延長(zhǎng)細(xì)胞逐漸衰亡,細(xì)胞生長(zhǎng)的每一階段特征較明顯(圖6)。
3 討論
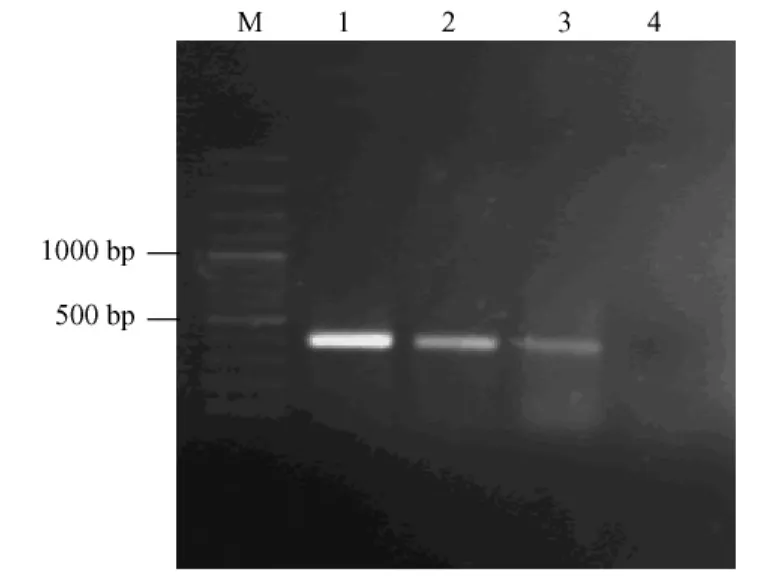
注:M.DNA marker;1.pSV3neo質(zhì)粒擴(kuò)增片段;2.第5代M fEF擴(kuò)增片段;3.第20代M fEF擴(kuò)增片段;4.未轉(zhuǎn)染M fEF擴(kuò)增片段圖4 PCR產(chǎn)物瓊脂糖凝膠電泳結(jié)果Note:M.W ide range DNA ladde rmarker;1. pSV3neo p lasmid;2.The 5 th passage M fEF cells;3. The 20 th passage M fEF cells;4.untransfected cellsFig.4 Agarose gel analysis of the PCR products
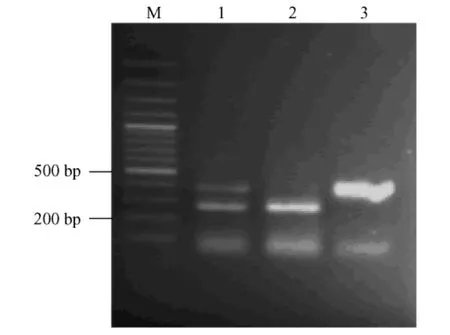
注:M.DNA marker;1.陽(yáng)性M fEF擴(kuò)增片段;2.陰性M fEF擴(kuò)增片段;3.pSV3neo質(zhì)粒擴(kuò)增片段圖5 SV40 T抗原mRNA在M fEF中的表達(dá)Note:M.DNA marker;1.MF embryonic fibroblasts of cell clones;2.untransfected cells;3.pSV3neo plasmidFig.5 RT-PCR analysis of SV40 T antigen mRNA in the M fEF cells
前期實(shí)驗(yàn)結(jié)果表明:體外培養(yǎng)的M fEF增殖能力較弱,生長(zhǎng)較緩慢,一般只能傳代培養(yǎng)8~9代,極大地限制了M fEF的相關(guān)應(yīng)用研究。細(xì)胞永生化是目前組織工程學(xué)領(lǐng)域研究的熱點(diǎn)。SV40(sim ian virus 40)是20世紀(jì)60年代初發(fā)現(xiàn)并分離出的猿猴腎細(xì)胞病毒,具有轉(zhuǎn)化動(dòng)物細(xì)胞和誘發(fā)腫瘤的特性[5]。SV40 T基因目前被廣泛用于轉(zhuǎn)基因動(dòng)物模型的建立和各種人類及動(dòng)物細(xì)胞的永生化,SV40 T
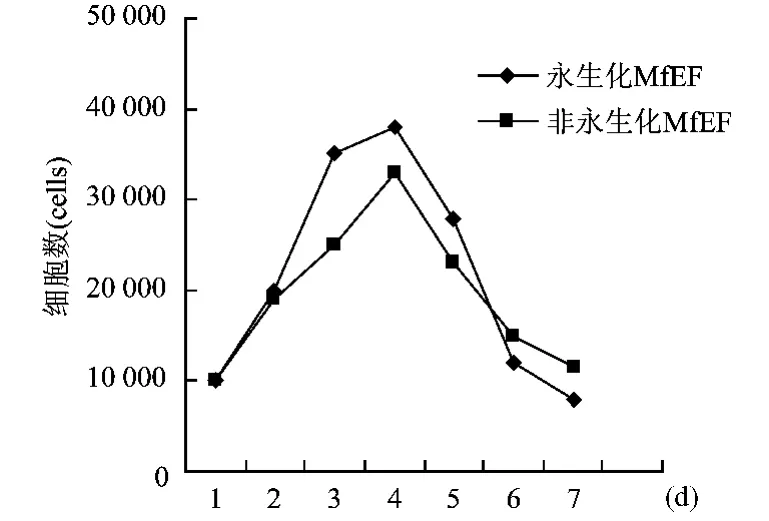
圖6 未轉(zhuǎn)染和轉(zhuǎn)染后M fEF生長(zhǎng)情況比較Fig.6 Comparison of the growth of pSV3neo plasmid-transfected and nontransfected M fEF cells
抗原基因的導(dǎo)入能加快轉(zhuǎn)化細(xì)胞的生長(zhǎng)速率,同時(shí)也能保留其原始細(xì)胞的許多分化表型,可作為體外研究原始細(xì)胞的標(biāo)準(zhǔn)細(xì)胞模型[6,7]。本研究將含有SV40 T基因的質(zhì)粒轉(zhuǎn)染體外培養(yǎng)的M fEF,建立永生化細(xì)胞系,永生化M fEF與非永生化M fEF相比,可見較多的細(xì)胞分裂相和增殖細(xì)胞,細(xì)胞傳代后貼壁速度較原代細(xì)胞慢;傳代、凍存和復(fù)蘇均不改變細(xì)胞形態(tài)和增殖能力。在實(shí)驗(yàn)中我們發(fā)現(xiàn):永生化M fEF傳代培養(yǎng)時(shí)對(duì)細(xì)胞密度和胰酶消化時(shí)間較敏感,如果細(xì)胞傳代密度過(guò)稀,胰酶消化時(shí)間過(guò)短,則M fEF在增殖過(guò)程中會(huì)出現(xiàn)小囊泡狀物,懸浮于培養(yǎng)液中,如果細(xì)胞生長(zhǎng)過(guò)密也會(huì)出現(xiàn)這種情況,通過(guò)更換培養(yǎng)液和及時(shí)傳代可消除此現(xiàn)象。本研究顯示:利用SV40 T基因使M fEF永生化是一種可行、可重復(fù)的建系方法。M fEF永生化細(xì)胞系的建立可為M f的相關(guān)研究提供細(xì)胞、蛋白質(zhì)、DNA、RNA樣本,與活體樣本相比,細(xì)胞系可以連續(xù)培養(yǎng),反復(fù)凍存,便于管理和使用,而且不會(huì)因?yàn)閭€(gè)體遺傳差異和生理狀態(tài)影響實(shí)驗(yàn)的可重復(fù)性,為M fEF長(zhǎng)期培養(yǎng)和全面研究東方田鼠抗日本血吸蟲機(jī)制奠定了基礎(chǔ)。
(本文圖1、3見彩插3)。
[1]李浩,何艷燕,林嬌嬌,等.東方田鼠抗日本血吸蟲現(xiàn)象的觀察[J].中國(guó)獸醫(yī)寄生蟲病,2000,8(2):12-15.
[2]陳輝,金輝,杜春艷,等.SV40T胃壁細(xì)胞定位表達(dá)轉(zhuǎn)基因小鼠的建立[J].解剖學(xué)報(bào),2008,39(5):699-702.
