小菜蛾GluCl受體α亞基cDNA基因克隆和序列分析
2010-06-12 02:43:54梁延坡吳青君劉長仲張友軍王少麗徐寶云
植物保護 2010年4期
梁延坡, 吳青君, 劉長仲, 張友軍, 王少麗, 徐寶云
(1.甘肅農業大學草業學院,蘭州 730070; 2.中國農業科學院蔬菜花卉研究所,北京 100081)
谷氨酸是脊椎動物和無脊椎動物神經系統內重要的神經遞質,在脊椎動物體內主要通過門控陽離子通道介導興奮性的神經傳遞,而在無脊椎動物體內谷氨酸既是興奮性的神經遞質[1-3]又是抑制性的神經遞質[4]。作為抑制性的神經傳遞介質,谷氨酸與突觸后受體結合后,開啟氯離子通道,稱為谷氨酸門控的氯離子通道(glutamate-gated chloride channel,GluCl)[5]。GluCl屬于半胱氨酸環門控離子通道基因超家族,目前僅在無脊椎動物的神經和肌肉細胞中發現[4],在脊椎動物中尚未發現,因此是較為理想的殺蟲劑作用靶標。作用于GluCl的殺蟲劑包括阿維菌素/美倍霉素類、苯基吡唑類殺蟲劑氟蟲腈以及吲哚二萜類化合物nodulisporic acid等[6-8]。人們對于GluCl受體的分子結構和功能的了解大多數來自于對線蟲和模式昆蟲的研究[5],而對農業昆蟲的研究較少。
小菜蛾[Plutellaxylostella(L.)]屬鱗翅目菜蛾科,是世界性的十字花科蔬菜重要害蟲,對多種殺蟲劑都產生了抗藥性[9]。近年來對特效生物制劑阿維菌素類也產生了不同程度抗性[10]。早期的研究認為,阿維菌素作用于γ-氨基丁酸(GABA)受體達到殺蟲效果[11]。但是后來的研究發現,在較低濃度時,阿維菌素可引起與GABA系統無關的Cl-通道的開放,即谷氨酸控制的Cl-的開放,從而導致膜對Cl-通透性增加,帶負電荷的Cl-引起神經元休止電位的超極化,使正常的動作電位不能釋放,神經傳導受阻,最終引起蟲體麻痹死亡[12]。而且研究發現,線蟲對伊維菌素(阿維菌素的衍生物)的主要抗性機理是GluCl受體α亞基(GluClα)的堿基突變導致與藥物的親和力下降[13]。可見GluCl受體是阿維菌素類殺蟲劑的更為主要的作用靶標。目前小菜蛾對阿維菌素的抗性機理尚未明確,小菜蛾GluCl受體國內外也未見研究報道。本研究利用反轉錄-多聚酶鏈式反應(reverse transcription-polymerase chain reaction,RT-PCR)的方法首先對小菜蛾的GluCl受體α亞基cDNA片斷進行克隆和序列分析,然后采用RACE技術獲取小菜蛾GluCl受體α亞基的全長序列,為深入研究小菜蛾GluCl受體的分子特性和揭示阿維菌素類殺蟲劑的抗性機理奠定基礎。
1 材料與方法
1.1 供試昆蟲
供試小菜蛾為于室內用甘藍苗連續飼養多年的敏感種群,室內飼養溫度為(25±2)℃,RH為50%~70%,光周期L∥D=16h∥8h。
1.2 主要試劑和儀器
Trizol Reagent RNA kit和cDNA synthesis kit(SuperScript III)購 自 Invitrogen (USA)公 司;dNTP Mix、DNA Marker,100bp DNA Ladder均購自天根生化科技(北京)有限公司;TaqDNA聚合酶購自Promega公司;克隆載體pEASY-T1vector、Mach1-T1感受態細胞:購自全式金公司;PCR產物回收與純化試劑盒購自北京百泰克公司;TaKaRa LA Taq?:TaKaRa寶生物工程(大連)有限公司,TaKaRa 3′-Full RACE coreset試劑盒:購自 TaKa-Ra公司;SMARTTMRACE cDNA Amplification Kit:Clontech公司產品,PCR引物由上海生工合成;測序由北京諾賽基因組研究中心有限公司完成;PCR擴增儀為PTC-200,MJR。
