低溫弱光對苗期紫花苜蓿根頸生理特性的影響
2010-07-25 08:52:08張彩峽呼天明
草業科學 2010年5期
關鍵詞:植物
張彩峽,呼天明
(1.西北農林科技大學動物科技學院,陜西楊凌 712100;2.西藏職業技術學院,西藏拉薩 850030)
①根頸是指根部上端膨大處,是聯系地上部和根系的關鍵部位,根頸上能夠形成大量的芽,是產生分枝的重要部位[1],同時也是吸收、運輸、儲存養分和水分的重要器官,直接影響植物的再生性、耐寒性、抗旱性和抗病蟲害性等生產性能和植物的可持續利用[2-3]。苜蓿Medicago sativa根頸為冷凍害的最敏感部位[4],對苜蓿越冬和春季返青時的萌芽發枝至關重要。目前,國內外有關苜蓿根系的研究主要集中于根系形態發育、根蘗特性與環境的關系等方面[5],有關苜蓿根頸的研究較少[6],只有少量研究涉及根頸病害及防治[5]。試驗采用人工控制溫度及光照條件的方法,分析了低溫弱光脅迫對不同秋眠級紫花苜蓿根頸的影響,以期為紫花苜蓿的品種選育及栽培提供支持,為苜蓿種質資源的鑒定、利用提供理論參考。
1 試驗材料與方法
1.1 試驗材料試驗選用的2個紫花苜蓿品種(WL414和巨人201)的種子由楊凌金道種子公司提供,秋眠級分別為6和2,發芽率90%以上。
1.2 處理方法將供試紫花苜蓿種子先用98%濃硫酸處理 1 h,再用5%次氯酸鈉溶液消毒5 min,最后用去離子水清洗5遍。將100粒種子置于鋪2層濾紙的培養皿(直徑10 cm)中,于培養箱(ZPQ350型)內20℃恒溫連續暗培養3 d。試驗期間根據種子吸脹情況適當補加蒸餾水。
試驗用苗于2009年6月1日-11月10日在西北農林科技大學全自動PVC溫室(光照培養16 h,暗培養8 h,恒溫25℃)營養盆培養。營養盆為圓錐形,內徑10 cm,高 25 cm,盆底有小孔,營養盆內裝細沙(0.8 kg/盆),澆透水后將發芽的幼苗從培養皿移栽于營養盆中(1株/盆),每隔1 d澆1次自來水,每周澆2次Hoagland營養液以促進其生長。植株到分枝期(形成大量分枝)時,將完整植株移至人工氣候培養箱(ZPQ350型)內,進行不同低溫弱光處理,處理方式見表1。
處理72 h后,采取根頸樣品,測定各生理指標。每處理中各參試材料分別為5株,每項測定設3次重復。
1.3 測定項目與方法參照高俊鳳主編的《植物生理學實驗指導》的方法[7]測定超氧化物歧化酶(SOD)、過氧化物酶(POD)活性和丙二醛(MDA)含量。
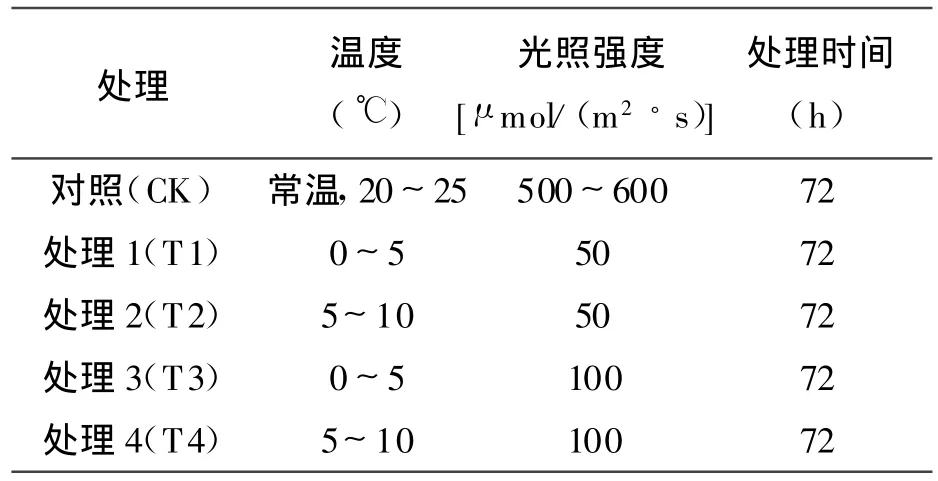
表1 處理方式
1.4 統計分析運用SPSS13.0數據進行統計分析,Microsoft Office Excel 2003進行數據運算及圖表處理。
2 結果與分析
