西葫蘆株型性狀主基因-多基因混合遺傳分析
2010-08-10 02:22:38陳鳳真何啟偉
中國蔬菜 2010年22期
關鍵詞:模型
陳鳳真 何啟偉
(1菏澤學院園林工程系,山東菏澤 274000;2山東省農業科學院蔬菜研究所,山東濟南 250100)
西葫蘆,即美洲南瓜(Cucurbita pepoL.)是冬春設施栽培的主要蔬菜作物之一。西葫蘆從生長習性來分,主要有蔓生、矮生和半蔓生(Kasrawi,1995)。株高和葉片數是西葫蘆重要的農藝性狀,它們與西葫蘆的產量及熟性性狀有密切的相關性,研究它們的基因效應,對西葫蘆的株型、產量及熟性性狀的遺傳改良有重要的指導意義(陳鳳真,2008)。陳鳳真等(2007)采用經典遺傳學對西葫蘆株高和葉片數進行了遺傳分析。經典遺傳學雖然能估計基因總體效應,但不能區分主基因、多基因效應。
本試驗在李建友(2004)研究的基礎上,利用蓋鈞鎰(2005)的主基因-多基因混合分析方法,對西葫蘆株型性狀的遺傳開展進一步的研究,旨在對西葫蘆雜種優勢產生的機理進行研究,并為開展西葫蘆的株型育種提供參考依據。
1 材料與方法
1.1 試驗材料
選用蔓生和矮生兩種不同株型,果形差異較大且經6代以上自交選育的3個西葫蘆自交系q-1(蔓生、白皮、長棒形)、23-4G(矮生、墨綠皮、粗短形)和A-7(矮生、深綠皮、粗短形)為試材。其中q-1和A-7分別來自奇臺和阿爾及利亞的引進品種,23-4G由雙豐特早自交分離選育而來。
1.2 試驗方法
試驗在山東農業大學蔬菜試驗站進行。2005年春配制F1,組合1:q-1(♀)×23-4G(♂),組合2:q-1(♀)×A-7(♂)。2005年10月在日光溫室中獲得F2、回交世代BC1(F1×q-1)和BC2(F1×23-4G 和 F1×A-7)。
