從放線菌發酵液中高通量篩選端粒酶抑制劑
2010-02-10 08:35:28張慧敏王衛國李波蘭沈會芳林壁潤龐建新吳曙光
中國藥理學通報 2010年1期
張慧敏,曹 瑩,王衛國,李波蘭,沈會芳,林壁潤,龐建新,吳曙光
(1.南方醫科大學藥學院新藥評價中心,廣東廣州 510515; 2.珠江醫院藥材科,廣東廣州 510280;3.廣東省農科院植物保護所,廣東廣州 510640)
以端粒酶為靶點篩選端粒酶抑制劑是近年來尋找抗腫瘤新藥的重要途徑之一[1]。端粒酶有活性的最小組成單位是端粒酶逆轉錄酶(human telomerase reverse transcriptase,hTERT)和起模板作用的端粒酶RNA(human telomerase RNA,hTR),體外只要重組這兩部分就能重構端粒酶活性[2]。hTR-hTERT結合是可逆的,容易受抑制劑的影響。因此,如果篩選模型是針對hTR-hTERT的結合,就能提高篩選的特異性。根據以上理論,本實驗采用了基于酵母三雜交原理[3]的端粒酶抑制劑篩選模型。該模型是將端粒酶主要成分即hTERT和hTR的易結合片斷導入酵母菌L40 ura3/pHybLex/Zeo-MS2,通過在96孔板缺陷培養基上培養酵母[4],根據酵母菌的生長狀態,高通量篩選抑制hTR與hTERT特異結合的抑制劑。
1 材料與方法
1.1 菌株 L40 ura3/pHybLex/Zeo-MS2由南華大學何淑雅教授惠贈;L40 ura3/pHybLex/Zeo-MS2/ hTR126-451/hTERT292-608aa、L40ura3/pHybLex/ Zeo-MS2/hTR240-450/hTERT292-608aa、L40ura3/ pHybLex/Zeo-MS2/hTR(m)/hTERT292-952aa[5]為本實驗室構建好的酵母三雜交系統。
1.2 試劑和儀器 2 000多株放線菌發酵液由廣東省農科院植保所提供;DNA聚合酶、dNTPS購自TaKaRa公司;YPD酵母培養基、YC-UHW酵母培養基[6]、PCR儀為美國AppLied Biosystems公司產品;多功能酶標儀Bench mark pLus,水平電泳槽及電泳儀購自為美國Bio-Rad公司;96孔細胞板購于美國Corning Costor公司。
1.3 酵母菌液PCR驗證篩選系統 參照GenBank hTR和hTERT序列,采用引物設計軟件設計5對引物如Tab 1所示,引物由英駿生物技術有限公司合成,用去離子水配制成20 μmol·L-1。挑取3種菌株的單克隆菌于YC-UHW中培養,然后用煮沸冷凍法[7]提取基因組。分別取以上3種上清1 μl于3個PCR管內,反應體系為50 μl,擴增hTR126-451、hTR240-450、hTRm,循環參數為95℃預變性5 min,95℃30 s,退火30 s(退火溫度依引物而定),72℃延伸30 s,30次循環,72℃延伸5 min。再分別取3種上清1μl于3個PCR管內,反應體系為50μl,擴增hTERT292-952、hTERT292-952,循環參數為95℃預變性5 min,95℃ 30 s,60℃退火1 min,72℃延伸2 min,30次循環,72℃延伸5 min。反應擴增產物瓊脂糖凝膠電泳鑒定。
1.4 放線菌發酵液樣品庫 2 000多株不同編號的放線菌樣品(經發酵的米飯)0.3 g置于5 ml貼有標簽的青霉素瓶內,用體積分數為0.5的乙醇3 ml浸泡,在室溫下經搖床振蕩24 h后過濾,收集濾過液置于貼有相同標簽的青霉素瓶內,4℃冰箱保存備用。
1.5 確定合適的篩選條件 分別挑取3種菌株的單菌落,于YC-UHW培養液中30℃,250 r·min-1振搖培養13 h,測其OD595nm,并用新鮮培養液調節OD值為0.1、0.08、0.04、0.02。將以上菌液每種每孔200 μl加入96孔板的第1、2兩列的12個孔,并在第3、4兩列設副孔,25℃,250 r·min-1振搖封閉培養,每12 h測量記錄OD值,直至其生長出現平臺期為止,繪制酵母生長曲線圖。通過該生長曲線確定此篩選條件下的最佳篩選起始濃度。

