長爪沙鼠線粒體DNA控制區全序列測定及分析
2010-09-08 08:14:12李長龍盧領群郭紅剛柯賢福戴方偉薩曉嬰
中國比較醫學雜志 2010年4期
李長龍,盧領群,郭紅剛,柯賢福,戴方偉,薩曉嬰
(浙江省醫學科學院 浙江省實驗動物中心,杭州 310013)
研究報告
長爪沙鼠線粒體DNA控制區全序列測定及分析
李長龍,盧領群,郭紅剛,柯賢福,戴方偉,薩曉嬰
(浙江省醫學科學院 浙江省實驗動物中心,杭州 310013)
目的對長爪沙鼠線粒體DNA控制區全序列進行測定,并對其進行鑒定及進化分析。方法根據長爪沙鼠已知基因序列設計引物,采用PCR產物測序法,對所得的片段進行測序鑒定。結合已公布嚙齒類動物D-loop區序列,分析其堿基組成、遺傳距離、并基于最小進化法和UPGMA法構建系統進化樹。結果獲得長爪沙鼠D-loop區序列,其與家鼠、小家鼠和倉鼠平均同源性為58%;堿基組成分析顯示,長爪沙鼠與嚙齒類動物有相似的堿基組成和堿基偏離,其A-skew和G-skew分別為0.0047和-0.28。進化分析結果顯示,長爪沙鼠與家鼠(0.35)、黑家鼠(0.38)和倉鼠(0.39)具有較近的遺傳距離,其分化順序為跳鼠、蔗鼠、長爪沙鼠、倉鼠、家鼠和小家鼠。結論
本研究獲得長爪沙鼠D-loop區全序列,確定了長爪沙鼠與倉鼠、家鼠、小家鼠及其它嚙齒動物的進化關系,為長爪沙鼠進化研究、線粒體的結構和功能研究奠定基礎。
長爪沙鼠;D-loop區;序列測定;進化分析
線粒體DNA(mitochondrial DNA,mtDNA)是高等動物唯一的核外遺傳物質,呈共價閉合的環狀雙鏈結構。與核基因組相比,線粒體基因組結構簡單、進化速度快(是單拷貝核基因的5~10倍),呈嚴格的母性遺傳,遺傳行為相對獨立,變異發生的幾率相對穩定、無組織特異性、提取方便[1]。由于mtDNA的這些獨特遺傳特性,已被廣泛地用于物種起源與進化、生物分類、及群體遺傳結構等方面的研究[2-4]。在進化分析時,研究者往往選擇不同的區域進行不同時間尺度的進化分析。線粒體控制區(D-loop region)是mtDNA中進化最快、變化最復雜的區域,常用于種內或種間遺傳分化研究[5]。對線粒體D-loop區結構和功能的研究不僅將有助于了解DNA復制、轉錄的機制和進化規律研究,而且對線粒體結構和功能研究也具有很大意義[6]。
長 爪 沙 鼠 (Mongolian gerbil, Meriones unguiculatus)俗稱蒙古沙鼠,屬于嚙齒目,倉鼠科,沙鼠亞科,沙鼠屬,又稱長爪沙土鼠、蒙古沙鼠和黃耗子等,野生長爪沙鼠主要分布于我國內蒙古及其毗鄰的干旱和半干旱地區[7]。1935年大連衛生所的春日送給日本北里研究所20對長爪沙鼠并開始馴化,后引種到美、英、法等國。日本國家實驗動物中心(NIBIO)現已培育成3個長爪沙鼠近交系[8]。我國有兩個主要的長爪沙鼠群體,分別保存于浙江省實驗動物中心和首都醫科大學[7]。長爪沙鼠具有獨特的解剖學、生理學和行為學性狀,對于一些疾病(如:腦缺血、癲癇、高血脂、寄生蟲、細菌、病毒和老年性疾病等)的研究具有極為重要的價值[9-11]。長爪沙鼠被認為是研究脂質代謝良好的模型動物[7,8,12,13],在越來越多的研究所使用。但是,國內外對長爪沙鼠線粒體D-loop區的研究還未見報道。本研究對長爪沙鼠線粒體 D-loop區全序列進行測定和鑒定,并結合已公布嚙齒類動物D-loop區序列進行系統進化分析,旨在為長爪沙鼠系統進化關系提供遺傳學資料,為全長線粒體測定分析奠定基礎。
