三江源地區高寒草地群落特征研究
2010-05-30 03:52:40杜巖功崔驍勇葛勁松趙旭東王勇
草業科學 2010年3期
關鍵詞:物種
杜巖功,崔驍勇,葛勁松,趙旭東,任 杰,王勇
(1.中國科學院研究生院資源與環境學院,北京100049;2.青海省環境監測中心站,青海 西寧810007)
生物多樣性與全球變化是目前各國面臨的主要生態、環境問題[1]。生物多樣性研究主要集中于物種多樣性[2],它是群落生態組織水平的主要特征之一,也是群落生態環境優劣的重要指標[3],物種多樣性較高,系統受到外界干擾時有較大的反饋調節能力,生態系統穩定性高。
高寒草地是青藏高原的主體類型之一,近年來由于長期超載放牧,造成高寒草地嚴重退化、沙化,生物多樣性降低,草地產草力下降,草地生態環境日趨惡化[4]。因此,青藏高原高寒草地群落特征研究、高寒草地退化和恢復治理研究尤為迫切。目前有關江河源區高寒草地多樣性研究主要集中在荒漠化草地和紫花針茅草原群落特征[5-6]及高寒草地不同放牧強度下演替過程、特征。結果認為:高寒草甸草地生產力高,牧草品質優良,物種多樣性和豐富度指數較高;物種多樣性受海拔、緯度、溫度和濕度等因素影響[1];隨著放牧干擾強度增加,物種多樣性和豐富度指數逐漸降低,草地生物量降低[6-7]。
本研究在三江源地區選擇典型的高寒草原和高寒草甸樣地,通過對其生物多樣性和分布格局研究,深入分析該地區群落特征,為該地區生物多樣性保護和生態恢復提供科學依據。
1 材料與方法
1.1 試驗區自然概況 三江源地區平均海拔4 000 m以上,氣候寒冷屬高寒大陸性氣候,只有冷暖兩季,沒有四季之分,冷季持續7~8個月,全年無絕對無霜期。年降水量420~560 mm,多集中在5-10月,植物生長季雨熱同期。草地類型主要以針茅草原和高寒草甸為主,土壤類型主要為高山草甸土、高山草原土[7]。
優勢物種為高山嵩草Kobresia pygmaea、短花針茅 Stipabrevi f lora、紫花針茅S.purpurea、青海固沙草Orinus kokonorica,伴生種主要有二裂委陵菜Potentilla bi f urca、早熟禾Poa indattenuata、苔草Carex ivanovae、矮火絨草 Leontopodium nanum 、棘豆 Oxytropisochrocephala(見表1)。
1.2 試驗設計與方法 2008年8月在三江源區選擇針茅草原和高山嵩草草甸的6處典型樣地,進行植物群落調查、土壤濕度測定。地上生物量采用標準收獲法,群落調查樣方面積1 m×1 m;0~10 cm土壤濕度采用TDR(時域反射儀)測定,3次重復。α多樣性用豐富度指數、物種多樣性指數和均勻度指數3類多樣性指數表征[8]。
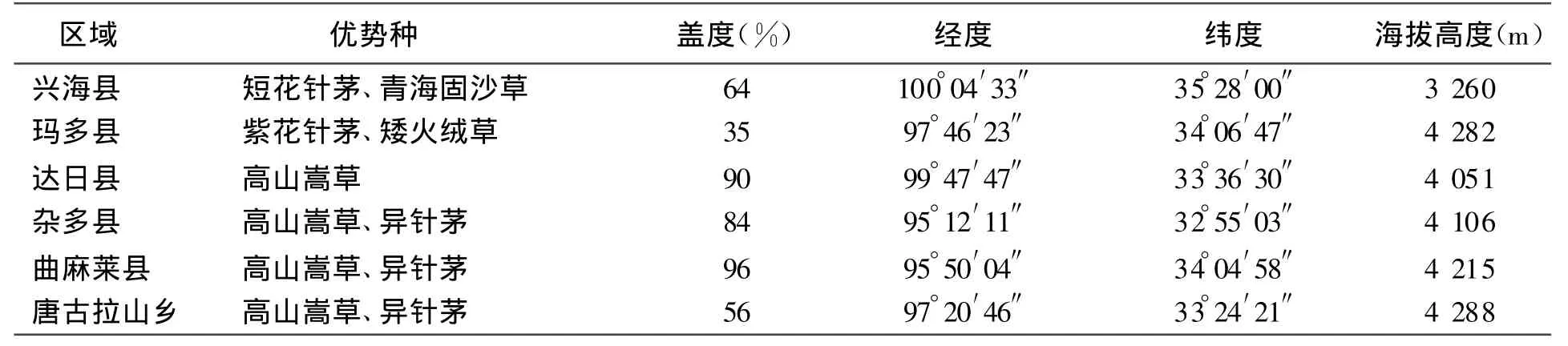
表1 樣地基本情況
豐富度指數R=S
物種多樣性指數計算采用 Shannon-Wiener指數:

式中,Pi為種i的相對重要值:Pi=(相對高度+相對蓋度)/2;S為種i所在樣方的物種總數。