Tab 1 Pimers of PCR
分別將3種OD595nm為0.04的酵母菌液190 μl加入96孔板,相應孔加入相應劑量的無菌水和體積分數為0.5的乙醇,使得乙醇最終體積分數分別為0、0.05、0.10、0.15,總劑量200 μl,然后置于搖床中,25℃,250 r·min-1振搖培養,于12、24、36、48 h測量記錄OD值并繪制酵母生長曲線,通過生長曲線選擇合適的乙醇濃度(即樣品的劑量)。
1.6 高通量放線菌發酵液篩選 首先進行酵母菌株L40 ura3/pHybLex/Zeo-MS2/hTR126-451/hTERT 292-608 aa的篩選。選擇OD595nm為0.04的酵母菌液進行篩選。
實驗中,96孔板1~11列用來檢測樣品,每孔含10 μl樣品和190 μl菌液,第12列上面4孔作溶劑對照,即每孔10 μl體積分數為0.5的乙醇和190 μl菌液;中間2孔作空白對照,每孔10 μl無菌蒸餾水和190 μl菌液;最后2孔做負對照,每孔10 μl無菌蒸餾水和190 μl YC-UHW培養液。加液后于25℃,250 r·min-1振搖培養,分別于每隔12 h測量記錄OD595nm值。所有藥物樣品測試完畢后,再分別以L40 ura 3/pH ybLex/Zeo-MS2/hTR 240-450/hTERT 292-608 aa、L40 ura 3/pH ybLex/Zeo-MS2/hTR (m)/hTERT 292-952 aa篩選,操作程序同上。
1.7 菌液抑制率的計算 抑制率/%=(溶劑對照孔OD595nm-樣品孔OD595nm)/(溶劑對照孔 OD595nm-空白對照孔OD595nm)×100%
1.8 復篩 經初篩計算抑菌效率后,對所有抑菌效率達0.45以上的高活性放線菌發酵液樣品進行復篩。復篩中我們設立了菌株對照板,以L40 ura3/ pHybLex/Zeo-MS2(用YPD酵母培養液培養)為對照。篩選起始菌液濃度,藥物樣品劑量以及程序同初篩。
2 結果
2.1 酵母菌液PCR結果電泳圖 酵母菌液PCR,反應擴增產物用1.5%(Fig 1A)、0.8%(Fig 1B)瓊脂糖凝膠電泳鑒定,均為正確的片段。
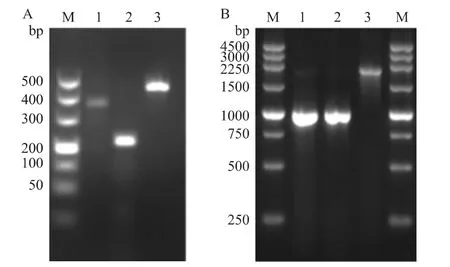
Fig 1 Electrophoresis of the three yeast PCR products
2.2 酵母菌生長曲線 實驗結果表明當酵母菌起始OD595nm為0.04是合適的酵母菌篩選起始濃度,其生長曲線見Fig 2A。在酵母菌液起始OD595nm為0.04時,乙醇體積分數為0.25時對酵母菌生長影響很小,其生長曲線見Fig 2B。
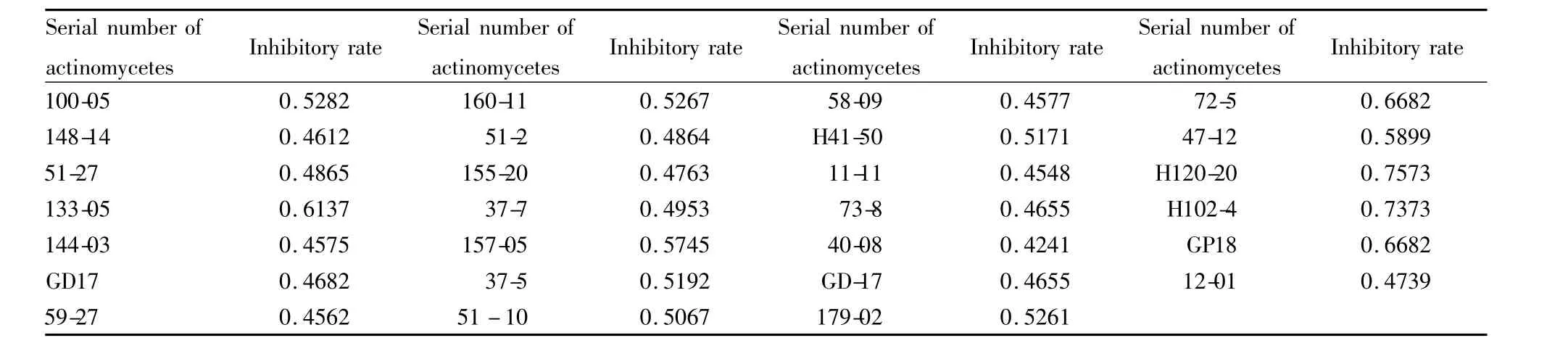
2.3 初篩結果 通過對2 000多株放線菌發酵液篩選,本實驗共篩出27株抑制率大于0.45的樣品,如Tab 2所示,可用于下一步復篩。
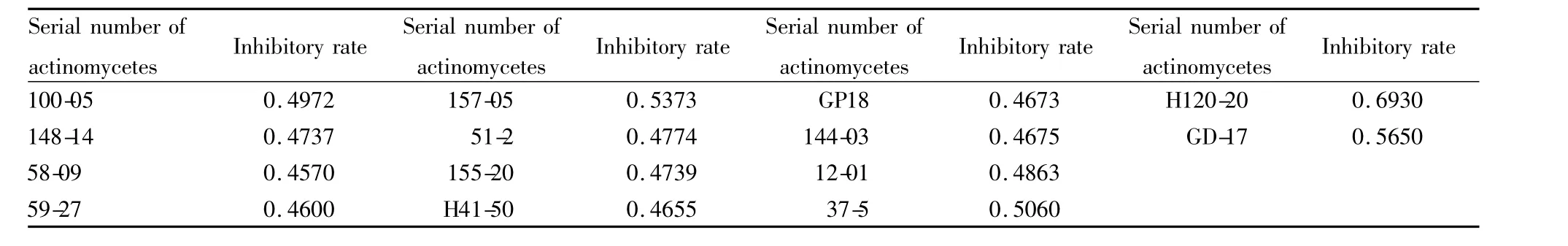
2.4 復篩結果 對27株放線菌發酵液進行復篩,篩出14株陽性樣品如Tab 3所示。
3 討論
端粒酶是廣譜腫瘤標志物之一,以端粒酶為靶標,建立腫瘤抑制劑篩選模型,尋找抗腫瘤新藥,已成為探索癌癥治療的新途徑[9]。但目前用于端粒酶抑制劑的篩選模型都是基于檢測端粒酶活性的TRAP法。該方法步驟繁瑣,成本高且特異性不強,不能用于RNA酶含量較高微生物代謝產物的篩選。而基于酵母三雜交系統的端粒酶抑制劑篩選模型克服了以上問題,在此系統中,hTERT和hTR的結合是已知的,當存在抑制兩者結合的因素時,酵母菌在酵母缺陷培養基上生長受抑制。因此只需通過培養酵母菌,觀察其生長狀態,就可進行端粒酶抑制劑的篩選。
