乙腦病毒prME蛋白與鼠GM-CSF編碼基因的融合構建與表達
2010-05-25 01:43:28翟永貞周言馬力馮國和
中國醫科大學學報 2010年9期
翟永貞,周言,馬力,馮國和
(中國醫科大學 附屬盛京醫院感染科,沈陽 110004)
流行性乙型腦炎 (Japanese encephalitis,JE)DNA疫苗的免疫效果已經得到實驗證實,編碼流行性乙型腦炎病毒(Japanese encephalitis virus,JEV)各種結構蛋白或非結構蛋白基因重組子能夠提供不同程度的保護性免疫,免于JEV病毒攻擊[1];通過比較上述重組質粒在分別免疫動物后所產生的中和抗體效價與保護性免疫效果,發現將前膜(prM)蛋白與包膜(E)蛋白編碼基因聯合構建于同一表達載體所產生的免疫效果明顯優于單純含E蛋白編碼基因重組子[2]。然而同現有JE滅活或減毒活疫苗免疫效果比較,DNA疫苗誘導的中和抗體產生時間、抗體效價增長速度[3]、效價絕對值還達不到預期標準[4]。JEDNA疫苗增強效應的研究已越來越受到人們的重視。
粒細胞-巨噬細胞集落刺激因子(granulocytemacrophage colony-stimulating factor,GM-CSF) 編碼基因已經被應運于JE DNA疫苗增強效應的研究,研究證實GM-CSF編碼基因重組子能夠增加抗JEV總抗體以及中和抗體效價,提高免疫動物生存率[5]。目前國內外尚未研究報道有關JEDNA疫苗與GM-CSF編碼基因的融合構建與表達。本項研究構建JEV prME蛋白與BALB/c鼠GM-CSF編碼基因真核重組表達載體,以脂質體法轉染中華倉鼠卵巢(Chinese hamster ovary,CHO)細胞并建立融合蛋白穩定表達株,為JEDNA疫苗增強效應的深入研究提供實驗依據。
1 材料與方法方法部分要刪減
1.1 質粒、細胞及主要試劑
含JEV prME蛋白編碼基因重組質粒被命名為pJME,由感染科實驗室構建[6],所選真核表達載體為pcDNA3,在BamH I與EcoR I酶切位點間插有JEV prME蛋白編碼基因。原核表達載體pMD19-T simple[Code No.D104C]購自Takara公司,用于BALB/c鼠GM-CSF編碼基因測序。真核表達載體pcDNA3.1(+)購自 Invitrogen公司,用于 JEV prME蛋白與GM-CSF編碼基因的融合構建。CHO細胞購自中國科學研究院上海細胞庫,生長于37℃,5%CO2及含 10%胎牛血清(fatal calf serum,FCS)的 Dulbecos modified eagle medium(D-MEM)中,脂質體(Lipofectamine2000)購自Invitrogen公司,分別用于轉染實驗中受體細胞及轉染試劑。大腸桿菌JM109與DH5α,限制性內切酶EcoRI、BamH I以及Not I等均購自Takara公司,用于重組質粒構建、轉化及酶切鑒定。新霉素類似物(G418)購自GiBCO公司,用于CHO細胞抗性克隆株的篩選。山羊抗鼠GMCSF購自Santa公司,辣根酶標記兔抗山羊IgG購自北京中山公司,電致化學發光(electrogenerated chemiluminescence,ECL) 檢測試劑購自 Pierce公司,用于重組質粒轉染后Western-blot分析。