2006年3月12日28 ℃恒溫催芽,3月16日播種,3月30日定植到拱圓大棚(長60 m,跨度10 m)。親本及F1各定植30株,回交世代BC1、BC2分別定植60株,F2定植90株,行距60 cm,株距50 cm;3次重復,隨機區組設計。單干整枝,落蔓管理,其他管理同一般生產。拔秧時測量株高及統計葉片數,并對數據進行統計分析。
1.3 統計分析方法
采用植物數量性狀主基因-多基因混合遺傳模型(蓋鈞鎰,2005)對西葫蘆6個世代的株型性狀進行聯合分析,通過極大似然法(MaximuMlikelihood method)和 IECM算法(Iterated expectation and conditional maximization)對混合分布中的有關成分分布參數做出估計;然后利用AIC準則(Akaike’s information criterion)選擇AIC值最小或較小的1個或幾個備選模型,再通過一組適合性測驗,共有5個統計量,即U12、U22、U32、nW2和Dn,其中U12、U22、U32為均勻性檢驗,nW2為Smirnov檢驗,Dn為Kolmogorov檢驗,根據結果選擇最優模型,即選擇統計量達到顯著水平個數最少的模型作為最優模型;并估計主基因和多基因效應值、主基因遺傳率等遺傳參數。分析軟件由南京農業大學國家大豆改良中心提供。
2 結果與分析
2.1 株高和葉片數測定值的次數分布
將各個世代的株高和葉片數次數分布分別列于表1、2。從表1、2可知,2個組合F1的株高性狀存在雜種優勢,2個組合的F1和F2葉片數的平均數低于低值親本的平均數,說明西葫蘆葉片數存在負向超親優勢。2個組合的F2株高均集中在150~350 cm之間,葉片數均集中在41~61片之間。由圖1~4可以看出,2個組合的F2株高和葉片數都呈單峰較明顯的偏態分布,表明主基因可能控制2個組合株高和葉片數的遺傳。
2.2 株型性狀遺傳模型