1.3 數據分析 不同樣地土壤濕度、物種數差異采用單因素方差分析;物種多樣性和物種數采用回歸分析;群落特征進行聚類分析;可食用鮮草產量與土壤濕度、物種數目、物種多樣性和海拔采用相關性分析(SPSS 16.0)。
2 結果與分析
2.1 不同樣地土壤濕度 高寒草地的水分來源主要依靠降水[9],土壤濕度是影響植物生長的重要環境因子。高寒草甸土壤濕度顯著高于高寒草原土壤(興海縣、瑪多縣,P<0.05)。曲麻萊縣土壤濕度最高,達到42.82%,唐古拉山鄉最低,為21.27%(P<0.05),雜多縣和達日縣土壤濕度一致(P>0.05);興海縣短花針茅草原土壤濕度為12.63%,而瑪多縣紫花針茅草原土壤濕度僅為7.80%,二者之間達到顯著性差異水平(P<0.05,圖1)。這可能是因為高寒嵩草草甸致密的草皮層具有較大的水分保蓄能力[10]。賀金生等[11]認為從大尺度來看,水熱條件及其組合是決定植物群落分布的主要因素。
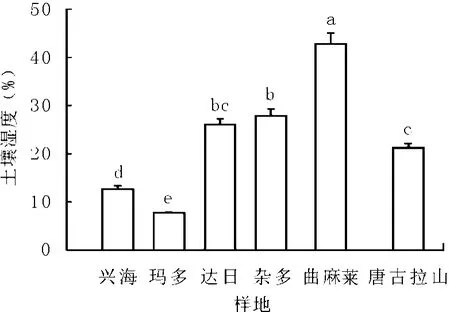
圖1 高寒草地土壤濕度比較
2.2 不同植物群落的α多樣性特征 物種豐富度是群落多樣性的最基本特征。達日縣和曲麻萊縣高寒草甸物種數較高,分別為20種和15種,二者之間達到顯著性差異水平(P<0.05);興海縣、瑪多縣和唐古拉山鄉草地物種數較少,均是8種,雜多縣物種數最低,僅有5種(P>0.05,圖2)。物種數與海拔和土壤濕度之間相關性不顯著(P>0.05)。雜多縣物種數最低,但其海拔不是最高,土壤濕度也不是最低,并且緯度最低,這說明物種豐富度可能是受溫度、降水、海拔和放牧利用強度等因素共同作用影響。岳鵬鵬等[5]2005年調查結果認為瑪多縣紫花針茅草原物種數為12~26,物種數高于本研究,這可能是由于不同年份和采樣地點受放牧干擾造成。李凱輝等[12]發現新疆天山高寒草原和草甸植物物種約有11~16種,這與本研究結果一致。
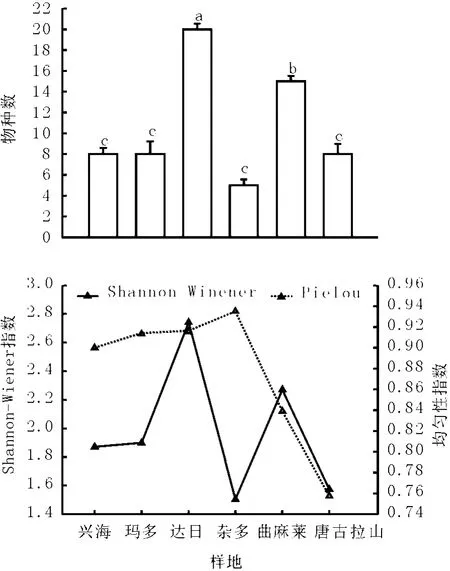
圖2 高寒草地物種豐富度和物種多樣性指數
物種多樣性是生物多樣性的核心內容之一,其在生物多樣性保護中具有重要意義[1]。達日縣和曲麻萊縣高寒草甸物種多樣性最高,分別為2.74和2.27;唐古拉山鄉和雜多縣最低,約為1.5;興海縣和瑪多縣針茅草原物種多樣性約為1.87。均勻度指數唐古拉山鄉和曲麻萊縣稍低,分別為0.76和0.84,其余各縣均勻度指數均較高(圖2)。楊元武等[1]在祁連山高寒草甸研究結果認為多樣性為0.65~3.37,而Wang等[8]在果洛州高寒草甸中發現物種多樣性為2.8~3.5;岳鵬鵬等[5]認為紫花針茅草原物種多樣性為2.2左右。