1 材料和方法
1.1 材料
長爪沙鼠來自浙江省實驗動物中心(生產許可證號:SYXK(浙)2008-033;使用許可證號:SYXK (浙)2008-0014),長爪沙鼠解剖采集肝臟樣品,-20℃保存。
1.2 總DNA提取
采用酚氯仿抽提法提取基因組DNA,0.7%瓊脂糖凝膠電泳檢測,-20℃保存。
1.3 引物設計和PCR擴增
分別參照長爪沙鼠 mtDNA序列 AB381901和AJ851249設計上下游引物[14,15],引物由上海生工合成。上游(cyt B 3′端)5’-ATCGGACAAGTCGCT TCAAT-3’,下游(12S 5′端)5’-AGCGATGGCTCG TAGTTCTC-3’。
擴增反應體系總體積為25 μL:模板DNA 100 ng、10×buffer 2.5 μL、MgC12(2.5 mol/L)2 μL、dNTP 2 μL、上下游引物(10 pmol/L)各1 μL、1 U pfuDNA聚合酶,加滅菌純水補足。擴增條件:94℃預變性5 min,94℃變性30 s,56.5℃復性30 s,72℃延伸1 min,30個循環,72℃延伸7 min。
1.4 序列純化、測序、拼接
用北京天根凝膠回收試劑盒對 PCR擴增產物進行純化,純化后的產物由上海美季公司完成測序工作,Chromas 2.22校對測序圖,DNAMAN5.5拼接序列。
1.5 序列的鑒定和分析

圖1 PCR擴增長爪沙鼠線粒體D-loop區Fig.1 The results of PCR amplified D-loop region in Mongolian gerbil.
從GenBank下載其他嚙齒類動物類mtDNA控制區序列,GenBank登錄號分別為:NC_005089家鼠(Mus musculus)、NC_012387小家鼠(Mus musculus castaneus)、NC_006914西歐家鼠(Mus musculus domesticus)、NC_006915日本小鼠(Mus musculus molossinus)、NC_010339東歐家鼠(Mus musculusmusculus)、NC_012389緬鼠(Rattus exulans)、NC_ 012461帥家鼠(Rattus praetor)、NC_012374黑家鼠(Rattus rattus)、NC_011638達氏家鼠 (Rattus tanezumi)、AC_000022褐家鼠(Rattus norvegicus)、NC_013068大倉鼠(Tscherskia triton)、NC_007936中國倉鼠(Cricetulus griseus)、NC_003041臺灣田鼠(Microtus kikuchii)、NC_001892胖睡鼠(Glis)、NC_ 005314非洲跳鼠(Jaculus)、NC_009056鱗尾松樹(Anomalurus)、NC_000884豚鼠(Cavia porcellus)和NC_002658蔗鼠(Thryonomys swinderianus)等18條全長 mtDNA序列中 D-loop序列進行分析。用DNAMAN5.5進行同源性分析;用 EMBOSS中compseq程序統計堿基含量和計算A和G偏離[16];用MEGA 4.1統計物種間遺傳距離,并基于Kimura-Parameter雙參數模型,用UPGMA和最小進化法構建分子系統進化樹[17]。
