甲基蓮心堿對帕金森病細胞線粒體自噬的影響
2025-02-11 00:00:00陳翠青譚臣臣王電翠蔣敏
中國藥房 2025年2期
關鍵詞:帕金森病
中圖分類號 R965;R742.5 文獻標志碼 A 文章編號 1001-0408(2025)02-0197-06
DOI 10.6039/j.issn.1001-0408.2025.02.11
摘要 目的 探究甲基蓮心堿(NEF)調節腺苷一磷酸活化的蛋白激酶(AMPK)/哺乳動物雷帕霉素靶蛋白(mTOR)/UNC-51 樣激酶1(ULK1)信號通路對帕金森病(PD)細胞線粒體自噬的影響,從而探討該藥改善PD的作用機制。方法 用100 μmol/L 的1-甲基-4-苯基吡啶離子(MPP+)處理SH-SY5Y細胞24 h 以構建PD細胞模型,將PD模型細胞分為模型組(PD組)和NEF低、中、高濃度處理組(NEF-L、NEF-M、NEF-H組,2.5、5.0、10.0 μmol/L)以及NEF高濃度+AMPK抑制劑組(NEF-H+Compound C組,10.0 μmol/LNEF和50 μmol/L Compound C);另取未經MPP+、NEF處理的細胞作為對照組。觀察各組細胞的超微形態,檢測其自噬體數量、存活率、凋亡率、線粒體膜電位的變化情況,胱天蛋白酶3(Caspase-3)、微管相關蛋白1 輕鏈3(LC3)、Beclin-1 蛋白的表達水平,以及mTOR、AMPK、ULK1 蛋白的磷酸化水平。結果 與PD組比較,NEF-L、NEF-M、NEF-H組細胞自噬體明顯增多,膜電位有所升高,存活率、LC3-Ⅱ/LC3-Ⅰ、Beclin-1 蛋白的表達和AMPK、ULK1 蛋白的磷酸化水平均顯著升高或上調,凋亡率和Caspase-3、p62 蛋白的表達及mTOR蛋白的磷酸化水平均顯著降低或下調,且上述改善作用均呈濃度依賴性(P<0.05);Compound C能顯著逆轉高濃度NEF的上述改善作用(P<0.05)。結論 NEF通過上調AMPK、ULK1 蛋白的磷酸化水平,下調mTOR蛋白的磷酸化水平來促進PD模型細胞的線粒體自噬,抑制細胞凋亡,從而發揮神經細胞保護作用。
關鍵詞 甲基蓮心堿;帕金森病;線粒體自噬;AMPK/mTOR/ULK1 信號通路
帕金森病(Parkinson’s disease,PD)是全球第二常見的神經退行性疾病,全球患病人數超過600 萬[1]。由于老年人口不斷增長,PD發病率逐漸升高,其預防及治療以藥物為主,但副作用較多、療效有限[1]。PD 病理機制復雜且高度多樣,以靜止性震顫、肌肉僵硬、運動遲緩等多種運動癥狀為主要臨床表現;此外,非運動癥狀在PD患者中也較為常見,包括認知和行為障礙、睡眠不規律、感覺和自主神經功能障礙[2―3]。
甲基蓮心堿(neferine,NEF)是從蓮種子中提取的雙芐基異喹啉生物堿成分,具有抗炎、抗氧化、抗心律失常、抗高血壓、抗血栓形成、抗血小板聚集、改善遺忘等多種藥理活性,已被用于高熱、心律失常、高血壓、血小板聚集、肥胖和睡眠不足等癥的治療[4]。據報道,NEF可通過阻斷核因子κB信號轉導來抑制脂多糖介導的小膠質細胞活化,進而抑制慢性神經炎癥,對PD小鼠具有一定的保護作用[5],但具體作用機制尚不完全清楚。