1.3 試驗方法
1.3.1 總 RNA的提取
參照Trizol Reagent RNA提取試劑盒說明書進行總RNA抽提。
1.3.2 cDNA第1鏈合成
按Invitrogen公司的cDNA kit說明書進行。cDNA合成步驟參照Invitrogen in SuperScript III First-strand synthesis systeMfor RT-PCR 使 用 程序進行。
1.3.3 GluCl受體α基因cDNA片段的克隆
根據GenBank中已收錄的果蠅(Drosophila melanogaster)和秀麗隱桿線蟲(Caenorhabditiselegans)GluCl受體α亞基氨基酸序列的保守性區域,設計簡并性上下游引物(表1),進行同源克隆。
以cDNA為模板進行PCR擴增,50μL反應體系中含:第1鏈cDNA產物2μL,5μL 10×TaqDNA Buffer,2mmol/L dNTP,2mmol/L MgCl2,2μL dNTP(10mmol/L),2μL(10μmol/L)DEGF/DEG-R和0.5μL(5U/μL)TaqDNA polymerase。PCR反應條件為:94℃預變性3min,94℃變性1min,50℃退火1min,72℃延伸1min,循環35次,72℃延伸10min。
1.3.4 GluCl受體α亞基全長基因的克隆
1.3.4.1 GluCl受體α亞基3′-末端cDNA的克隆
根據 TaKaRa 3′-Full RACE coreset Kit說明書合成3′端cDNA,并根據片段測序結果,設計了3條特異性上游引物(表1),用于小菜蛾GluClα3′端cDNA序列的擴增,以3′GSP1和3′RACE Outer Primer為第1輪擴增引物,以3′GSP2或3′GSP3和3′RACE Inner Primer為第2輪擴增引物。
第1輪PCR在50μL反應體系中3′cDNA 4μL,1×cDNA Dilution BufferⅡ (Mg2+Plus)6μL,3′GSP1(10μmol/L)2μL,3′Outer Primer(20μmol/L)2 μL,TaKaRa LATaq(5U/μL)0.25μL,10×LA PCR BufferⅡ(Mg2+Plus)4μL,去離子水31.75μL;擴增程序為:95℃3min,94℃30s,55 ℃ 30s,72 ℃ 1.5min,35個循環;72℃10min。將第1輪PCR產物稀釋50倍作Nest-PCR 模 板,以 (3′NGSP2/3′RACE Innerprimer)和(3′NGSP3/3′RACE Inner-primer)為引物,進行第2輪PCR,擴增程序為:95℃3min;94℃30s,58 ℃ 30s,72 ℃ 1.5min,35個循環;72℃10min。
1.3.4.2 GluCl受體α亞基5′-末端cDNA的克隆
參照SMARTTMRACE cDNA Amplification Kit(Clontech)使用說明進行5′cDNA的合成,根據片段和3′RACE結果設計5′RACE特異性引物(表1)用于獲取5′端cDNA序列。
以5′GSP1和UPM為第1輪引物,94℃30s,72℃3min,5個循環;94℃30s,70℃ 30s,72 ℃3min,5 個 循 環;94 ℃ 30s,68 ℃ 30s,72 ℃3min,30個循環;72℃10min。將第1輪PCR產物稀釋50倍用5μL做Nest-PCR模板,以5′NGSP1和NUP為第2輪引物進行PCR擴增,擴增程序為:94℃30s,68℃30s,72℃3min,30個循環;72℃10min。