2 結果與分析
2.1 D-loop區序列組成
PCR產物電泳結果顯示,特異性擴增約1.5 kb的片段(圖1)。除去兩端不準確的部分,共測得長爪沙鼠mtDNA序列1 677 bp,其中包括D-Loop區全序列、tRNA-Phe、tRNA-Val、tRNA-Thr、tRNA-Pro基因部分序列。序列分析顯示,長爪沙鼠線粒體 D-loop區序列全長為997 bp(圖2),堿基A、C、G、T的含量分別為32.1%、23.2%、12.9%、31.8%;A +T(63.9%)含量大于G+C(46.1%)含量。A-skew的范圍為0.15(松鼠)至-0.034(臺灣田鼠); G-skew的范圍為-0.17(豚鼠)至 -0.41(睡鼠)。長爪沙鼠的 A-skew和 G-skew分別為 0.0047和-0.28,A和G偏離明顯(圖3)。
2.2 長爪沙鼠D-loop區序列鑒定
長爪沙鼠與其他嚙齒動物的 D-loop序列進行同源性分析,結果顯示,家鼠(97%)、小家鼠(83%)和倉鼠(68%)動物均具有較高同源性,長爪沙鼠與上述三種動物的同源性為 58%。鑒于 D-loop是DNA中進化最快的部分,說明該序列為長爪沙鼠線粒體D-loop區(圖4)。
2.3 系統進化樹的建立及遺傳距離計算

圖2 長爪沙鼠D-loop區序列Fig.2 The complete sequence of D-loop region in Mongolian gerbil
基于D-loop區全序列,用鄰接法和最小進化法構建相關動物系統進化樹得到基本一致的拓撲結構(圖5,圖6)。結果顯示,進化樹主要有了兩個分支,A分支由家鼠、小家鼠、倉鼠、長爪沙鼠、蔗鼠和睡鼠構成;在A分支內,家鼠、小家鼠、倉鼠分別聚合后依次與長爪沙鼠、蔗鼠和跳鼠聚合。家鼠、小家鼠、西歐家鼠、日本小鼠和東歐家鼠集中在小家鼠分支上。緬鼠、帥家鼠、黑家鼠、達氏家鼠和褐家鼠集中在家鼠分支上。大倉鼠、中國倉鼠和臺灣田鼠集中在倉鼠分支上。B分支由松鼠、豚鼠和跳鼠聚合而成。結果說明嚙齒動物的線粒體控制區序列包含了重要的系統發育信息。枝長表示分歧度,枝上的數值為 1 000次重復抽樣檢驗得到的支持率。選擇家鼠、黑家鼠和中國倉鼠分別代表家鼠、小家鼠和倉鼠與長爪沙鼠計算遺傳距離。結果顯示(表1),長爪沙鼠與黑家鼠遺傳距離最近為0.35;與家鼠遺傳距離為0.37;與中國倉鼠遺傳距離為0.39。結果表明長爪沙鼠與黑家鼠最近的親緣關系,與家鼠較近的親緣關系,與豚鼠親緣關系最遠為0.49。
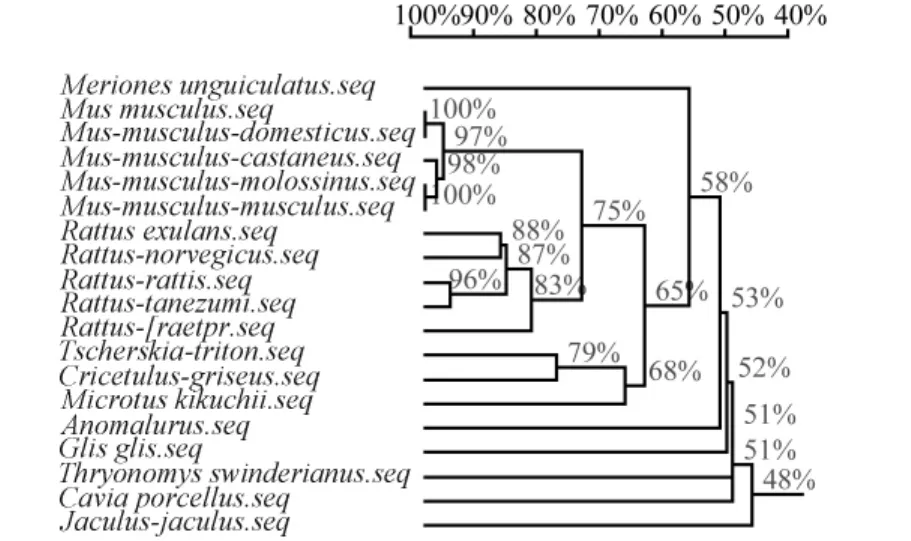
圖4 長爪沙鼠與其他嚙齒類動物的同源性分析Fig.4 The homology analysis between Mongolian gerbil and other rodents
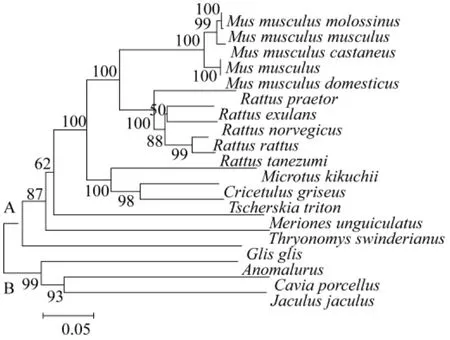
圖5 M-E法構建的D-loop基因系統發生樹Fig.5 Phylogenetic tree based on D-loop region sequences generated by minimum-evolution methods.
