3-氧十二烷酰高絲氨酸內(nèi)酯對淋巴細胞Toll樣受體2和4的影響*
2010-03-17 09:06:58余加林楊錫強
中國病理生理雜志 2010年5期
關(guān)鍵詞:劑量
蘆 起, 余加林, 楊錫強, 劉 偉
(重慶醫(yī)科大學附屬兒童醫(yī)院,重慶400015)
銅綠假單胞菌是醫(yī)院感染中最常見的機會致病菌。它常引起急性和慢性肺部感染,多見于局部或全身免疫功能障礙的患者。現(xiàn)代微生物學觀點認為銅綠假單胞菌(Pseudomonas aeruginosa,PA)感染大多以生物膜形式存在[1]。PA內(nèi)各個細胞間的交流有賴于群體感應(yīng)系統(tǒng)(quorum-sensing,QS系統(tǒng))。QS系統(tǒng)不僅在調(diào)節(jié)銅綠假單胞菌生物膜形成上,而且在免疫調(diào)節(jié)上均有關(guān)鍵作用[2]。Toll樣受體(Toll-like receptor,TLR)是一組重要的免疫模式識別受體,TLR信號轉(zhuǎn)導在啟動機體的固有免疫反應(yīng)、介導獲得性免疫反應(yīng)中起到重要作用。本研究探討QS系統(tǒng)信號分子對外周血淋巴細胞增殖、TLR 2、TLR4表達,及腫瘤壞死因子α(tumor necrosis-α,TNF-α)分泌的影響。
材料和方法
1主要試劑
3-氧十二烷酰高絲氨酸內(nèi)酯(3-oxo-C12-h(huán)omoserine lactone,3-O-C12-HSL)購自Cayman公司,R/MINI-1640培養(yǎng)基和小牛血清購自杭州四季青生物制品公司。MTT和 DMSO購自Sigma,淋巴細胞分離液購自天津灝洋生物制品科技有限公司。RNA提取試劑盒購自北京Biotake公司,逆轉(zhuǎn)錄-聚合酶鏈反應(yīng)(reverse transcription polymerase chain reaction,RT-PCR)試劑盒和DNA marker購自大連寶生物工程有限公司,PCR引物由上海生工生物工程公司合成,CD3單克隆抗體、人TNF-α試劑盒購自深圳晶美生物工程有限公司。
2 外周血淋巴細胞的分離和分組
取健康獻血者外周靜脈血40 mL,肝素抗凝,以淋巴細胞分離液分離出外周血單個核細胞(peripheral blood mononuclear cells,PBMCs),PBS充分洗滌,用含10%小牛血清RPMI-1640培養(yǎng)液調(diào)整細胞數(shù)為1×1010cells/L,采用12孔塑料培養(yǎng)板,隨機分成5組。分別加入0、1、10、50、100 μmol/L 3-O-C12-HSL各10 μL,在37℃、5%CO2培養(yǎng)箱中培養(yǎng)12 h。吸取上層淋巴液。經(jīng)CD3單克隆熒光抗體染色后用流式細胞儀檢測,證實所得的細胞85%以上為CD3+淋巴細胞。
3 TLR 2和4 mRNA的表達檢測