研究指出,自噬是負責細胞清除和維持細胞穩態的生物學過程,可能是PD發病機制的重要組成部分[6]。腺苷一磷酸活化的蛋白激酶(adenosine monophosphateactivated protein kinase,AMPK)/哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)/UNC-51 樣激酶1(UNC-51-like kinase 1,ULK1)信號通路與自噬調節相關。其中,AMPK為高度保守的絲氨酸/蘇氨酸蛋白激酶,是調節代謝過程和炎癥反應的關鍵能量傳感器,可直接磷酸化mTOR 復合物1(mTOR complex 1,mTORC1)信號通路的若干組分,從而導致mTORC1 活性下調,減少mTOR的表達,下調的mTOR則可通過磷酸化促使ULK1 活化并促進自噬的發生[7]。據報道,厚樸酚可通過激活AMPK/mTOR/ULK1 信號通路促進自噬,從而改善阿爾茨海默病相關病理損傷[8]。但NEF是否能夠通過調節AMPK/mTOR/ULK1 信號通路來影響PD 細胞的線粒體自噬尚不清楚。為此,本研究基于AMPK/mTOR/ULK1 信號通路,初步探討NEF 對PD 細胞線粒體自噬的影響,旨在為闡明NEF改善PD的作用機制提供參考。
1 材料
1.1 主要儀器
本研究所用主要儀器包括GENios Plus 型酶標儀(成都市科邁普生物科技有限公司)、FormaTM Steri-CycleTM i160 型CO2 培養箱和Talos L120C TEM 型透射電鏡(美國Thermo Fisher Scientific 公司)、BX51 型熒光顯微鏡(日本Olympus 公司)、FACSymphonyTM A5 型流式細胞儀(美國BD公司)、Tanon EPS 600 型電泳儀(上海天能科技有限公司)等。
1.2 主要藥品與試劑
NEF對照品(批號SN8050,純度≥98%)購自北京索萊寶科技有限公司;AMPK抑制劑Compound C的對照品(批號HY-13418A,純度99.91%)、1-甲基-4-苯基吡啶離子(1-methyl-4-phenylpyridinium ion,MPP+;批號HYW008719,純度99.94%)均購自美國MCE公司;兔源胱天蛋白酶3(Caspase-3)、磷酸化AMPK(phosphorylatedAMPK,p-AMPK)、磷酸化mTOR(phosphorylatedmTOR,p-mTOR)、磷酸化ULK1(phosphorylated ULK1,p-ULK1)、AMPK、mTOR、ULK1、Beclin-1、β-肌動蛋白(β-actin)抗體和辣根過氧化物酶標記的羊抗兔免疫球蛋白G二抗、DMEM培養基(批號分別為MA5-45039、PA5-104982、44-1125G、PA5-78249、PA5-105297、PA5-34663、MA5-32699、PA1-16857、PA1-183、31460、11965092)均購自美國Thermo Fisher Scientific 公司;兔源p62、微管相關蛋白1 輕鏈3(microtubule-associated protein 1 lightchain 3,LC3)抗體(批號分別為ab109012、ab192890)均購自英國Abcam 公司;Annexin Ⅴ-FITC 細胞凋亡檢測試劑盒(批號WK304)購自北京百奧萊博科技有限公司;單丹磺酰尸胺(monodansyl cadaverine,MDC)試劑盒、線粒體膜電位試劑盒(批號分別為C3018S、C2006)均購自上海碧云天生物技術有限公司。