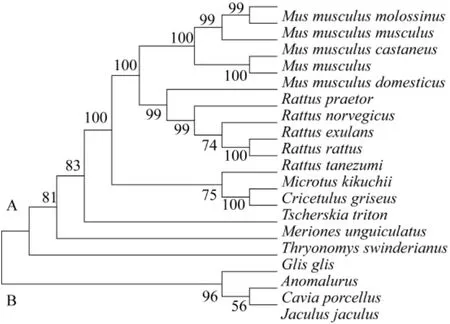
圖6 UPGMA法構建的D-loop基因系統發生樹Fig.6 Phylogenetic tree based on D-loop region sequences generated by UPGMA methods.
3 討論
3.1 長爪沙鼠D-loop區序列的測定和鑒定
mtDNA是閉合的雙鏈環狀DNA,D-loop區位于mtDNA復制起點(終點)前,在cyt B和12S基因之間。本研究根據已有的長爪沙鼠線粒體cyt B和12S基因序列分別設計上下游引物,并在線粒體基因組DNA中擴增出約1.5 kb的DNA片段(圖1)。經過重復雙向測序,結果拼接后獲得長1 677 bp的DNA片段。序列中去除轉運RNA后獲得997 bp序列。為了鑒定該序列,本研究選擇18種不同的嚙齒類動物線粒體D-loop區序列進行同源性分析,結果顯示,長爪沙鼠與家鼠、小家鼠和倉鼠的平均同源性為58%。D-loop區是mtDNA中高度變異的非編碼區,進化速率是mtDNA其它區域的3~5倍[18],據此,該序列為長爪沙鼠全長D-loop區序列。
3.2 長爪沙鼠D-loop區序列及進化分析
動物的起源進化長期以來一直困擾著遺傳學家。分子遺傳學方法為研究群體的起源和進化開辟了新的途徑[19]。mtDNA具有很多的特性,利用mtDNA序列構建的系統發生數能很好的反映動物母系的遷徙和進化。mtDNA比核DNA的突變率高5~10倍,其中的 D-loop區是線粒體基因組進化速率最快的區域,進化速率是mtDNA其他區域的3~5倍[18]。許多科學家利用 mtDNA序列差異來研究動物的遺傳多樣性與起源演化。Giufra對歐洲野豬、歐洲家豬、亞洲野豬和中國梅山豬 mtDNA D-1oop區進行分析,發現家豬有歐洲野豬和亞洲野豬兩個母系起源并證實歐洲大白豬是亞洲豬和歐洲豬的雜交種,該結果與18~19世紀初亞洲豬被引入歐洲的歷史記載相符[20]。在近年來,動物 mtDNA研究取得了一系列令人矚目的成果。
不同物種的mtDNA堿基組成和特性存在明顯的差異[21]。DNA的鳥嘌呤和胞嘧啶的堿基含量(G +C含量)在不同物種的基因組中波動范圍廣闊[22,23],甚至在同一物種的基因組不同區域也不相同。這種基因組局部堿基不均衡的現象表現為不同區域的核苷酸堿基組成存在差異[24]。長爪沙鼠 D-loop區序列A、C、G、T的含量分別為32.1%、23.2 %、12.9%、31.8%,其中G的含量顯著低于其他堿基的含量,這是mtDNA的一個顯著特征[25];A+ T(63.9%)含量大于G+C(46.1%)含量,與其他哺乳動物 A、T含量高,G、C含量低的特點相似[26]。長爪沙鼠的 A-skew和 G-skew分別為0.0047和 -0.28(圖3)。與長爪沙鼠遺傳距離較近的動物在堿基組成和A和G偏離上較為接近,具有明顯的種屬特異性。