物種多樣性的緯度格局,一般表現為隨著緯度降低,物種多樣性增加[11]。在針茅草原和高寒嵩草草甸中這種規律都不明顯,其中雜多縣緯度最低,但物種多樣性也最低(圖2),岳鵬鵬等[5]認為在瑪多縣紫花針茅草原物種多樣性指數與緯度呈顯著負相關關系。Currie[13]認為在美國東部物種多樣性變化存在明顯的緯度梯度,而在西部這種特征表觀不明顯,這可能是由于地形、溫度差異等原因造成[11]。物種多樣性隨著海拔梯度變化規律很復雜[11],本研究發現物種多樣性與海拔之間無顯著相關性(P>0.05),隨海拔增加,物種多樣性變化規律不明顯。這與楊元武等[1]、楊立軍等[14]在祁連山和達日縣高寒草地研究結果一致,這表明在低溫、高海拔的地區高寒草地物種多樣性可能受多種因素影響。
三江源地區平均海拔在4 000 m以上,生態系統脆弱,地形、溫度和濕度差異會成為生物多樣性增加的主要限制因素。
6處樣地統計分析發現物種多樣性和物種數極顯著相關(r=0.967,P<0.01),這與 Wang等[8]和胡玉昆等[15]研究結果一致。在三江源地區高寒草地多樣性指數擬合方程為y=1.130 1+0.079 5x(R2=0.935 9,P <0.01,圖 3),用物種數可以較好地擬合物種多樣性指數。
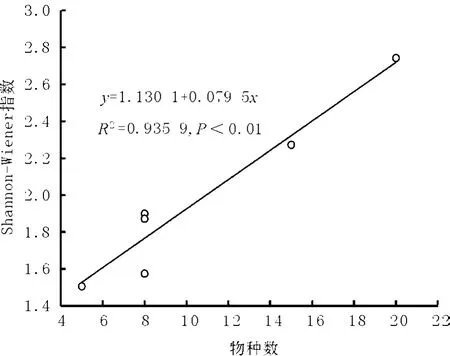
圖3 高寒草地Shannon-Wiener指數與物種數目擬合方程
2.3 高寒草地鮮草量和地下生物量比較可食用鮮草量是草地承載力的重要指標,高寒草甸鮮草生產力均高于針茅草原,其中達日縣高山嵩草草甸產草量最高,約為466.8 g/m2,其次是雜多縣、曲麻萊縣和唐古拉山鄉高寒草甸,瑪多縣紫花針茅草原鮮草產量較低,約為71.0 g/m2(圖4)。統計結果發現可食用鮮草量與土壤濕度、海拔、物種數沒有顯著相關性(P>0.05)。總鮮草量和地下生物量與可食用鮮草量表現出一致規律。達日縣高寒草甸地下生物量最高,為3 916.7 g/m2,且鮮草產量最低的瑪多針茅草原地下生物量也達到733.3 g/m2。這可以看出高寒草地植物根系發達,生物量高,這與杜巖功等[10]在海北州高寒草甸研究結果一致。杜利霞等[16]發現放牧會增加冷蒿Atemisia f rigida地下部分生物量而減少地上部分生物量,這可以使其回避被采食。董全民等[17]看到高山嵩草和早熟禾生態位較寬,而紫羊茅Festuca rubra和青海野青茅D.kokonorine生態位很窄。劉發央等[18]發現適度的放牧干擾抑制草地中優勢種的競爭作用,為其他物種提供潛在生態位。高寒草甸根系致密,并能發育出其特有的草氈表層,可耐放牧家畜的踐踏,是維持高寒草甸生態系統穩定性的重要生態屏障[10],這可能也是高寒草甸植物長期對環境適應的生態策略。
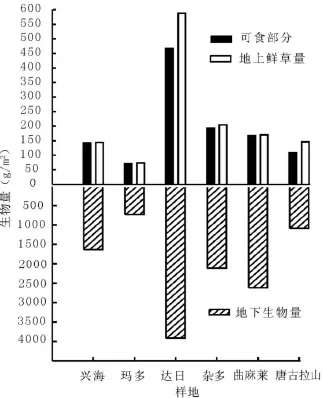
圖4 高寒草地不同樣地生物量
3 討論
不同地區高寒草地群落在蓋度、高度、植物種類組成、重要值和多樣性等方面都變現出一定的差異。這種分布特征,可能是因為對地帶性氣候、地理隔離等較敏感[5]。