按Trizo1說明書抽提總RNA。以oligod T為引物將提取的總RNA反轉(zhuǎn)錄,然后采用25 μL反應(yīng)體系,運用PCR方法對TLR2和4及內(nèi)參照β-actin同時進行PCR擴增。引物序列:TLR2:正義鏈5'-ggacttctcccatttccgtct-3,反義鏈5'-ctccaggtaggtcttggtgttc-3,產(chǎn)物138 bp;TLR4:正義鏈5'-ctgtccctgaaccctatgaact-3,反義鏈5'-cttctaaaccagccagaccttg-3’,產(chǎn)物135 bp;β-actin正義鏈5'-ccacgaaactaccttcaactcc-3,反義鏈5'-gtgatctccttctgcatcctgt-3',產(chǎn)物131 bp。擴增產(chǎn)物采用2%瓊脂糖凝膠電泳分離(電泳條件:100 V,20 min),溴化乙啶(5 mg/L)紫外燈下照相,用圖像分析系統(tǒng)分析計算吸光度值(A),表達的相對量用目的基因/β-actin A值表示。
4 MTT法測定
淋巴細胞懸液置于96孔微量細胞培養(yǎng)板中,每孔加入7×l09cells/L細胞100 μL,隨機分成5組。同時分別加入0、1、10、50、100 μmol/L 3-O-C12-HSL 1 μL,各設(shè)6個重復孔,另設(shè)培養(yǎng)液空白對照孔,在37℃、5%CO2培養(yǎng)箱中繼續(xù)培養(yǎng)12 h,加入20 μL MTT(5 g/L)。37℃繼續(xù)培養(yǎng)6 h,加入150 μL DMSO低速振蕩10 min。用酶聯(lián)儀測定波長490 nm A值。
5 ELISA法檢測TNF-α
將加入0、1、10、50、100 μmol/L 3-O-C12-HSL加入1×1010cells/L外周血單個核細胞,培養(yǎng)12 h后,收集培養(yǎng)上清,14 000 r/min離心10 min,保存于-80℃冰箱中。用人TNF-α試劑盒按其說明書進行操作。
6 統(tǒng)計學處理
結(jié)果
1 不同劑量3-O-C12-HSL對CD3+淋巴細胞TLR2/4 mRNA的表達
未處理組TLR2/4 mRNA表達最低,隨著3-O-C12-HSL劑量的加大,單核細胞TLR 2/4 mRNA逐漸增高,僅TLR4 mRNA未處理組和1 μmol/L比較P>0.05,余組間比較P<0.05,差異顯著,見圖1、表1。
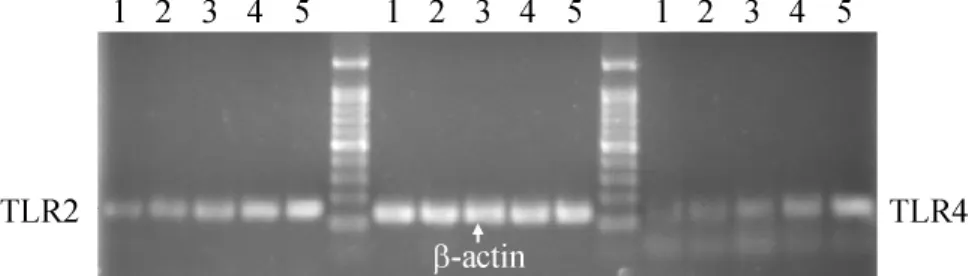
Figure 1.TLR2/4 mRNA expression in lymphocyte cells stimulated with 3-O-C12-HSL at different concentrations for 12 h.Lane 1:3-O-C12-HSL 0 μmol/L; Lane 2:1 μmol/L;Lane 3:10 μmol/L;Lane 4:50 μmol/L;Lane 5:100 μmol/L.圖1 不同濃度3-O-C12-HSL刺激后CD3+淋巴細胞TLR2/4 mRNA的表達
表1 不同濃度3-O-C12-HSL刺激后淋巴細胞TLR2/4 mRNATable 1 .Comparison of TLR2/4 mRNA expression in lymphocyte cells stimulated with 3-O-C12-HSL at different concentrations for 12 h(±s.n=3)
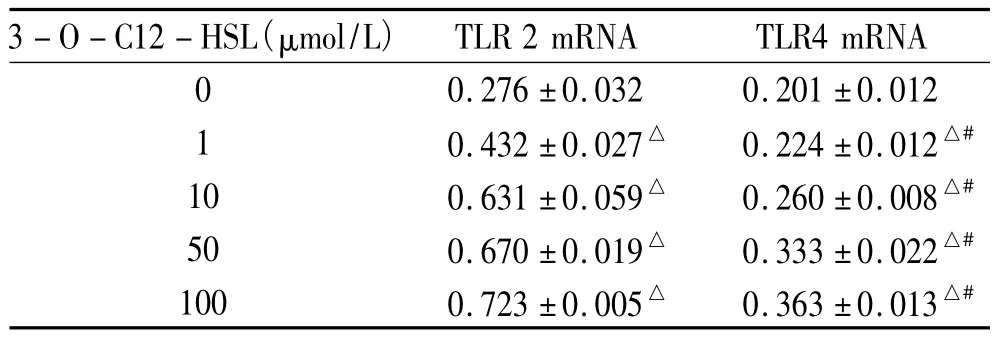