1.2 BALB/c鼠GM-CSF編碼基因的重組構建
從BALB/c鼠脾臟獲取細胞總RNA,采用套式-RT-PCR法擴增GM-CSFCDS區域序列(426 bp)。反轉 錄 引 物 :5′-CAGGCACAAAAGCAGCAGTC-3′;PCR外引物:上游引物為5′-CAGAGAGAAAGGCTAAGGTC-3′,下游引物為上述反轉錄引物;PCR內引物:上游引物為 5′-GAATTCGGTGGAGGCGGTTCAGGTGGAGGTGGTTCAGGAGGAGGTGGATC GATGTGGCTGCAGAATTTAC-3′,下游引物為 5′-GCGGCCGCTCATTTTTGGCCTGGTTTTTTG-3′,用于PCR內引物的上游引物和下游引物的5’端分別引入EcoRI和Not I酶切位點,劃線部分為由15個氨基酸組成的Gly-Linker。使用DNA片斷純化試劑盒(Takara,product No.DV807) 與 DNA A 尾試劑盒(Takara,product No.D404)分別對擴增的DNA片段進行純化及“A”處理,使用瓊脂糖凝膠DNA純化試劑盒(Takara,product No.DV805)進行切膠回收DNA,按照DNA連接試劑盒(Takara,product No.D6023)說明書要求,后者與pMD19-Tsimple連接構建重組子pMD-GM-CSF,轉化大腸埃希菌JM109。質粒小劑量提取試劑盒(Takara,product No.DV801A)提取重組質粒并經EcoRI/Not I進行酶切鑒定,采用ABIPRSMTM310Genetic Analyzer(Pekin-Elmer/Applied)及引物Bca BESTPrimer M13-47對重組質粒進行測序分析。將測序正確的目的基因經EcoR I/Not I雙酶切并亞克隆到真核表達載體pcDNA3.1(+),將此重組子命名為pGM-CSF。
1.3 JEV prME蛋白與BALB/c鼠GM-CSF編碼基因的融合構建
使用限制性內切酶BamH I/EcoR I對質粒pGM-CSF、pJME進行雙酶切,將獲取的JEV prME蛋白編碼基因亞克隆至pGM-CSF的BamH I/EcoRI酶切位點之間,含有JEV prME蛋白與BALB/c鼠GM-CSF編碼基因的重組子被命名為pJME/GMCSF。轉化pJME/GM-CSF于大腸桿菌DH5α并經篩選后,分別經BamH I/EcoRI,BamH I/Not I酶切以及1%瓊脂糖凝膠電泳鑒定。
1.4 pJME/GM-CSF轉染及CHO細胞抗性克隆株篩選