2.1 低溫弱光對SOD活性的影響SOD作為植物體內保護酶之一,能清除過多積累的超氧陰離子自由基,避免或減輕逆境傷害。低溫弱光處理72 h后,WL414和巨人201根頸中的SOD活性顯著降低(圖1)。72 h后,WL414根頸的SOD活性各處理分別比 CK下降 57.24%、41.06%、54.12%和 38.63%,巨人201根頸中的SOD活性各處理分別比 CK下降 29.65%、15.01%、27.47%和13.52%。相同弱光條件下,低溫0~5℃處理SOD活性下降幅度顯著大于5~10℃處理(P<0.05)。相同低溫條件下,弱光50 μ mol/(m2·s)處理SOD活性下降幅度顯著大于 100 μ mol/(m2·s)處理(P<0.05),即2個品種的T1和T3處理的SOD活性下降幅度均大于T2和T4,T1的下降幅度最大,T4的下降幅度最小,且 WL414的SOD活性下降程度大于巨人201。在相關研究中也得到一致結論,秋眠級高的紫花苜蓿在低溫脅迫下SOD活性下降程度大于秋眠級低的[8]。
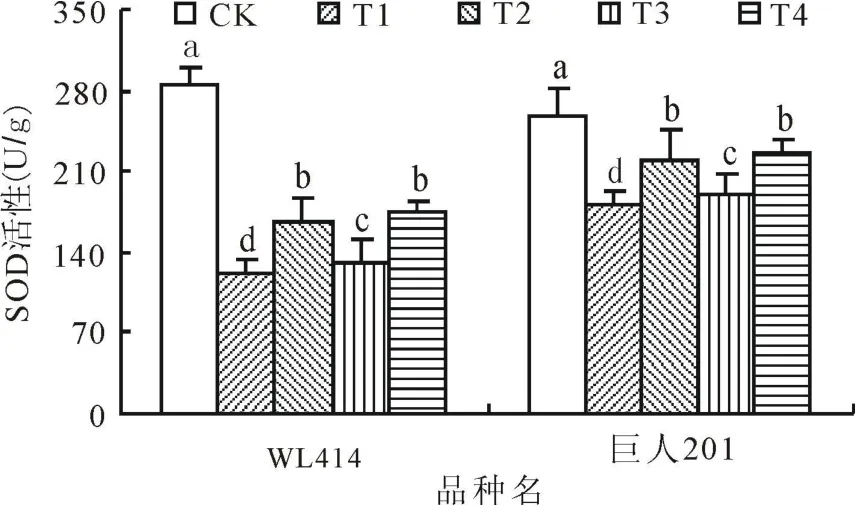
圖1 低溫弱光脅迫對紫花苜蓿根頸SOD活性的影響
2.2 低溫弱光對POD活性的影響POD是植物體內普遍存在的、活性較高的一種保護酶,是分解H2O2的主要保護酶,其活性的高低及增強幅度的大小直接關系到植物細胞的損傷程度。低溫弱光處理72 h后,WL414和巨人201根頸中POD活性呈顯著上升趨勢(圖2)。相同弱光條件下,低溫5~10℃處理POD活性增加幅度顯著大于0~5℃處理(P<0.05)。相同低溫條件下,弱光50和100 μ mol/(m2·s)處理組間 WL414差異不顯著,而巨人201的POD活性增加幅度弱光50 μ mol/(m2·s)處理顯著大 于 100 μ mol/(m2·s)處理(P<0.05)。WL414根頸的POD活性增加幅度小于巨人201,WL414根頸的POD活性各處理分別比CK增加13.30%、20.20%、10.20%和 18.70%,而巨人201根頸中POD活性各處理則分別比CK增加41.97%、50.43%、35.83%和44.27%,T2和 T4處理的POD活性增加幅度均大于T1和T3,其中T2的增加幅度最大,T3的增加幅度最小。從整個變化過程來看,巨人201保持著較高的POD活性,具有較高的H2O2清除能力,抵御低溫弱光傷害的能力較強。
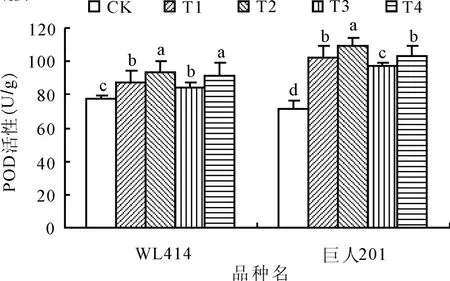
圖2 低溫弱光脅迫對紫花苜蓿根頸POD活性的影響