以2個組合的6個不同世代材料株高和葉片數觀測值為基礎,利用植物數量性狀主基因-多基因混合遺傳模型的多世代聯合分析方法對6個世代群體進行分析,計算不同遺傳模型的極大對數似然值和AIC值(表3)。
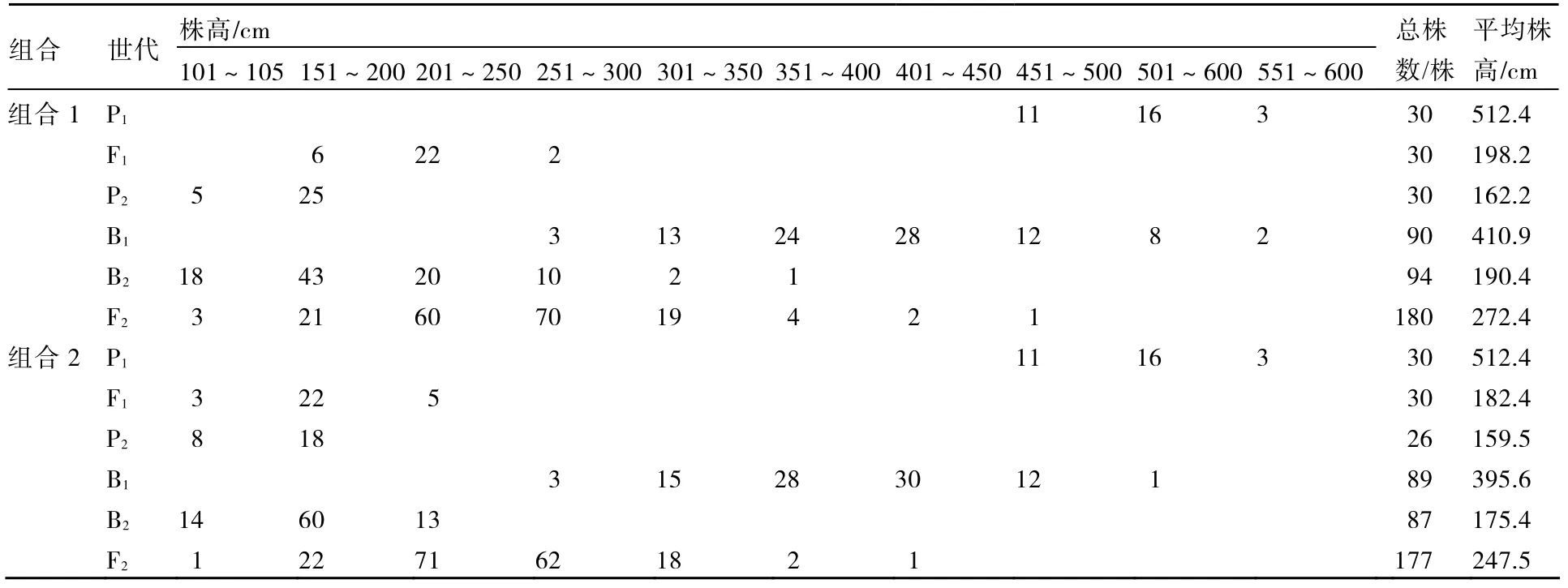
表1 各世代株高表型值次數分布

表2 各世代葉片數表型值次數分布

圖1 組合1的F2株高次數分布

圖2 組合2的F2株高次數分布
根據AIC準則,組合1的株高B-1模型和E模型AIC值較小,B-1和E模型可作為備選模型;組合2的株高B-1和E-1模型AIC值較小,B-1和E-1模型可作為備選模型;組合1與組合2的葉片數B-1模型AIC值最小,組合1葉片數的D-4模型和組合2葉片數的D-2、D-4模型AIC值與最小值接近,可作為備選模型。
進一步對株高和葉片數性狀的B-1模型與其他備選模型進行適合性檢驗,選擇統計量達到顯著水平個數較少的模型作為最優模型。表4、5結果表明:在6個世代的30個統計量中,組合1的株高B-1模型有5個統計量達到顯著水平,而E模型有15個統計量達到顯著水平;組合1的葉片數B-1模型達到顯著水平的統計量個數少于D-4模型;組合2的株高B-1模型的各統計量不顯著,葉片數B-1模型有1個統計量達到顯著水平,而株高和葉片數的其他備選模型均有10個以上的統計量達到顯著水平;因此2個組合的株高和葉片數B-1模型不僅最佳而且最適。
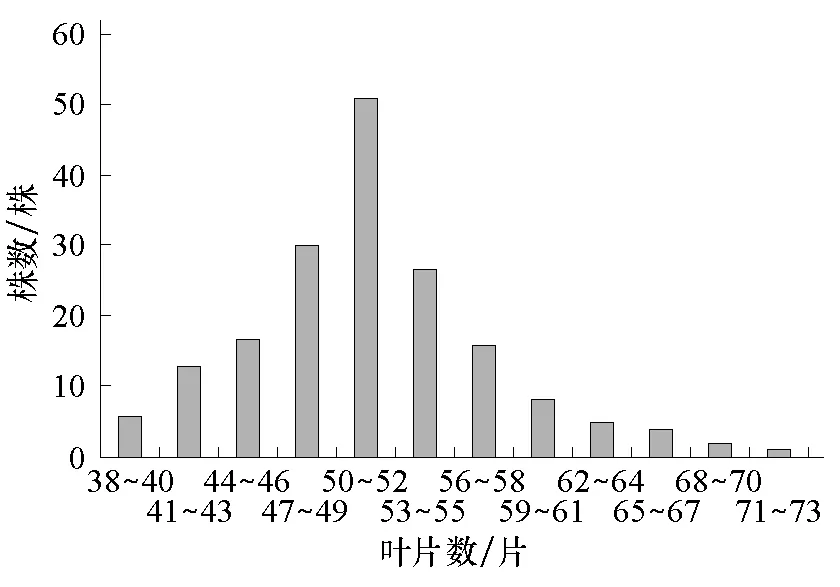
圖3 組合1的F2葉片數次數分布
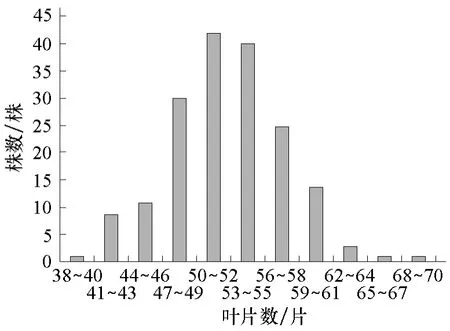
圖4 組合2的F2葉片數次數分布
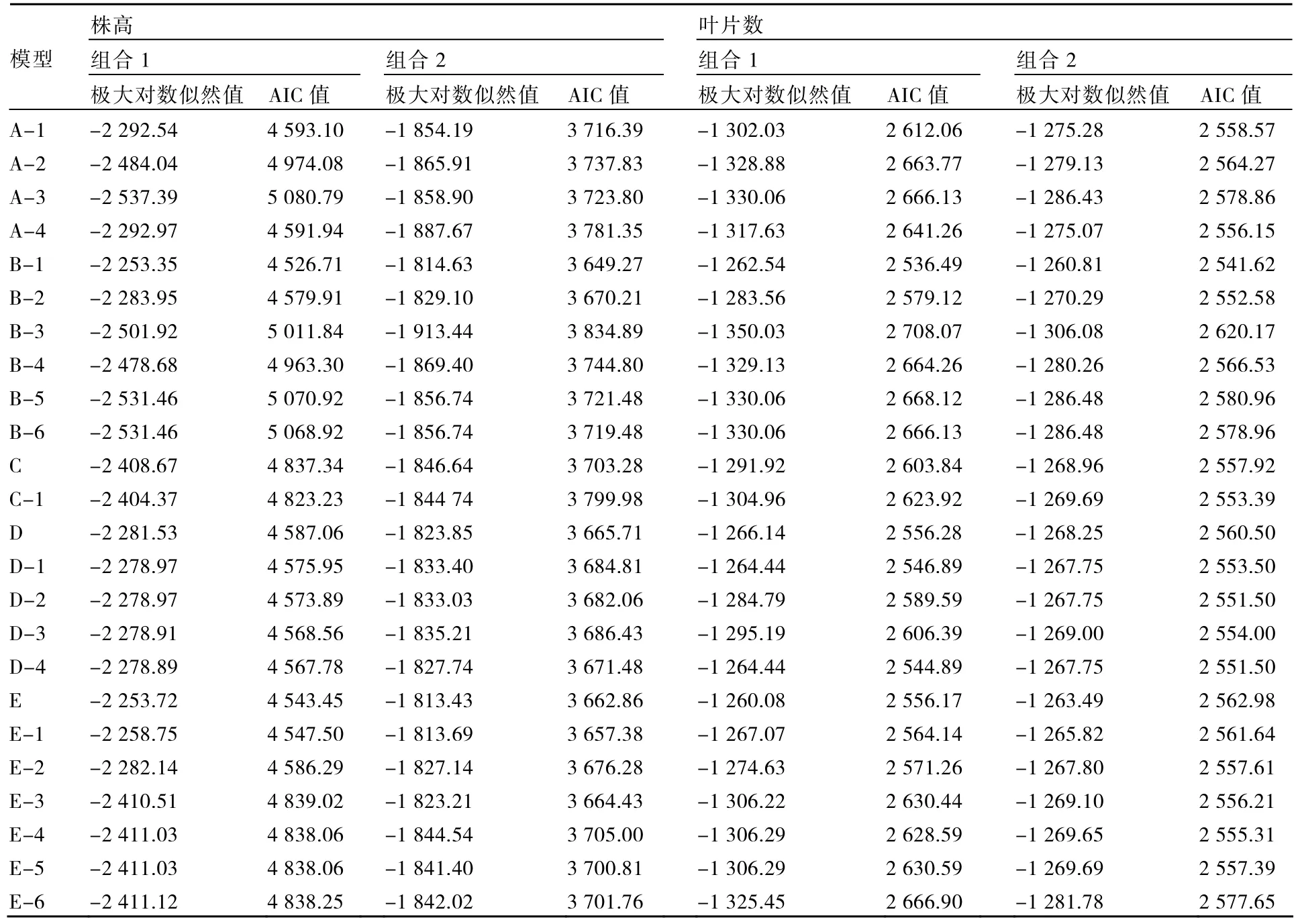