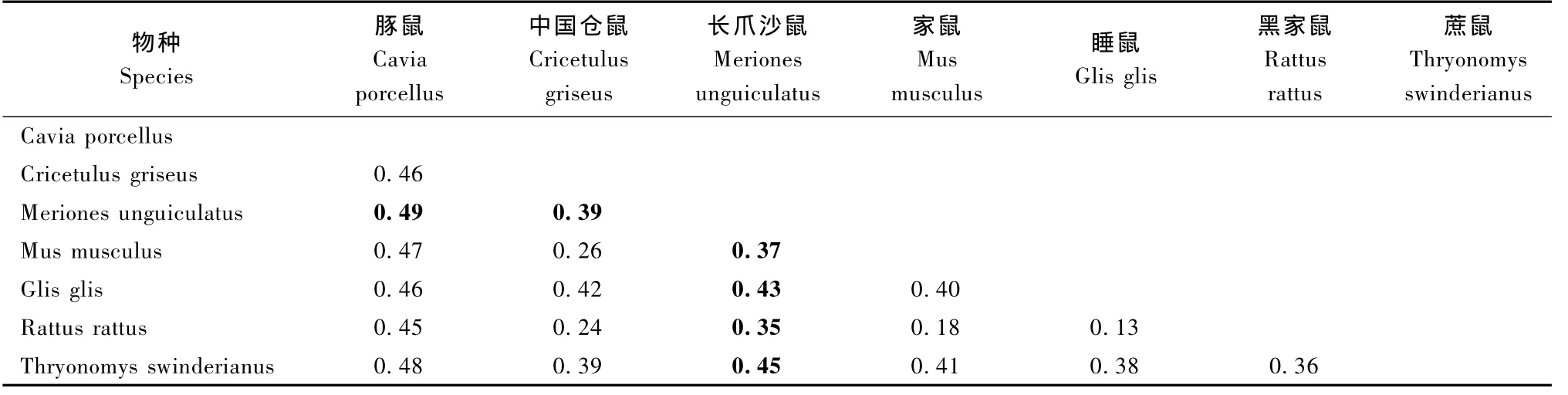
表1 長爪沙鼠與其他鼠類遺傳距離Tab.1 The genetic distance between Mongolian gerbil and other rodents
Chevret等曾利用DNA在溶解溫度的方法研究長爪沙鼠與家屬和小家鼠的進化關系,結果顯示長爪沙鼠與家屬和小家鼠存在較相近的進化關系[27]。Colangelo等利用 cyt B和16S rRNA序列研究非洲沙鼠的分子進化關系[28];Chevret等利用 cyt B和12S rRNA基因序列研究沙鼠亞科內分子進化關系[29]。本研究應用MEGA 4.1軟件,基于D-loop區全序列,用鄰接法和最小進化法構建系統進化樹得到基本一致的拓撲結構(圖5,6),結果顯示,家鼠、小家鼠和倉鼠等動物均分別各自聚合到同一個分支上來,之后依次與長爪沙鼠、蔗鼠和睡鼠聚合在一起。結果表明,長爪沙鼠與家鼠、小家鼠和倉鼠有具有相近的親緣關系。遺傳距離計算結果驗證了進化分析結果,長爪沙鼠與褐家鼠最近的親緣關系,說明家鼠較近的親緣關系,與豚鼠親緣關系最遠。其分化順序為跳鼠、蔗鼠、長爪沙鼠、倉鼠、家鼠和小家鼠。以上數據根據動物單一個體的D-loop區序列計算得出,進一步研究應增加每種動物數量,并綜合線粒體基因組信息和考古學等其他數據綜合分析。
3.3 長爪沙鼠線粒體研究展望
線粒體是細胞的“能量工廠”,多數細胞內90%的氧消耗在線粒體內進行[30]。線粒體序列的測定將為分子進化、能量代謝及相關疾病研究奠定基礎。長爪沙鼠是一種正在廣泛應用的多功能實驗動物。特別是長爪沙鼠的脂質代謝與其它實驗動物存在明顯差異,但是與人類又有很大的相似之處,是研究人類脂質代謝良好的動物模型[7,8,12,13]。長爪沙鼠線粒體 D-loop區序列的測定和進化分析不但為長爪沙鼠進化研究提供基礎數據,而且為獲得全長mtDNA序列奠定基礎,為長爪沙鼠線粒體的結構和功能研究及廣泛應用提供保障。
[1] 徐其放,陳嘉昌,朱世杰,等.動物線粒體DNA的特異結構及應用分子系統學分析的方法[J].中國比較醫學雜志,2005,15(5):315-319.