確定本實驗最佳G418濃度篩選:將常規培養的CHO細胞(1 000個/mL)加入24孔板(0.7 mL/每孔)孵育,6 h后每孔分別加入不同濃度的G418,依次為 600 mg/L,650 mg/L,700 mg/L,750 mg/L,800 mg/L,850 mg/L,900 mg/L,每種濃度設 3 個孔,余下3孔為正常細胞對照。培養11d后確定能殺死所有細胞的最低G418濃度作為本實驗的最佳篩選濃度,即為800 mg/L。脂質體法轉染pJME/GM-CSF于CHO 細胞[6],24 h 后將轉染細胞按 1∶10傳代,48 h后更換為含800 mg/L G418的D-MEM進行篩選,7 d后將G418濃度降為400 mg/L維持培養。
1.5 Western-blot分析
參照參考文獻[7]并加以改進,收集pJME/GMCSF轉染的CHO細胞,用細胞裂解液破碎細胞后收集總蛋白,12%SDS-PAGE凝膠電泳完畢后電轉移至PVDF膜上,用含5%脫脂奶粉的TBST封閉1 h,加入山羊抗鼠GM-CSF多克隆抗體(1∶200稀釋),4℃過夜,TBST洗滌10 min×3次,加入HRP標記的兔抗山羊的二抗(1∶50 000稀釋),室溫2 h,TBST洗滌10 min×3次,加入ECL試劑,顯影、定影。方法部分內容太細,常規方法寫明名稱就可,不用詳細介紹步驟。盡量節省版面。
2 結果
2.1 pJME/GM-CSF的重組構建
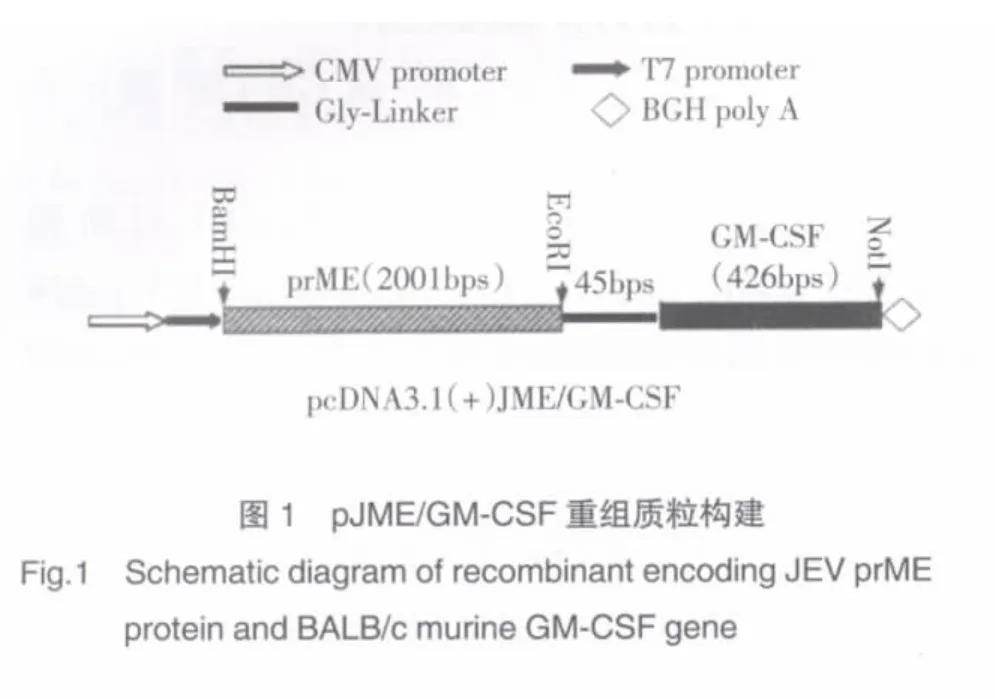
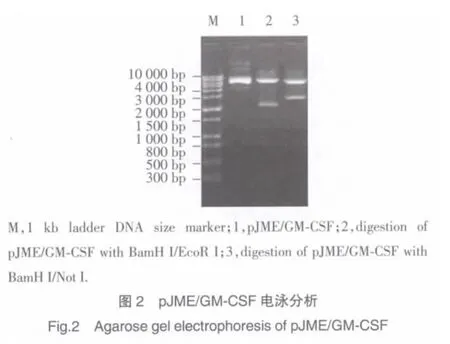
套式-RT-PCR法獲得的GM-CSF編碼基因經測序分析與發表的GM-CSFCDS區域序列相符合。將處理后的GM-CSF基因插入真核表達載體pcDNA3.1(+)EcoR I/Not I酶切位點,構建重組子pGM-CSF(圖1)。pJME中JEV prME蛋白編碼基因插入pGMCSF BamH I/EcoR I酶切位點,構建重組子pJME/GM-CSF(圖 1,圖 2 lane1)。pJME/GM-CSF 經 BamH I/EcoR I酶切釋出插入子與prME蛋白編碼基因片段(2 001 bp)大小相符(圖 2 lane2),pJME/GM-CSF經BamH I/Not I酶切釋出的插入子片段約為2 472 bp(圖 2 lane3),為鼠 GM-CSF與 JEV prME蛋白編碼基因大小之和。
2.2 G418篩選過程中pJME/GM-CSF轉染的CHO細胞抗性克隆株的演變
G418最適篩選濃度為800 mg/L。將轉染pJME/GM-CSF的CHO細胞加壓篩選(800 mg/L G418)7 d,培養板中可見小細胞團簇形成,G418濃度減半(400 mg/L)維持培養26 d,普通光鏡檢查顯示小細胞團簇逐步增大,細胞團簇之間逐漸融合(圖3)。

2.3 pJME/GM-CSF轉染的CHO細胞的Wester blot分析
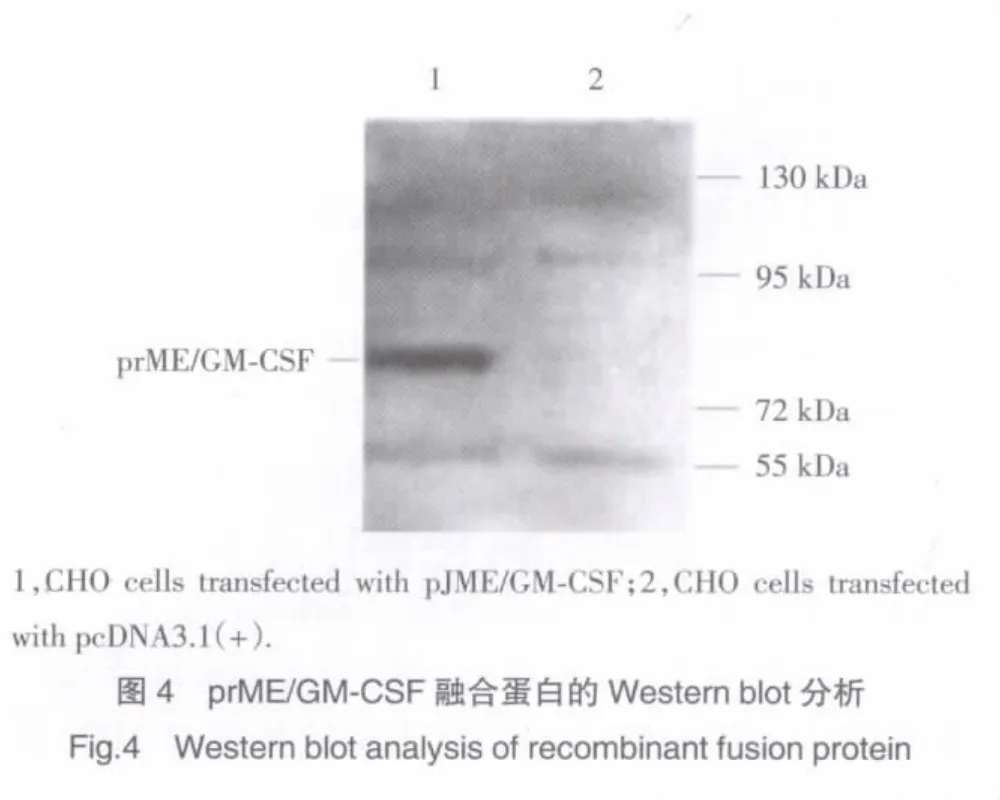
將pJME/GM-CSF轉染的CHO細胞裂解并收集蛋白進行Western blot分析,在Mr為85×103區域內可見一特異性蛋白帶,空載體轉染的CHO細胞未見相應蛋白帶(圖4)。