表1 不同濃度3-O-C12-HSL刺激后淋巴細胞TLR2/4 mRNATable 1 .Comparison of TLR2/4 mRNA expression in lymphocyte cells stimulated with 3-O-C12-HSL at different concentrations for 12 h(±s.n=3)
△P<0.05 vs 0 μmol/L 3-O-C12-HSL;#P<0.05 TLR2 mRNA at same dose of 3-O-C12-HSL.
?
2 不同劑量3-O-C12-HSL對CD3+淋巴細胞活力的影響
淋巴細胞的活力與未處理組比較增高,P<0.05,差異顯著。100 μmol/L 3-O-C12-HSL促進細胞增殖的作用最明顯,見表2。
表2 不同濃度3-O-C12-HSL刺激淋巴細胞MTT比色結(jié)果Table 2 .Comparison of MTT results in 3-O-C12-HSL-stimulated lymphocytes(±s.n=4)

表2 不同濃度3-O-C12-HSL刺激淋巴細胞MTT比色結(jié)果Table 2 .Comparison of MTT results in 3-O-C12-HSL-stimulated lymphocytes(±s.n=4)
△P<0.05 vs 0 μmol/L 3-O-C12-HSL.
?
3 不同劑量3-O-C12-HSL對TNF-α分泌的影響
不同劑量3-O-C12-HSL作用于外周血單個核細胞12 h后,在0 μmol/L、1 μmol/L、10 μmol/L、50 μmol/L、100 μmol/L 3-O-C12-HSL濃度組,單個核細胞TNF-α分別為719.968±5.732、593.256 ±25.040、490.649 ±9.654、367.137±15.386、267.090±13.878。與0 μmol/L組相比,其余各劑量組均可抑制外周血單個核細胞上清中的TNF-α的表達(P<0.05),差異顯著。100 μmol/L組3-O-C12-HSL抑制TNF-α分泌作用最明顯,見圖2。
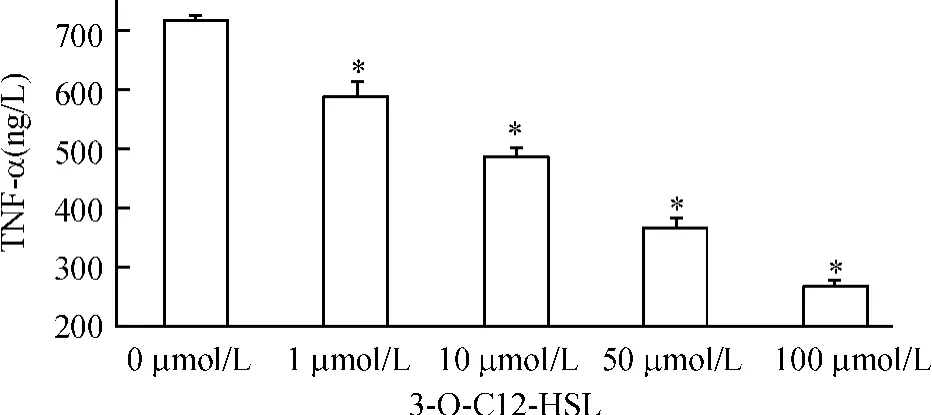
Figure 2.Effect of 3-O-C12-HSL on production of TNF-α in mononuclear cells.±s.n=3.*P<0.05 vs 3-O-C12-HSL.圖2 不同濃度3-O-C12-HSL抑制單個核細胞TNF-α的分泌
討論
PA主要產(chǎn)生2種QS系統(tǒng)信號分子:3-氧十二烷酰高絲氨酸內(nèi)酯[N-(3-oxododecanoyl)-L-h(huán)omoserine lactone,3-O-C12-HSL]和N-丁酰基高絲氨酸內(nèi)酯[N-butyryl-L-h(huán)omoserine lactone,C4-HSL]。QS系統(tǒng)信號分子具有調(diào)節(jié)宿主免疫反應(yīng)的作用。研究發(fā)現(xiàn):lasI基因或lasI rhlI雙基因缺失的銅綠假單胞菌不能在C57BL/6鼠肺部形成感染模型[3]。QS系統(tǒng)缺陷銅綠假單胞菌與相對應(yīng)的野生型比較誘導HCE細胞產(chǎn)生IL-8能力降低[4]。3-O-C12-HSL能夠誘導巨噬細胞的細胞毒性導致凋亡[7]。將3-OC12-HSL直接注射到小鼠皮膚里,其可通過NF-κB通路導致某些細胞因子的分泌,檢測發(fā)現(xiàn)皮膚組織中的IL-1、IL-6、MIP-2、MIP-1 mRNAs表達增高[5]。