聚類分析是將研究樣品或者變量之間相似性程度大的聚為一類,把另外一些相似性大的聚為另一類,然后繼續分別聚類,最終聚為一個大類,結果采用直觀的聚類樹狀分類圖表示。重要值是反映物種在群落中重要性的一個主要指標[4],根據樣方調查數據,按照群落物種重要值進行聚類分析發現,雜多縣和曲麻萊縣高山嵩草-異針茅草甸首先被歸為一類,這二者蓋度也基本一致;其次將唐古拉山鄉高山嵩草草甸合為一類;然后依次是興海縣短花針茅草原、達日縣高寒嵩草草甸;瑪多縣紫花針茅草原最后被聚合為一類,這可能是由于其物種數和蓋度均較低。聚類分析的結果和樣方調查數據中的優勢種和草地類型分類結果一致(表1,圖5)。岳鵬鵬等[5]對江河源區紫花針茅草原的不同地區樣地群落進行研究也發現聚類分析結果和草地類型分類相當一致,本研究也發現該地區紫花針茅草原分布呈現出小尺度同質性,大尺度異域異質性特征。孫海松[19]對青海省21個草地類型進行聚類分析發現21類草地可以分為12類,認為用這種定量方法分類結果合理。張靜等[20]采用聚類分析方法將三江源區12塊退化草地分為9類,看到草地之間有較大差異,治理上應采取不同措施。
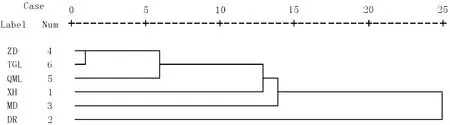
圖5 高寒草地群落聚類分析
人類活動干擾和過度放牧,以及氣候干暖化是造成草地退化的重要原因[21]。隨著主要以根莖繁殖的莎草科植物物種數減少,草皮層壞死,凍融過程和鼠害同時加速草皮層剝蝕,最終造成土層裸露,土壤隨雨水發生大量流失[20]。對處于輕度和中度干擾高寒草地,需要通過降低或者解除放牧壓力,以及補播、耙松等措施,使草地得到休養生息,保持草氈表層在一定限度,既可以使草地保持較高生產力又可以緩解草地的退化[10,22-23]。而對于已經出現草皮的塌陷和斑駁等重度、極度退化高寒草地,采用建植栽培草地,短期內可實現草地植被的恢復,張耀生等[24]通過連續13年試驗發現,建植栽培草地可以有效逆轉高寒草地退化趨勢。
4 主要結論
4.1 高寒草甸土壤濕度顯著高于高寒草原土壤(P<0.05),曲麻萊縣土壤濕度最高,唐古拉山鄉最低(P<0.05),興海縣短花針茅草原土壤濕度約為12.63%,而瑪多縣紫花針茅草原土壤濕度僅為7.80%,二者之間達到顯著差異(P<0.05)。
4.2 達日縣和曲麻萊縣高寒草甸物種數較高,分別為20種和15種,二者之間達到顯著性差異水平(P<0.05)。興海縣、瑪多縣和唐古拉山鄉草地物種數都是8種,雜多縣物種數最低僅有5種(P>0.05)。物種數與海拔和土壤濕度均沒有顯著相關性(P>0.05)。
4.3 達日縣和曲麻萊縣物種多樣性最高,分別為2.74和2.27;唐古拉山鄉和雜多縣最低,約為1.5;興海縣和瑪多縣針茅草原物種多樣性約為1.87。均勻性指數唐古拉山鄉和曲麻萊縣稍低,分別為0.76和0.84,其余各縣均勻性指數均較高。多樣性指數隨緯度和海拔變化規律不明顯,但與物種數目極顯著相關(r=0.967,P<0.01),擬合方程為 y=1.130 1+0.079 5x(R2=0.935 9,P<0.01)。
[1] 常學向,趙文智,趙愛芬.祁連山區不同海拔草地群落的物種多樣性[J].應用生態學報,2004,15:1599-1603.