Tab 2 The active samples and corresp inhibition rate in primary screening
Tab 3 The active samples and corresp inhibition rate in secondary screening
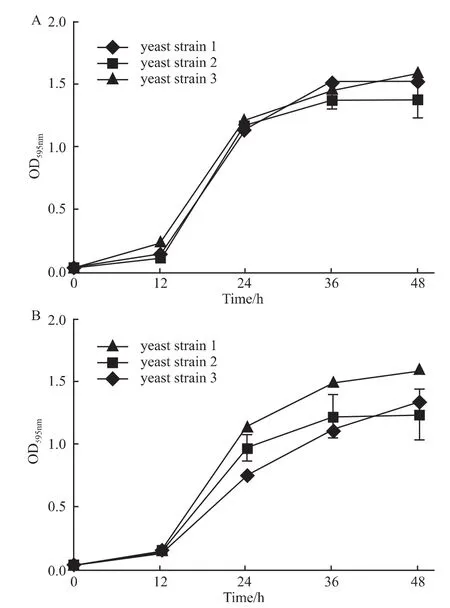
Fig 2 Growth curve of yeast strains
抑制劑的篩選,理論上只要能抑制hTERT和hTR的正確結合,就能抑制端粒酶活性,因此特異的抑制劑可能只抑制hTERT和hTR的其中一個結合部位,因此,為提高篩選模型的靈敏性和特異性,本實驗選擇了導入不同hTERT和hTR易結合結構片段的篩選模型,利用不同有效部位之間的相互作用來建立特異的酵母端粒酶抑制劑篩選模型。
由于放線菌發酵物的理化成分是未知的,為防止其有效成份不能完全溶出,本實驗使用0.5的乙醇作為代謝產物的溶出液。但是乙醇可以使蛋白類物質失活且對酵母菌的生長有較強的抑制作用,影響實驗的操作及結果,對此本實驗需尋找對酵母生長影響較小的乙醇濃度,以避免出現假陽性的結果。
酵母菌細胞壁厚,通透性差,實驗中需要延長培養時間以促進藥物進入細胞,而培養時間長,菌液起始培養濃度低,容易污染,因此,本實驗中采取酵母菌培養液添加氨芐青霉素(0.5 g·L-1),96孔板[10]封板培養等措施以避免菌液污染。
本實驗采用3種模型分別對2 000多株放線菌發酵液進行篩選,篩選模型菌在96孔板中培養48 h,每隔12 h測量并計算抑菌率,因此每個樣品對應12個抑菌率,將其中任一抑菌率大于0.45的放線菌發酵液視為陽性樣品,并取最高抑制率作為該樣品抑制率。初篩從樣品庫中篩選出的27株陽性樣品,其中編號為100-05、148-14、51-27、133-05、144-03、GD17、59-27的樣品為酵母菌 L40 ura3/pHybLex/Zeo-MS2/hTR126-451//hTERT292-608aa篩選而得;編號為51-27、144-03、160-11、51-2、155-20、37-7、157-05、37-5、51-10、58-09、H41-50、11-11、73-8樣品為酵母菌 L40 ura3/pHybLex/Zeo-MS2/hTR240-450/hTERT292-608aa篩選而得,40-08、GD17、GD-17、179-02樣品為酵母菌 L40 ura3/pHybLex/Zeo-MS2/hTR(m)/hTERT292-952aa篩選而得;而編號為72-5、47-12、H120-20、H102-4、GD18、12-01這6個樣品則對3種酵母菌篩選模型抑制劑率大于0.45。若一個樣品對兩個或三個模型抑制率均大于0.45時,總的抑制率取其平均值。復篩設立了陰性酵母菌對照,對初篩篩出的27株放線菌發酵液進行再篩,篩出14種陽性放線菌發酵液。初篩和復篩結果基本吻合,說明了本篩選模型和篩選方法的可靠性及有效性。
[1] Fletcher T M.Telomerase:a potential therapeutic target for cancer[J].Expert Opin Ther Targets,2005,9(3):457-69.
[2] Bachand F,Autexier C.Functional regions of human telomerase reverse transcriptase and human telomerase RNA required for telomerase activity and RNA-protein interactions[J].Mol Cell Biol,2001,21:1888-97.