表3 株高和葉片數模型的極大對數似然值和AIC值
2.3 株型性狀遺傳參數估計
2.3.1 株高遺傳參數的估計 各組合株高性狀遺傳參數估計值列于表6。1階遺傳參數結果表明:組合1和組合2兩對主基因顯性效應值之和(ha+hb)分別為-93.60和-8.01;組合1與組合2的兩對主基因加性效應值之和(da+db)分別為151.89和 180.76;加性×加性上位性互作效應(i)分別為34.77和13.70,加性×顯性上位性互作效應之和(jab+jba)分別為13.71和8.67;2個組合的兩對主基因顯性效應值之和(ha+hb)均為負值,且小于兩對主基因加性效應值之和,故F1株高性狀無超親優勢;因此西葫蘆的株高性狀均以加性效應為主,其次均為加性×加性上位性互作效應;由表6中的2階參數估計值可知:組合1與組合2的分離后代的差異均是由主基因引起的,2個組合的F2株高的主基因遺傳率分別為67.00 %和74.21 %,分別還有33.00 %和25.79 %的變異是由環境因素決定的,說明環境對西葫蘆株高性狀有一定的影響,宜早代選擇。

表4 株高的適合性檢驗

表5 葉片數的適合性檢驗
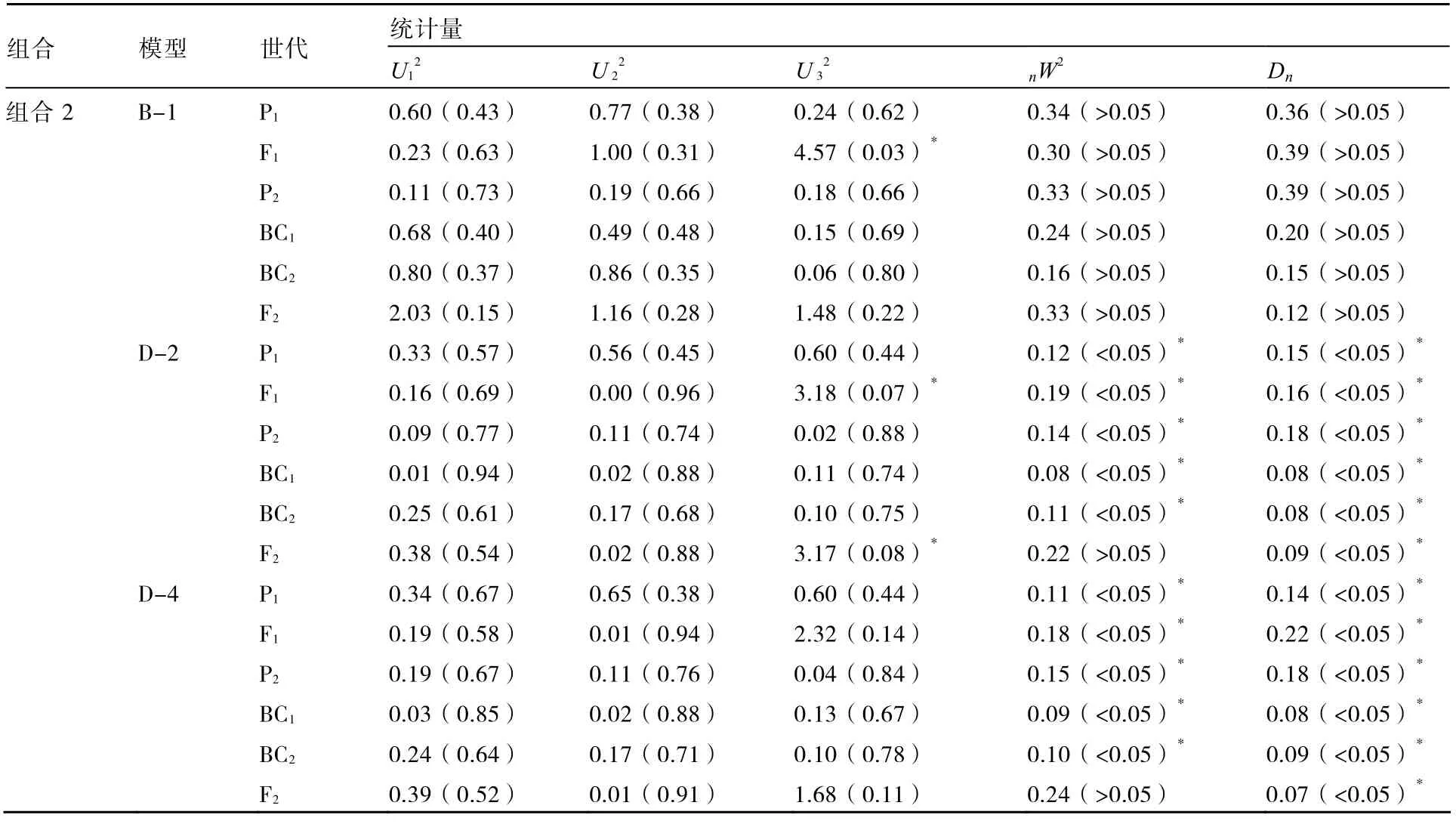
續表

表6 株高、葉片數的各遺傳參數估計值
2.3.2 葉片數性狀遺傳參數估計 將2個組合葉片數的適合模型下遺傳參數估計值列入表6。1階遺傳參數結果表明:組合1和組合2的主基因顯性效應值之和(ha+hb)分別為-2.04和-0.84,兩對主基因加性效應值之和(da+db)分別為-1.20和-0.63,加性×加性上位性(i)分別為-6.13和-1.52、加性×顯性上位性互作效應之和(jab+jba)分別為-8.89和-2.03,顯性×顯性上位性互作效應值(l)分別為-6.46和-1.31;2個組合的主基因顯性效應值之和為負值,說明F1葉片數性狀減少,表現負向超親優勢;在遺傳效應值中加性效應值之和較大,因此2個組合的葉片數性狀以加性和顯性效應為主。