[3]劉梅冬,張華莉,龔環(huán)宇,等.HSF1基因敲除小鼠胚胎成纖維細(xì)胞的永生化[J].中南大學(xué)學(xué)報(bào)(醫(yī)學(xué)版)2006,31(2): 174-177.
[4]成鋼,蓋楠,熊德慧,等.東方田鼠胚胎成纖維細(xì)胞的分離培養(yǎng)及生物學(xué)特性[J].中國(guó)實(shí)驗(yàn)動(dòng)物學(xué)報(bào)2010,18(3):254-257.
[5]Chen W,Hahn WC.SV40 early region oncoproteins and human cell transformation[J].Histol Histopathol,2003,18:541-550.
[6]Poulin DL,Kung AL,Decaprio J A.p53 targets simian virus 40 large T antigen for acetylation by CBP[J].J Virol,2004,78: 8245-8253.
[7]Sullivan CS,Baker AE,Pipas JM..Simian virus 40 infection disrupts pl30-E2F complexes but does not perturb pRbE2F complexes[J].Virology,2004,320:218-228.
Estab lishm ent of an Imm ortalized M icrotus fortis Em bryonic Fibrob last Cell Line
CHENG Gang,LUO Sai-qun,GAI Nan,XIONG De-h(huán)ui,LI Rong,HU Wei-xin
(Molecular Biology Research Center,School of Biological Science and Technology,Central South University,Changsha 410078,China)
Objective To establish an immortalized Microtus fortis embryonic fibroblast(M fEF)cell line in order to lay the foundation for further studies on M icrotus fortis against Schistosma japonicum infection and for comparative study of fibroblasts from different animals.M ethods A mammalian expression vector(pSV3 neo)containing the SV40 large T antigen was used to transfect the 3rd passage M fEF cells using LipofectamineTM2000.Colonies were screened by G418 and expanded into immortalized cell lines.PCR was used to detect the integration of the large T antigen with genome in the Microtus fortis embryonic fibroblasts.Expression of SV40 large T antigen gene in the expanded cells was identified by RTPCR.ResultsStable growth and serial propagation were observed in the immortalized M icrotus fortis embryonic fibroblast line.The mRNA of SV40 T antigen gene was expressed in cells of the immortalized cell line.ConlusionAn immortalized cell line of M icrotus fortis embryonic fibroblasts has been successfully established.
M icrotus fortis;Embryonic fibroblasts;Immortalization,SV40 T antigen
Q25
A
1005-4847(2010)04-0292-04
2010-01-08
國(guó)家自然科學(xué)基金項(xiàng)目(30400256);湖南省研究生科研創(chuàng)新項(xiàng)目(No.2340-74236000001)資助。
成鋼(1976-),男,講師,博士研究生,研究方向:東方田鼠抗日本血吸蟲抗性相關(guān)基因篩選。E-mail:chenggang876@126. com。
胡維新(1950-),男,教授,博士生導(dǎo)師。E-mail:weixinhu@yahoo.com.cn