2.3低溫弱光對MDA含量的影響MDA是膜脂過氧化產物,是反映細胞膜傷害程度的重要指標之一。低溫弱光處理72 h后,WL414和巨人201根頸中MDA含量顯著上升(圖3)。相同弱光條件下,低溫0~5℃處理MDA含量增加幅度顯著大于5~10℃處理(P<0.05)。相同低溫條件下,弱光 50 μ mol/(m2·s)處理MDA 含量增加幅度顯著大于 100 μ mol/(m2·s)處理(P<0.05),WL414的MDA含量增加幅度大于巨人201。WL414根頸的MDA含量各處理分別比 CK 增加 79.53%、33.57%、62.90%和29.97%,而巨人201根頸中的MDA含量各處理則分別比CK增加21.44%、5.57%、14.99%和1.92%。T1和T3的MDA含量增加幅度均大于T2和T4,其中 T1的增加幅度最大,T4的增加幅度最小。與CK相比,低溫弱光脅迫均導致紫花苜蓿根頸中MDA含量的上升,這說明經低溫弱光脅迫,紫花苜蓿根頸細胞膜遭到不同程度的破壞[9],MDA大量積累。
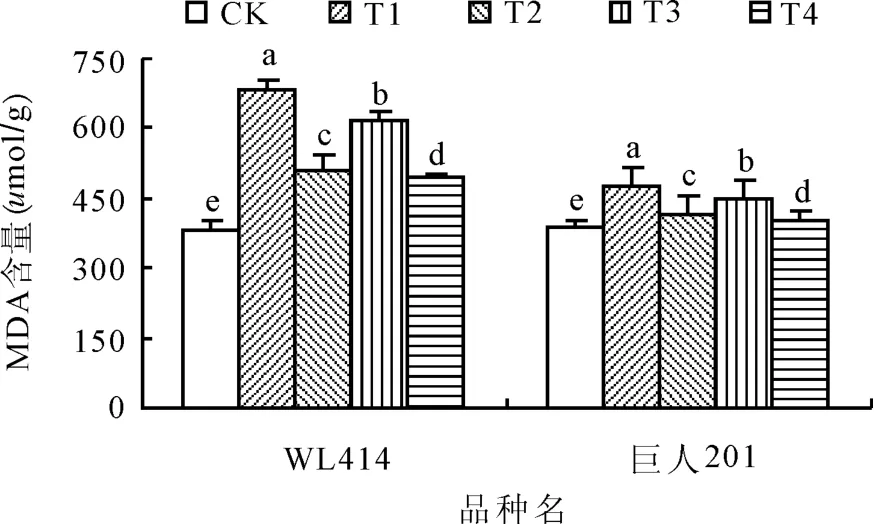
圖3 低溫弱光脅迫對紫花苜蓿根頸MDA含量的影響
3 討論與結論
植物體內清除活性氧的酶類主要包括SOD、過氧化氫酶(CAT)和POD等,這些酶能減輕或消除逆境對植物造成的傷害,清除自由基并防止自由基毒害,它們活性的變化可作為植物的耐寒指標[10]。當植物遭遇逆境脅迫時,往往保護酶活性越高其忍耐逆境的能力越強。近年來的研究表明,逆境下植物體內活性氧超過正常水平時,加劇了膜脂過氧化而導致膜系統受損,最終使組織結構遭到破壞。SOD的主要功能是催化超氧物陰離子自由基(O2-)發生歧化反應,生成 O2和H2O2。H2O2可以產生更多的羥基自由基,影響植物生長與光合生理特性,而POD可以清除體內的H2O2,維持體內的活性氧代謝平衡,保護膜結構,從而使植物能在一定程度上忍耐、減緩或抵抗逆境的脅迫。MDA作為膜系統磷脂不飽和脂肪酸的過氧化產物對細胞膜起傷害作用[10]。
本試驗中,低溫弱光處理72 h后,WL414和巨人201根頸中的POD活性上升,但SOD活性下降,MDA含量增大,表明為避免遭受低溫弱光逆境的傷害根頸作出了適應性反應,而根頸活性氧積累過多,超出了SOD和POD等膜保護酶的清除能力,部分未能清除的活性氧引起膜脂過氧化(MDA含量的增加顯著),導致膜的完整性被破壞。就相同弱光條件而言,低溫0~5℃對植株的影響比5~10℃更為顯著,POD活性上升幅度較小,而SOD活性的下降、MDA含量的增加幅度較大,這與蔡仕珍等[11]在花葉細辛Asarum gephilum的MDA變化研究中的結果一致,隨著濕度的降低MDA含量升高,使膜系統傷害更為嚴重。相同低溫條件下,弱光 50 μ mol/(m2·s)處理使POD活性上升、SOD活性的下降、MDA含量的增加幅度比 100 μ mol/(m2·s)更大。從品種間比較可知,巨人201對低溫弱光脅迫的適應能力強,能維持較高水平的SOD和POD活性[12-13],避免或減輕膜脂過氧化作用,細胞組織受的傷害較小[14]。結果表明,SOD和 POD活性、MDA含量可較好地反映植物抗逆能力[15]。