1.3 細胞
人神經母細胞瘤細胞SH-SY5Y(批號SNL-092)購自武漢尚恩生物技術有限公司。
2 方法
2.1 SH-SY5Y細胞培養與PD細胞模型建立
取SH-SY5Y細胞,在常規條件下培養,待其生長、融合至80%~90% 時進行實驗。取上述SH-SY5Y細胞,用100 μmol/L 的MPP+誘導培養24 h,以構建PD細胞模型(經MTT法檢測,若誘導后的細胞存活率較正常細胞顯著降低,則說明PD細胞模型構建成功)[9]。
2.2 NEF干預濃度篩選
依據相關文獻[6]及前期實驗結果,將構建成功的PD模型細胞用不同濃度的NEF(0、1.0、2.5、5.0、10.0、20.0μmol/L)處理24 h,另設不加MPP+、NEF 的對照組以及不加藥物、不加細胞的空白組,每組設置6 個復孔。處理后,每孔細胞加入MTT溶液10 μL,孵育4 h,加二甲基亞砜混勻,用酶標儀于490 nm波長下檢測其吸光度,并按下式計算細胞存活率:細胞存活率=(實驗組吸光度-空白組吸光度)/(對照組吸光度-空白組吸光度)×100%。根據細胞存活率檢測結果篩選最優藥物干預濃度,用于后續實驗。
2.3 細胞分組與給藥
將PD 模型細胞隨機分為PD 組和NEF 低、中、高濃度處理組(NEF-L、NEF-M、NEF-H 組)以及NEF 高濃度+AMPK 抑制劑組(NEF-H+Compound C 組),另取未經MPP+、NEF處理的SH-SY5Y細胞作為對照組,每組設6 個復孔。NEF-L、NEF-M、NEF-H組細胞分別用2.5、5.0、10.0 μmol/L 的NEF(濃度根據“2.2”項下結果設置)處理24 h,NEF-H+Compound C組細胞用10.0 μmol/L 的NEF和50 μmol/L的Compound C[10]共同處理24 h。
2.4 細胞增殖檢測
采用MTT法檢測。取對數生長期SH-SY5Y細胞,按1×104個/孔接種至96 孔板內,按“2.1”“2.3”項下方法造模、分組、處理。24 h 后,按“2.2”項下方法檢測各組細胞的存活率。
2.5 細胞超微形態觀察
采用透射電鏡觀察。取對數生長期SH-SY5Y 細胞,按1×104個/孔接種至96 孔板內,按“2.1”“2.3”項下方法造模、分組、處理。24 h后,以1 500 r/min離心5 min,收集細胞,以3% 戊二醛溶液固定,再經醋酸雙氧鈾和枸櫞酸鉛染色后,使用透射電鏡觀察各組細胞的超微形態。
2.6 細胞自噬體數量檢測
采用MDC 染色法檢測。取對數生長期SH-SY5Y細胞,按5×105個/孔接種至6 孔板內,按“2.1”“2.3”項下方法造模、分組、處理。24 h 后,收集細胞,用磷酸鹽緩沖液清洗,每孔加入5×10-5 mol/L 的MDC試劑,于室溫下避光孵育15 min,使用熒光顯微鏡觀察各組細胞內自噬體(呈綠色熒光)數量的變化情況。