[2] Tracy C,Brussard P.Preserving biodiversity:Species in landscapes[J].Ecology Application,1994(4):205-207.
[3] 劉振國,李鎮清.退化草原冷蒿群落13年不同放牧強度后的植物多樣性[J].生態學報,2006,26:475-480.
[4] 杜巖功,曹廣民,王啟蘭,等.放牧對高寒草甸地表特征和土壤物理性狀的影響[J].山地學報,2007,25:338-343.
[5] 岳鵬鵬,盧學峰,葉潤蓉,等.江河源不同區域紫花針茅草原群落特征[J].植物生態學報,2008,32:1116-1125.
[6] 楊元武,李希來,祁盛倉.江河源地區不同荒漠化草地物種多樣性研究[J].青海大學學報(自然科學版),2005,23:42-45.
[7] 張靜,李希來,于海.青藏高原不同退化程度小嵩草草甸群落結構特征與土壤理化特征分析[J].草原與草坪,2008(4):5-9.
[8] Wang C,Long R,Wang Q,et al.Effects of altitude on plant-species diversity and productivity in an alpine meadow,Qinghai-Tibetan plateau[J].Australian Journal of Botany,2007,55:110-117.
[9] 曹廣民,李英年,鮑新奎.高寒地區寒凍雛形土的持水特性[J].土壤,1998(1):27-31.
[10] 杜巖功,梁東營,曹廣民,等.放牧強度對嵩草草甸草氈表層及草地營養和水分利用的影響[J].草業學報,2008,17(3):146-150.
[11] 賀金生,陳偉烈.陸地植物群落物種多樣性的梯度變化特征[J].生態學報,1997,17:91-99.
[12] 李凱輝,胡玉昆,王鑫,等.不同海拔梯度高寒草地地上生物量與環境因子關系[J].應用生態學報,2007,18:2019-2024.
[13] Currie D,Paquin V.Large-scale biogeographical pattern of species richness of trees[J].Nature,1987,329:326-327.
[14] 楊力軍,李希來,石德軍,等.青南高海拔地區高寒草甸植物群落多樣性的研究[J].草原與草坪,2000(2):32-35.
[15] 胡玉昆,李凱輝,柳妍妍,等.天山南坡高寒草地海拔梯度上的植物多樣性變化格局[J].生態學雜志,2007,26:182-186.
[16] 杜利霞,李青豐,董寬虎.放牧強度對短花針茅草原冷蒿繁殖特性的影響[J].草地學報,2007,15:367-370.
[17] 董全民,趙新全,馬玉壽,等.放牧對高寒小嵩草草甸冷季草場主要植物種群生態位的影響[J].中國草地學報,2006,28:10-17.
[18] 劉發央,徐長林,龍瑞軍.牦牛放牧強度對金露梅灌叢草地群落物種多樣性的影響[J].草地學報,2008,16(6):613-618.
[19] 孫海松.應用聚類分析法對青海21個草地型分類的初步探討[J].青海大學學報(自然科學版),2003,21(6):7-10.
[20] 張靜,李希來,謝得雄.三江源地區不同退化草地聚類分析[J].草業科學,2008,25(6):8-13.
[21] 王建兵,王振國,呂虹.黃河重要水源補給區草地退化的氣候背景分析[J].草業科學,2008,25(4):23-27.
[22] 曹廣民,龍瑞軍.三江源區黑土灘型退化草地自然恢復的瓶頸及解決途徑[J].草地學報,2009,17(1):4-9.
[23] 崔慶虎,蔣志剛,劉季科,等.青藏高原草地退化原因述評[J].草業科學,2007,24(5):20-26.
[24] 張耀生,趙新全,黃德清.青藏高寒牧區多年生人工草地持續利用的研究[J].草業學報,2003,12(3):22-27.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
學生天地(2019年36期)2019-08-25 08:59:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