3 討論
許多研究表明,細胞因子基因佐劑對DNA疫苗所致的免疫應答有著重要的調節作用[8]。GM-CSF具有刺激樹突狀細胞分化與成熟、增強樹突狀細胞表面MHC以及共刺激分子表達等免疫促進功能,其編碼基因被作為DNA免疫佐劑并用于許多感染性疾病和腫瘤性疾病DNA疫苗增強效應的研究[9]。通常采用三種方式將GM-CSF編碼基因作為免疫佐劑用于DNA疫苗增強效應的研究,一種是將GMCSF編碼基因與目的抗原基因構建于雙順反子真核表達載體;一種是分別將GM-CSF編碼基因和目的基因構建于不同載體并共同注射動物模型,通過這種組合方式免疫,能夠篩選出具有較強免疫效應的組合DNA疫苗;另一種是將GM-CSF編碼基因和目的基因融合表達于同一載體,編碼這種融合蛋白的DNA疫苗能夠改善抗原特異性免疫應答和腫瘤性疾病的免疫治療。
本實驗將JEV prME蛋白編碼基因與鼠GMCSF編碼基因(2 001 bp、426 bp)以定向克隆的方法分別插至真核表達載體pcDNA3.1(+),構建重組子pJME/GM-CSF,后者經BamH I/EcoRI酶切釋出的插入子與pJME經相同酶切釋出插入子的分子量大小相一致(圖2 lane2),從分子層面說明該重組子含有JEV prME蛋白編碼基因,又經BamH I/Not I酶切釋出的插入子分子量約為2 472 bp(圖2 lane3),理論推算符合JEV prME蛋白編碼基因與鼠GM-CSF編碼基因的總和標準。DNA測序分析進一步明確了兩個目的基因在真核表達載體中的正常連接,提示含JEV prME蛋白編碼基因與鼠GM-CSF編碼基因重組子pJME/GM-CSF構建成功。
將轉染pJME/GM-CSF的CHO細胞加壓篩選(800 mg/L G418)7 d后,繼續以400 mg/L的G418維持培養,可見小細胞團簇逐步增大,細胞團簇之間逐漸融合(圖3),說明pJME/GM-CSF轉染的CHO細胞抗性克隆株逐步形成。進一步Western blot分析發現,pJME/GM-CSF轉染的CHO細胞經裂解電泳后,在大于Mr約72×103以上區域內可檢測到一特異性蛋白帶,推算其Mr大小約85×103,進而明確了融合蛋白分子量,通過對融合蛋白氨基酸數目(824AA)的理論推算,所檢測的融合蛋白分子量與理論計算相符合。本實驗成功構建了JEV prME蛋白與鼠GM-CSF融合基因重組子,并建立CHO細胞穩定表達株,將為JEDNA疫苗增強效應等研究提供實驗依據。
[1]Putnak R,Porter K,Schmaljohn C.DNA vaccinesfor flaviviruses[J].Adv.Virus Res,2003,61:445-468.
[2]Wu HH,Chen CT,Lin YL,et al.Sub-fragments of the envelope gene arehighly protectiveagainst the Japaneseencephalitisviruslethal infection in DNA priming-protein boosting immunization strategies[J].Vaccine,2004,22(5-6):753-800.