[2] 廖順堯,魯成.動物線粒體基因組研究進展[J].生物化學與生物物理進展,2000,27(5):508-512.
[3] 趙興波,李寧,吳常信.動物線粒體核質基因互作的研究進展[J].遺傳,2001,23(1):81-85.
[4] Nishibori M,Shimogiri T,Hayashi T,et al.Molecular evidence for hybridization of species in the genus Gallus except for Gallus varius[J].Animal Genetics,2005,36(5):367-375.
[5] Donne-Goussé C,Laudet V,Hanni C.A molecular phylogeny of anseriformes based on mitochondrial DNA analysis[J].Mol Phylogenet Evol,2002,23(3):339-356.
[6] 孫玉華,王偉,劉思陽,等.中國胭脂魚mtDNA控制區遺傳多樣性分析[J].遺傳學報,2002,29(9):787-790.
[7] 方喜業,邢瑞昌,賀爭鳴.實驗動物質量控制[M].北京:中國標準出版社,2008:pp 913-916.
[8] 丁賢明,錢寶珍,Junichiro M,等.長爪沙鼠的遺傳多樣性分析[J].遺傳,2008,30(7):877-884.
[9] Hegsted DM,Gallagher A.Dietary fat and cholesterol and serum cholesterol in the gerbil.J Lipid Res,1967,8(3):210-214.
[10] Tovar-palacio C,Potter SM,Hafermann JC,et al.Intake of soy protein and soy protein extracts influences lipid metabolism and hepatic gene expression in gerbils[J].J Nutr,1998,128(5): 839-842.
[11] Forsythe WA 3rd.Comparison of dietary casein or soy protein effects on plasma lipids and hormone concentrations in the gerbil (Meriones unguiculatus)[J].J Nutr,1986,116(7):1165-1171
[12] Ohkusa T,Okayasu I,Miwa H,et al.Helicobacter pylori infection induces duodenitis and superficial duodenal ulcer in Mongolian gerbils[J].Gut,2003,52(6):797-803.
[13] 鐘民濤,黃敏,盧靜,等.長爪沙鼠速發型高脂血癥模型的初步建立[J].中國實驗動物學報,2006,14(3):217-221.
[14] Martin Y,Gerlach G,Schlotterer C,et al.Molecular phylogeny of European muroid rodents based on complete cytochrome b sequences[J].Mol Phylogenet Evol,2000,16(1):37-47.
[15] Chevret P, Dobigny G. Systematicsand evolution ofthe subfamily Gerbillinae(Mammalia,Rodentia,Muridae)[J]. Mol Phylogenet Evol,2005,35(3):674-688.
[16] Salvato P,SimonatoM,BattistiA,etal. Thecomplete mitochondrial genome of the bag-shelter moth Ochrogaster lunifer (Lepidoptera,Notodontidae)[J].BMC Genomics,2008,15 (9):331-346.
[17] Tamura K,Dudley J,Nei M,et al.MEGA4:Molecular Evolutionary Genetics Analysis(MEGA)software,version 4.0[J].Mol Biol Evol.2007,24(8):1596-1599.
[18] Wenink PW,Baker AJ,Tilanus MG.Mitochondrial control region sequences in two shorebird species:the Tumstone and the dunlin,and their utility in population genetic studies[J].Mol Biol Evol,1994,11(1):22-31.
[19] Garrigan D,Hammer MF.Reconstructing human origins in the genomic era[J].Nat Rev Genet,2006,7(9):669-680.
[20] Giuffra E,Kijas J MH,Amarger V,et al.The origin of the domestic pig:independent domestication and subsequent introgression[J].Genetics,2000,154(4):1785-1791.