TLRs是一組與固有免疫密切相關(guān)的受體家族[6]。可介導固有免疫應(yīng)答的產(chǎn)生和細胞因子的分泌[7]。TLR 2、TLR4在非特異性免疫中可廣泛識別配體,在識別危險信號并誘發(fā)機體免疫反應(yīng)中具有重要作用。感染后細胞TLR2和TLR4表達上調(diào),并可誘導合成一系列炎性介質(zhì),是機體免疫系統(tǒng)對感染的一種防御反應(yīng),清除外來病原菌。目前對于抗原呈遞細胞上TLRs研究較透徹,而淋巴細胞上的TLRs研究較少,多集中在調(diào)節(jié)性T細胞上。研究表明,TLR2可以引起調(diào)節(jié)性T細胞(T regulate cell,Treg)增殖,但同時Treg抑制活性發(fā)生了一過性消除;隨著TLR2配體的清除,Treg抑制活性可以完全恢復。離體實驗顯示,細菌產(chǎn)物可以直接作用于Treg的TLR4,增強Treg的抑制活性[8,9]。
本研究結(jié)果表明,不同濃度的3-O-C12-HSL刺激單個核細胞后,淋巴細胞TLR 2、TLR4表達幾乎均上調(diào),TLR2亞家族的上調(diào)較TLR4為明顯,提示TLR 2、TLR4上調(diào)不僅僅是增殖的結(jié)果,3-O-C12-HSL可影響淋巴細胞TLR 2、TLR4的不同表達,其原因可能為TLR 2、TLR4對3-O-C12-HSL的刺激有著明顯的差異,TLR 2對3-O-C12-HSL更易感。其次RT-PCR檢測存在一定誤差,可通過實時定量PCR進一步證實。此外,3-O-C12-HSL能誘導淋巴細胞的增殖,與Hooi等[10]報道的QS系統(tǒng)可以通過抑制T細胞的增殖,擾亂免疫調(diào)節(jié)活動的結(jié)果不一致,這可能與Hooi等[10]直接用3-O-C12-HSL刺激淋巴細胞,而本研究是用3-O-C12-HSL刺激單個核細胞后,然后收集淋巴細胞有關(guān),其原因可能為3-O-C12-HSL刺激單核細胞后,其分泌的細胞因子影響淋巴細胞TLR 2、TLR4表達有關(guān)。本研究發(fā)現(xiàn)TNF-α表達是隨著3-O-C12-HSL增高而降低,而Kim等[11]發(fā)現(xiàn)CD4+CD25+Foxp3-或CD8+T細胞能夠抑制不同固有免疫細胞的細胞因子的釋放。因此,我們推測淋巴細胞的TLR 2、TLR4表達抑制了TNF-α表達,獲得性免疫對固有免疫有反饋作用。至于是哪種(CD4+或CD8+)淋巴細胞亞型的增高,以及該亞群淋巴細胞的增高,對細胞內(nèi)信號轉(zhuǎn)導的影響,尚不清楚,還需進一步深入研究和探討。
[1] Bauer TT,Torres A,F(xiàn)errer R,et al.Biofilm formation in endotracheal tubes.Association between pneumonia and the persistence of pathogens[J].Monaldi Arch Chest Dis,2002,57(1):84-87.