表1 試驗中設計的引物序列
1.3.5 cDNA 全長的驗證
根據擴增得到的5′和3′端片段的序列分別設計擴增全長cDNA的引物 QC-F/QC-R(表1)以驗證基因的完整性。擴增程序為:95℃3min,94℃30s,55 ℃ 30s,72 ℃ 2min,35 個 循 環;72 ℃10min。
1.3.6 PCR產物的克隆與測序
將PCR產物經DNA純化回收試劑盒回收純化后,克隆于PEASY-T1載體中,挑取陽性克隆經酶切和PCR鑒定后,送北京諾賽基因組研究中心有限公司進行測序。
1.3.7 在線序列比對和分析程序
蛋白質序列分析采用ExPASy在線ScanProsite程序;蛋白質分子量和等電點分析采用http∥:www.expasy.org/tools/pi-tool.html在線工具;序列比對使用ClustalW程序;序列和數據搜索使用NCBI上的Blast在線程序;信號肽序列預測采用http:∥www.cbs.dtu.dk/services/SignalP 在線程序進行;進化分析采用 MEGA4.0軟件[14]。
2 結果與分析
2.1 小菜蛾GluCl受體α亞基cDNA全長的克隆及序列分析
通過簡并性上下游引物進行RT-PCR擴增,結果見圖1a,擴增所得的片段大小約為192bp,與預期的目的擴增片段大小相符。將PCR產物克隆測序,所得cDNA片段總長度為192bp,與已知昆蟲的GluCl受體α亞基氨基酸序列進行相似性分析,與果蠅和飛蝗(Locustamigratoria)的氨基酸序列相似性為80%,表明克隆的cDNA片段是小菜蛾GluClα亞基基因的部分序列。根據已獲得的基因序列,分別設計5′-和3′-特異性引物,通過 RACE策略,成功的獲得含有5′-及3′-非編碼區的序列,其中3′端長度為1 014bp(圖1b),測序結果分析后發現,在這個片段包含一個TGA終止子以及多個TAA,在終止子的下游包含一個長603bp的非編碼區(UTR)。末端具有真核生物mRNA典型特征:一個含15個腺嘌呤的poly(A),在前面出現了加尾信號 AATAAA;5′-端序列長為1 090bp(圖1c),在這個片段中包含一個起始密碼子(ATG),在起始密碼子前有197bp的非編碼區(圖2)。序列分析結果可以證實得到了小菜蛾GluClαcDNA的完整序列。
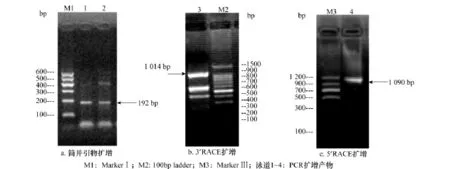
圖1 PCR擴增電泳圖譜
將5′-和3′-端序列拼接,得到了基因的全序列。為了進一步驗證拼接序列的正確性,通過已獲得的序列設計了基因編碼區的特異性引物以擴增全長cDNA片段。PCR擴增獲得了與預期大小相符的條帶,克隆結果同拼接序列比較,氨基酸序列并無差異,進一步驗證了獲得的基因序列的準確性和完整性。
2.2 序列分析
GluClα亞基cDNA全長2 144bp,用DNAMAN軟件進行分析,其開放閱讀框(ORF)長為1 344bp,編碼447個氨基酸殘基,根據在線軟件SignalP 3.0(http:∥www.cbs.dtu.dk/servi-ces/SignalP/)分析GluClα亞基氨基酸序列信號肽剪切位點,其信號肽最有可能的切割位點在第24位與第25位之間,預測出GluClα亞基蛋白信號肽序列含25個氨基酸,其序列為:MDVLRPSCALFVLFLLYCAHLTECV(圖2);蛋白質切割信號肽后經過簡單的加工過程而成為成熟肽;預測其蛋白分子量為51348.7u,其蛋白序列的理論等電點為8.46;相似性分析表明:它與埃及伊蚊的相似性為75%,與果蠅的相似性為73%,與赤擬谷盜的相似性為73%,與東亞飛蝗的相似性為79%。用在線軟件TMHMM2.0預測跨膜結構其包含有4個跨膜結構域,TM1-4(圖2)具有 GluClα亞基的典型特征:有一個長的N-末端半胱氨酸的胞外區和4個跨膜區和C-端胞外區,在其親水的胞外區由2對十分保守的半胱氨酸二硫鍵形成氨基酸殘基的橋環,其中第1對包含13個氨基酸殘基,第2個包含10個氨基酸殘基(圖2),這些特征表明該基因編碼受體α亞基。