[21] 高英凱,苗永旺,蘇小茜,等.74種鳥類線粒體基因組堿基組成及特征分析[J].云南農業大學學報:自然科學版,2009,24(1):51-58.
[22] Belozersky AN, Soirin AS. A correlation between the compositions of deoxyribonucleic and ribonucleic acids[J]. Nature,1958,182(4628):111-112.
[23] Daubin V,Perrière G.G+C3 structuring along the genome:a common feature in prokaryotes[J].Mol Biol Evol,2003,20 (4):471-483.
[24] Sueoka N.Two aspects of DNA base composition:G+C content and translation-coupled deviation from intra-strand rule of A=T and G=C[J].Molecular Evolution,1999,49(1):49-62.
[25] 樊乃昌,施銀柱.中國鼢鼠(Eospalax)亞屬分類研究[J].獸類學報,1982,2(2):183-197.
[26] 楊路存,陳桂琛,劉榮堂,等.3種鼢鼠mtDNA控制區結構及其系統發育分析[J].草業科學,2009,26(2):100-106.
[27] Chevret P,Denys C,Jaeger JJ,et al.Molecular evidence that the spiny mouse(Acomys)is more closely related to gerbils (Gerbillinae)than to true mice(Murinae).Proc Natl Acad Sci U S A,1993,90:3433-3436
[28] Colangelo P,Granjon L, Taylor PJ, etal. Evolutionary systematics in African gerbilline rodents of the genus Gerbilliscus:Inference from mitochondrial genes.Mol Phylogenet Evol,2007,42(3):797-806.
[29] Chevret P,Dobigny G. Systematics and evolution ofthe subfamily Gerbillinae(Mammalia,Rodentia,Muridae).Mol Phylogenet Evol,2005,35(3):674-688.
[30] Chance B,Sies H,Boveris A.Hydroperoxide metabolism in mammalian organs[J].Physiol Rev,1979,59(3):527-605.
Determination and Analysis of Mitochondrial DNA D-Loop Region Complete Sequence of Mongolian Gerbil(Meriones unguiculatus)
LI Chang-long,LU Ling-qun,GUO Hong-gang,SA Xiao-ying
(Zhejiang Center of Laboratory Animals,Zhejiang Academy of Medical Sciences,Hangzhou 310013,China)
ObjectiveTo determine the complete sequence of mitochondrial D-loop region of Mongolian gerbil based on the PCR products.The sequences were analyzed for phylogenetic tree,nucleotide composition and genetic distance。MethodsThe primer was designed according to the published partial sequences.The PCR products were sequenced and determined.Combined with the known D-loop region sequence of other rodents,nucleotide composition and genetic distance were analysed,and the phylogenetic tree was constructed by minimum-evolution(ME)methods and UPGMA methods。ResultsThere was a high homology(58%)between the sequences of Mongolian Gerbil and that of rat,mouse and hamster.The nucleotide composition of Mongolian Gerbil was similar with that of other rodents.The A-skew and G-skew were 0.0047 and -0.28,respectively.The results of phylogenetic tree analysis showed a higher genetic relationship with rat(0.35),mouse(0.38)and hamster(0.39).The order of differentiation was cane rat,desert jerboa,Mongolian gerbil,hamster,mouse and rat。Conclusions The mitochondrial D-loop region sequence of Mongolian gerbil has been determined.To our knowledge,it has not been reported in the literature before.In this study,the genetic relationship between Mongolian gerbil and other rodents has been analyzed.The results of this study may be of importance for studies on evolution,structure and function of the mitochondria in Mongolian gerbil.
Mongolian gerbil;D-loop region;Sequence determination;Phylogenetic analysis
R-33
A
1671-7856(2010)04-0040-06
2009-12-17
“十一五”科技支撐重點項目(2009BAI83B02);浙江省實驗動物與安全性研究重點實驗室(2008F3021)。
李長龍(1976-),男,助理研究員,博士,研究方向:實驗動物遺傳學,E-mail:li-changlong@126.com。
薩曉嬰(1952-),男,研究員,研究方向:實驗動物學,E-mail:saxiaoyin@163.com。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