[3]Zhao Z,Wakia T,Yasui K.Inoculation of plasmidsencoding Japanese encephalitis virus-prM-Epritains with colloidal gold elicits a protectiveimmuneresponsein BALB/c mice[J].JVirol,2003,77(7):4248-4260.
[4]Ashok MS,Rangarajan PN.Protective efficacy of a plasmid DNA encoding Japanese encephalitis virus envelope protein fused to tissue plasminogen activator signal sequences:studies in a murine intracerebral viruschallenge model[J].Vaccine,2002,20(11-12):1563-1570.
[5]Bharati K,Appaiahgari MB,Vrati S.Effect of Cytokine-encoding plasmid delivery on immune response to Japanese encephalitis virus DNAvaccineinmice[J].Microbiol Immunol,2005,49(4):349-353.
[6]馮國和,趙桂珍,竹上勉,等.乙腦病毒prME與E蛋白編碼基因重組構建的DNA免疫試驗研究[J].中華微生物學和免疫學雜志,2002,22(5):505-509.
[7]Feng GH,Liu N,Zhou Y,et al.Immunologic analysis induced by DNA vaccine encoding E protein of Beijing-1 strain derived from Japaneseencephalitisvirus[J].Intervirology,2007,50(2):93-98.
[8]翟永貞,馮國和.流行性乙型腦炎DNA疫苗增強效應研究進展[J].國際兒科學雜志,2007,34(5):319-322.
[9]Wang J,Snider DP,Hewlett BR,et al.Transgenic expression of granulocyte-macrophage colony-stimulating factor induces the differentiation and activation of a novel dendritic cell population in the lung[J].Blood,2000,95(7):2337-2345.