[3] Hook B,Bernstein D,Zhang B,et al.RNA-protein interactions in the yeast three-hybrid system:affinity,sensitivity,and enhanced library screening[J].RNA,2005,11(2):227-33.
[4] Young K,Lin S,Sun L,et al.Identification of a calcium channel modulator using a high throughput yeast two-hybrid screen[J].Nat Biotechnol,1998,16(10):946-50.
[5] 曹 瑩,王衛國,李 琳,等.人端粒酶RNA突變體酵母三雜交誘餌質粒的構建[J].南方醫科大學學報,2009,29(4):652-5.
[5] Cao Y,Wang W G,Li L,et al.Construction of a bait plasmid containing human telomerase RNA mutant in yeast three-hybrid system[J].J South Med Univ,2009,29(4):652-5.
[6] 亞當斯A,特施林D E.酵母遺傳學方法實驗指南[M].劉子鐸譯.北京:科學出版社,2000:109.
[6] Adams A,Sheeran D E.Guide to yeast genetics experiment[M]. Translated by Liu ZY.Beijing:Science Press,2000:109.
[7] 薩姆布魯克J,拉塞爾D W.分子克隆實驗指南[M].北京:科學出版社,2002:1109-12.
[7] Sambrook J,Russell D W.Molecular Cloning A laboratory manual[M].Beijing:Science Press,2002:1109-12.
[8] Simple and reliable procedure for PCR amplification of genomic DNA from yeast cells using short sequencing primers[J].Biochen Mol Biol Int,1997,42(1):169-72.
[9] Greider C W,Blackburn E H.Telomeres Telomere and cancer[J]. Sci Am,1996,2:274(2):92-7.
[10]張 輝,喻莉萍,方 芳,方云祥.Edelfosine抑制S.pombe細胞分裂作用機制的研究[J].中國藥理學通報,2007,3(23):374-9.
[10]Zhang H,Yu L P,Fang F,Fang Y X.Study of the inhibition mechanism of edelfosine on cytokinesis in Schizosaccharomyces pombe[J].Chin Pharmacol Bull,2007,3(23):374-9.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小讀者(2021年2期)2021-03-29 05:03:48
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
光學精密工程(2016年6期)2016-11-07 09:07:19
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