2.7 細胞凋亡檢測
采用流式細胞術檢測。取對數生長期SH-SY5Y細胞,按5×105個/孔接種至6 孔板內,按“2.1”“2.3”項下方法造模、分組、處理。24 h 后,收集細胞,用磷酸鹽緩沖液清洗3 次,離心,棄上清液,細胞用培養基調整密度后,依次加入Annexin Ⅴ-FITC 緩沖液、PI 染液,于室溫下避光孵育30 min,使用流式細胞儀檢測各組細胞的凋亡情況,并記錄凋亡率(即流式圖右上和右下象限細胞百分率之和)。
2.8 細胞線粒體膜電位檢測
采用JC-1 探針法檢測。取對數生長期SH-SY5Y細胞,按5×105個/孔接種至6 孔板內,按照“2.1”“2.3”項下方法造模、分組、處理。24 h 后,收集細胞,用磷酸鹽緩沖液清洗,于37 ℃下加入0.5 μmol/L 的JC-1 探針試劑,避光染色1 h;收集細胞,用乙二胺四乙酸-胰蛋白酶分散后,使用熒光顯微鏡觀察細胞線粒體膜電位的變化情況(JC-1 為單體時,呈綠色熒光,提示線粒體發生損傷,膜電位較低;JC-1 為聚合物時,呈紅色熒光,提示線粒體損傷緩解,膜電位較高)。
2.9 細胞中凋亡、自噬相關蛋白表達檢測
采用Western blot 法檢測。取對數生長期SH-SY5Y細胞,按1×106個/孔接種至6 孔板內,按照“2.1”“2.3”項下方法造模、分組、處理。24 h 后,收集細胞,提取蛋白,檢測濃度后加熱變性。取變性蛋白適量,經電泳分離后轉移至聚偏二氟乙烯膜上,于室溫下以脫脂牛奶封閉;洗膜后,加入Caspase-3、p-mTOR、mTOR、p62、LC3、Beclin-1、p-AMPK、AMPK、p-ULK1、ULK1、β-actin 一抗(稀釋比例分別為1∶5 000、1∶1 000、1∶1 000、1∶10 000、1∶2 000、1∶2 000、1∶2 000、1∶2 000、1∶1 000、1∶5 000、1∶2 000),于4 ℃下孵育過夜;洗膜后,加入相應二抗(稀釋比例為1∶10 000),于室溫下孵育2 h;洗膜后,以ECL發光顯影、成像。使用Quantity One 軟件分析蛋白條帶的灰度值,以目的蛋白與內參蛋白(β-actin)的灰度值比值表示目的蛋白的表達水平,再以LC3-Ⅱ與LC3-Ⅰ、p-mTOR 與mTOR、p-AMPK 與AMPK、p-ULK1 與ULK1 的表達水平比值分別表示自噬活性和mTOR、AMPK、ULK1 蛋白的磷酸化水平。
2.10 統計學方法
采用SPSS 25.0 軟件對數據進行統計分析。實驗數據以x±s 表示,多組間比較采用單因素方差分析,進一步兩兩比較采用SNK-q檢驗。檢驗水準α=0.05。
3 結果
3.1 NEF干預濃度篩選結果