[2] Saleh AC,F(xiàn)igarella W,Kammouni S,et al.Pseudomonas aeruginosa quorum-sensing signal molecule N-(3-oxododecanoyl)-L-h(huán)omoserine lactone inhibits expression of P2Y receptors in cystic fibrosis tracheal gland cells[J]. Infect Immun,1999,67(10):5076-5082.
[3] Smith RS,Harris SG,Phipps R,et al.The Pseudomonas aeruginosa quorum-sensing molecule N-(3-oxododecanoyl)homoserine lactone contributes to virulence and induces inflammation in vivo[J].J Bacteriol,2002,184 (4):1132-1139.
[4] Zhu H,Conibear TC,Thuruthyil SJ,et al.Pseudomonas aeruginosa quorum-sensing signal molecules induce IL-8 production by human corneal epithelial cells[J].Eye Contact Lens,2008,34(3):179-181.
[5] Tateda K,Ishii Y,Horikawa M,et al.The Pseudomonas aeruginosa autoinducer N-3-oxododecanoyl homoserine lactone accelerates apoptosis in macrophages and neutrophils[J].Infect Immun,2003,71(10):5785-5793.
[6] 田 青,周 恒,王 蕾,等.Toll樣受體3在自身免疫性心肌炎小鼠心肌細胞中的表達及意義[J].中國病理生理雜志,2009,25(12):2323-2328.
[7] 劉 穎,鄭春泉.Toll樣受體和免疫性疾病[J].中國眼耳鼻喉科雜志,2008,8(1):58-60.
[8] Sutmuller RP,den Brok MH,Kramer M,et a1.Tolllike receptor 2 controls expansion and function of regulatory T cells[J].J Clin Invest,2006,116(2):485-494.
[9] Caramalh I,Lopes-carvalho T,Ostler D,et al.Regulatory T cells selectively express Toll-like receptors and activated by lipopolysaccharide[J].J Exp Med,2003,197 (4):403-411.
[10]Hooi DS,Bycroft BW,Chhabra SR,et al.Differential immune modulatory activity of Pseudomonas aeruginosa quorum-sensing signal molecules[J].Infect Immun,2004,72(11):6463-6470.
[11]Kim KD,Zhao J,Auh S,et al.CD4+CD25+Foxp3+Tregs resolve experimental lung injury in mice and are present in humans with acute lung injury[J]Nat Med,2007,13(10):1248-1252.
猜你喜歡
課堂內(nèi)外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
今日農(nóng)業(yè)(2022年4期)2022-11-16 19:42:02
鄉(xiāng)村科技(2021年33期)2021-03-16 02:26:54
國際放射醫(yī)學核醫(yī)學雜志(2021年10期)2021-02-28 08:41:58
藥學與臨床研究(2015年4期)2015-06-05 11:35:54
衛(wèi)生職業(yè)教育(2014年24期)2014-05-20 09:05:38
同位素(2014年2期)2014-04-16 04:57:20
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