2階遺傳參數結果表明:組合1和組合2葉片數性狀的遺傳均由兩對主基因控制,組合1主基因在分離后代BC1、BC2與F2的遺傳率分別為:66.53 %、62.74 %和68.07 %;組合2的 BC1、BC2與F2的主基因遺傳率分別為:63.67 %、60.89 %和71.96 %,說明環境對西葫蘆葉片數性狀有一定的影響,2個組合的F2葉片數的遺傳率分別有31.93 %和28.04 %的變異是由環境因素決定的,因此對葉片數的選擇也宜在早代。
3 結論與討論
西葫蘆良好的株型性狀是實現設施栽培高產、早熟和優質理想品種的必要條件。西葫蘆株型育種的目的是選擇株型緊湊的品種,本研究運用多世代聯合分析方法,對西葫蘆株型性狀進行了遺傳分析,并為西葫蘆育種提供了依據。
許多學者采用主基因-多基因混合遺傳分析方法對蔬菜的株高及葉片數進行了研究。辣椒和普通白菜(小白菜)的株高分別符合1對加性主基因+加性-顯性多基因模型與1對負向完全顯性主基因+加性-顯性多基因遺傳模型(陳學軍和陳勁楓,2006;韓建明 等,2008);而黃瓜株高受1對加性-顯性主基因+加性-顯性多基因共同控制,以主基因加性效應為主(嵇怡 等,2009);劉二艷等(2009)對青花菜花球莢葉性狀進行了遺傳分析,結果表明不同的群體同一性狀的遺傳模型不同。可見不同蔬菜株高和葉片數的遺傳模型并不一致,但其株高和葉片數均以主基因控制為主;而且不同植物或同一種植物不同基因型株高性狀的遺傳模型也不一致,可見材料的不同基因型對同一性狀的遺傳有著重要的影響;此外,環境對株高和葉片數有不同的影響。