圖2 編碼小菜蛾的GluClα亞基核酸序列及推導的氨基酸序列
2.3 進化分析
在GenBank中搜索果蠅、赤擬谷盜(TriboliuMcastaneum)、蝗蟲、蜜蜂(Apismellifera)和秀麗隱桿線蟲、犬心絲線蟲(Cylicocyclusnassatus)、腫孔古柏線蟲(Cooperiaoncophora)等物種的GluCl基因的α、β亞基的氨基酸序列,應用MEGA4.0軟件通過鄰位法(neighbor-joining)進行系統進化分析,以1 000次回抽的置信度作置信分析,結果如圖3所示,分別標注在分支節點上,每個分支末端依次為不同物種的GluCl的蛋白基因登錄號和物種名。系統進化分析表明:GluCl基因分為Ⅰ、Ⅱ兩個組,分別代表GluClα基因和GluClβ基因,Ⅰ組又可分兩亞族:昆蟲組Ⅰa和線蟲組Ⅰb;小菜蛾的GluCl屬于Ⅰa,昆蟲α亞基;由此證明了克隆的基因為GluCl受體的α亞基。
3 討論
谷氨酸是一種重要的神經遞質,在無脊椎動物中GluCl作為抑制性配體起作用,在昆蟲和甲殼類動物中,谷氨酸被公認為是作用于H受體而導致其去極化的這一假說。目前,在一些線蟲和果蠅屬中,GluCl已被克隆,并在爪蟾卵母細胞中表達,通過已克隆的通道顯示,無脊椎動物的GluCl受體上具有阿維菌素類藥劑的作用位點[15]。相對于線蟲,人們對昆蟲GluCl受體研究較少,其通道特性與線蟲的亦有所不同。目前GluCl受體在其他昆蟲如非洲鈍緣蜱、扁虱、草地貪夜蛾、谷實夜蛾[16]、美洲大蠊[7]、家蠅[17]和東亞飛蝗[18]等體內也有發現,但目前對任何生物的GluCl亞基的構成都不明確。
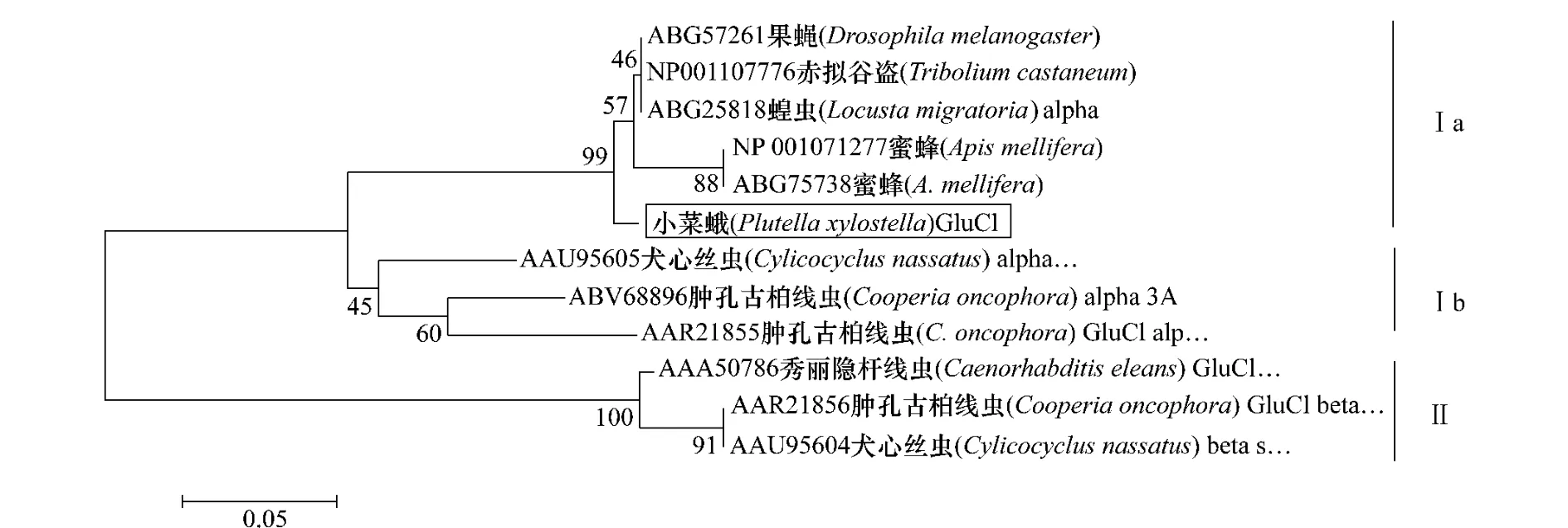
圖3 不同物種間GluCl蛋白的系統進化分析
靶標不敏感性是害蟲對殺蟲劑產生抗性極為重要的一種機制,尤其與高水平抗性有關。遺傳學研究表明,小菜蛾對阿維菌素的抗性是常染色體、不完全隱性遺傳,而且可能是有多基因控制的抗性遺傳[19]。吳青君[20]的研究認為,GABA 受 體 結合 數目的降低是小菜蛾對阿維菌素產生抗性的重要機制之一,放射性配體結合分析表明,敏感和抗性種群的受體親和力均無顯著差異,但抗性種群受體最大的結合數目比敏感種群降低了63.6%,說明是受體數目的減少,而不是結構上的改變導致了小菜蛾對阿維菌素的抗性。目前,還沒有GluCl受體不敏感性與昆蟲抗藥性關系的報道。本研究首次利用簡并引物通過RT-PCR和RACE技術成功地克隆了小菜蛾GluClα亞基的全長序列,根據這段序列可設計特異性引物進行抗性小菜蛾cDNA全長克隆和功能表達,比較其抗性和敏感種群的氨基酸差異,從分子水平明確小菜蛾GluCl功能亞基的分子性質,為研究小菜蛾對阿維菌素類殺蟲劑抗性靶標不敏感的分子機制奠定基礎。
[1]Gration K A,Clark R B,Usherwood P N.Three types of L-glutamate receptor on junctional membrane of locust muscle fibres[J].Brain Research,1979,171(2):360-364.