植物耐低溫弱光的機制相當復雜,僅從某一側面或層次去研究植物的耐低溫弱光能力是遠遠不夠的。植物耐低溫弱光的表現是多方面的,紫花苜蓿根頸的耐低溫弱光性與成株階段及開花結實階段的耐低溫弱光性是否一致,仍有待于進一步研究。
[1]耿華珠,吳永敷,曹致中,等.中國苜蓿[M].北京:中國農業出版社,1995.
[2]Marquez-Ortiz J J,Johnson L D,Basigalup D H,et al.Crown morphology relationships among alfalfa plant introduction and cultivars[J].Crop Science,1996,36:766-770.
[3]王月勝,于林清,張利軍.播種當年苜蓿根系研究初報[J].草地學報,2008,16(3):313-315.
[4]張寶田,穆春生,李志堅,等.紫花苜蓿受凍害后促進根頸枝條再生方法的研究[J].中國草地,2003,25(5):48-52.
[5]孟嫣,李敏權.苜蓿根和根頸腐爛病病原及防治研究進展[J].草業科學,2005,22(5):89-92.
[6]劉玉華,賈志寬.苜蓿秋眠性的研究進展[J].陜西農業科學,2002(7):20-22.
[7]高俊鳳.植物生理學實驗指導[M].北京:高等教育出版社,2006.
[8]史瑩華,王成章,張偉毅,等.苜蓿秋眠型與其超氧化物歧化酶和過氧化物酶關系的研究[J].草業科學,2009,26(8):128-131.
[9]施征.三個棗品種抗寒生理特性的研究[D].河北:河北農業大學,2003.
[10]崔國文.紫花苜蓿田間越冬期抗寒生理研究[J].草地學報,2009(2):145-150.
[11]蔡仕珍,潘遠智,陳其兵,等.低溫脅迫對花葉細辛生理生化及生長的影響[J].草業學報,2010,19(1):95-102.
[12]寇建村,楊文權,賈志寬,等.不同苜蓿品種抗寒性研究[J].中國草地學報,2008(4):11-15.
[13]魏臻武,王德賢,賀連昌.超氧化物歧化酶在苜蓿抗寒鍛煉過程中的作用[J].草業科學,2006,23(7):15-18.
[14]盧欣石,申玉龍.苜蓿秋眠性的研究與應用[J].國外畜牧業——草原與牧草,1992(1):1-4.
[15]何云.苜蓿秋眠性研究進展[J].草業科學,2005,22(11):25-29.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