本研究明確了西葫蘆株高和葉片數性狀受加性-顯性-上位性兩對主基因(B-1)遺傳模型控制,株高以基因的加性效應為主,葉片數以加性效應和顯性效應為主;F2的主基因遺傳率較高,環境影響相對較小,表明西葫蘆株型育種宜早代選擇。就豐產而言,株型緊湊的西葫蘆品種更高產(陳鳳真,2008)。因此,對西葫蘆株型性狀的遺傳改良,可選擇植株矮小、葉片數少、節間距短的親本材料,通過雜交、回交轉移主基因,選育株型緊湊的材料。
陳鳳真,何啟偉,樊治成,盛金.2007.西葫蘆8個農藝性狀的遺傳效應分析.園藝學報,34(5):1183-1188.
陳鳳真.2008.西葫蘆農藝性狀的遺傳、遺傳圖譜的構建及遺傳多樣性的研究〔博士論文〕.泰安:山東農業大學.
陳學軍,陳勁楓.2006.辣椒株高遺傳分析.西北植物學報,26(7):1342-1345.
蓋鈞鎰.2005.植物數量性狀遺傳體系的分離分析方法研究.遺傳,27(1):130-136.
韓建明,侯喜林,史公軍,陳沁濱.2008.不結球白菜株高性狀主基因+多基因遺傳分析.南京農業大學學報,31(1):23-26.
嵇怡,徐強,陳學好.2009.黃瓜株高性狀遺傳模型分析.揚州大學學報:農業與生命科學版,30(3):75-79.
李建友.2004.西葫蘆主要性狀雜種優勢及其形成機理的研究〔碩士論文〕.泰安:山東農業大學.
劉二艷,劉玉梅,方智遠,楊麗梅,莊木,張揚勇,袁素霞,孫繼峰,李占省,孫培田.2009.青花菜花球‘莢葉’性狀主基因+多基因遺傳分析.園藝學報,36(11):1611-1618.
Kasrawi MA.1995.Diversity in landraces of summer squash froMJordan.Genetic Resources and Crop Evolution,42:223-230.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