[2]Patlak J B,Gration K A,Usherwood P N.Single glutamateactivated channels in locust muscle[J].Nature,1979,278:643-645.
[3]Uitsch A,Schuster C M,Laube B,et al.Glutamate receptors ofDrosophilamelanogaster:cloning of a kainite-selective subunit expressed in the central nervous system[J].Proc Natl Acad Sci USA,1992,89(21):10484-10488.
[4]Cleland T A.Inhibitory glutamate receptor channels[J].Mol Neurobiol,1996,13:97-136.
[5]吳青君,張友軍,徐寶云.抑制性谷氨酸受體(IGluRs)通道及其相關殺蟲劑的作用[J].農藥學學報,2008,10(3):251-259.
[6]Bloomquist J R.Chloride channels as tools for developing selective insecticides[J].Arch Insect BiocheMPhysiol,2003,54:145-156.
[7]Zhao X L,Yeh J Z,Salgado V L,et al.Fipronil is a potent open channel blocker of glutamate-activated chloride channels in cockroach neurons[J].Journal of Pharmacology and Experimental Therapeutics,2004,314(1):363-373.
[8]Smith MM,Warren V A,Thomas B S,et al.Nodulisporic acid opens insect glutamate-gated chloride channels:identification of a new high affinity modulator[J].Biochemistry,2000,39(18):5543-5554.
[9]吳青君,張文吉,朱國仁.小菜蛾的發生為害特點及抗藥性現狀[J].中國蔬菜,2001(5):49-51.
[10]Syed T S,Abro G H,Ahmed S.Efficacy of different insecticides againstPlutellaxylostellaunder field conditions[J].Pakistan Journal of Biological Sciences,2004,7(1):10-13.
[11]Campbell W C.Ivermectin:an update[J].Parasitol Today,1985,1(1):10-16.
[12]Shoop W L,Mrozik H,Fisher MH.Structure and actvity of avermectins and milbemycins in animal health[J].Veterinry Parasitology,1995,59(2):139-156.
[13]Kane N S,Hirschberg B,Qian S,et al.Drug-resistantDrosophilaindicate glutamate-gated chloride channels are targets for the antiparasitics nodulisporic acid and ivermectin [J].PNAS,2000,97(25):13949-13954.
[14]Tamura K,Dudley J,Nei M,et al.MEGA4:molecular evolutionary genetics analysis(MEGA)software version 4.0[J].Mol Biol Evol,2007,24:1596-15991.
[15]Cully D F,Wilkinson H,Vassilatis D K,et al.Molecular biology and electrophysiology of glutamate-gated chloride channels of invertebrates[J].Parasitology,1996,113:S191-S200.
[16]Cully D F,Paress P F,Liu K K,et al.Identification of aDrosophilamelanogasterglutamate-gated channel sensitive to the antiparasitic agent avermectin[J].The American Society for Biochemistry and Molecular Biology,1996,271:20187-20191.
[17]Eguchi Y,Ihata M,Ochi E,et al.Functional characterization ofMuscaglutamate-and GABA-gated chloride channels expressed independently and coexpressed inXenopusoocytes[J].Insect Molecular Biology,2006,15(6):773-783.
[18]Janssen D,Derst C,Buckinx R,et al.Dorsal unpaired median neurons ofLocustamigratoriaexpress ivermectin and fipronilsensitive glutamate-gated chloride channels[J].Journal of Neurophysiology,2007,97:2642-2650.
[19]李騰武,高希武,鄭炳宗,等.小菜蛾對阿維菌素的抗性遺傳分析及交互抗性研究[J].植物保護,1999,25(6):12-14.
[20]吳青君,張文吉,張友軍,等.表皮穿透和GABAA受體不敏感性在小菜蛾對阿維菌素抗性中的作用[J].昆蟲學報,2002,45(3):336-340.