與對照組(100%)比較,經MPP+誘導而未經NEF處理的細胞的存活率[(53.64±5.21)% ] 顯著降低(P<0.05)。經MPP+誘導并經1.0、2.5、5.0、10.0、20.0 μmol/L的NEF 處理后,細胞的存活率分別為(56.47±6.25)%、(67.19±7.03)% 、(79.24±8.18)% 、(88.37±9.09)% 、(89.76±9.20)%;當NEF≥2.5 μmol/L 時,對應組別細胞的存活率均顯著高于未經NEF 處理的細胞(P<0.05),且經10.0、20.0 μmol/L NEF處理的細胞的存活率組間比較差異無統計學意義(P>0.05)。基于此,本研究選擇2.5、5.0、10.0 μmol/L作為后續實驗NEF的干預濃度。
3.2 細胞增殖變化
與對照組(100%)比較,PD 組細胞的存活率[(52.79±5.18)% ]顯著降低(P<0.05);與PD 組比較,NEF-L、NEF-M、NEF-H 組細胞的存活率[(68.23±7.09)%、(80.48±8.21)%、(89.78±9.14)%]均顯著升高,且呈濃度依賴性(P<0.05);與NEF-H組比較,NEF-H+Compound C 組細胞的存活率[(72.62±7.35)%]顯著降低(P<0.05)。
3.3 細胞超微形態變化
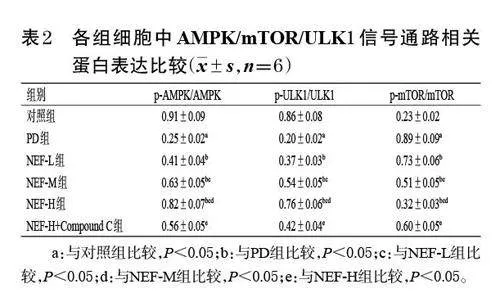
對照組細胞較為完整,形態正常,自噬體較少;與對照組比較,PD組細胞內可見自噬體形成,線粒體中空明顯;與PD組比較,NEF-L、NEF-M、NEF-H組細胞內部分線粒體中空并呈多膜樣結構,自噬體明顯增多,且上述變化有一定的濃度依賴趨勢;與NEF-H組比較,NEF-H+Compound C 組細胞內線粒體中空減少,自噬體明顯亦減少。結果見圖1。
3.4 細胞內自噬體數量變化
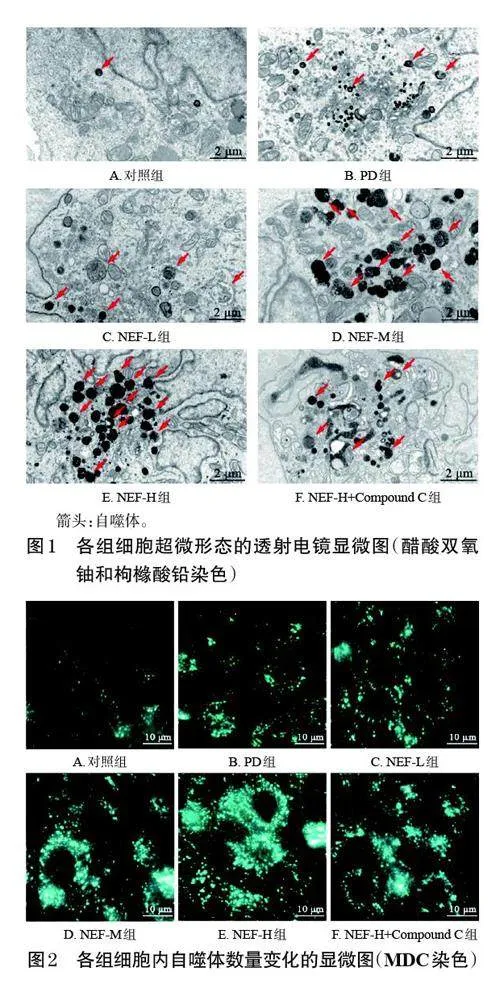
對照組細胞內綠色熒光較弱,自噬體較少;與對照組比較,PD組細胞內綠色熒光增強,自噬體增多;與PD組比較,NEF-L、NEF-M、NEF-H組細胞內綠色熒光進一步增強,自噬體進一步增多,且上述變化有一定的濃度依賴趨勢;與NEF-H組比較,NEF-H+Compound C 組細胞內綠色熒光減弱,自噬體減少。結果見圖2。
3.5 細胞凋亡情況
與對照組[(2.28±0.26)%]比較,PD 組細胞的凋亡率[(39.72±4.13)%]顯著升高(P<0.05);與PD組比較,NEF-L、NEF-M、NEF-H 組細胞的凋亡率[(32.58±3.31)%、(27.02±2.57)%、(21.04±2.10)%]均顯著降低,且呈濃度依賴性(P<0.05);與NEF-H組比較,NEF-H+Compound C 組細胞的凋亡率[(30.86±3.23)%]顯著升高(P<0.05)。各組細胞凋亡的流式圖略。
3.6 細胞線粒體膜電位變化
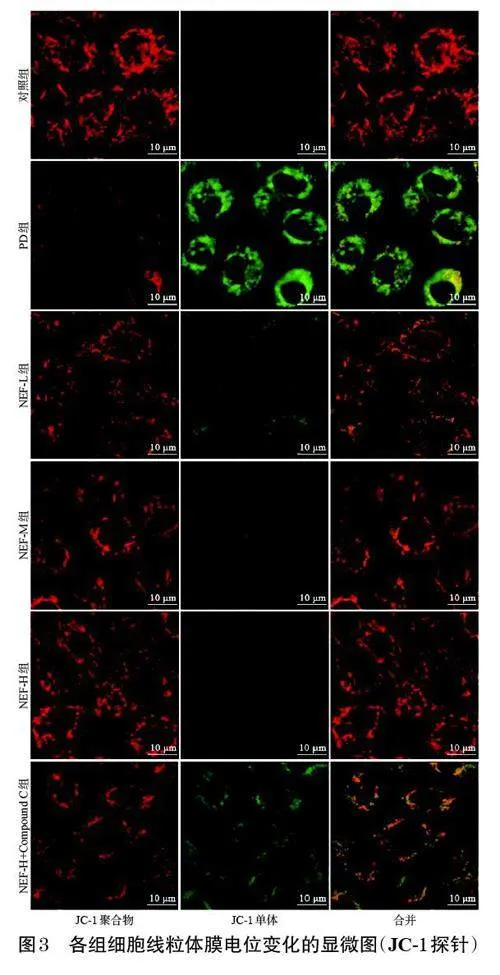
與對照組比較,PD組細胞的綠色熒光較強,紅色熒光較弱,線粒體發生損傷,膜電位較低;與PD 組比較,NEF-L、NEF-M、NEF-H組細胞的綠色熒光減弱,紅色熒光增強,線粒體損傷有所緩解,膜電位有所升高,且上述變化有一定的濃度依賴趨勢;與NEF-H組比較,NEF-H+Compound C組細胞的綠色熒光增強,紅色熒光減弱,線粒體發生損傷,膜電位降低。結果見圖3。
3.7 細胞中凋亡、自噬相關蛋白表達比較
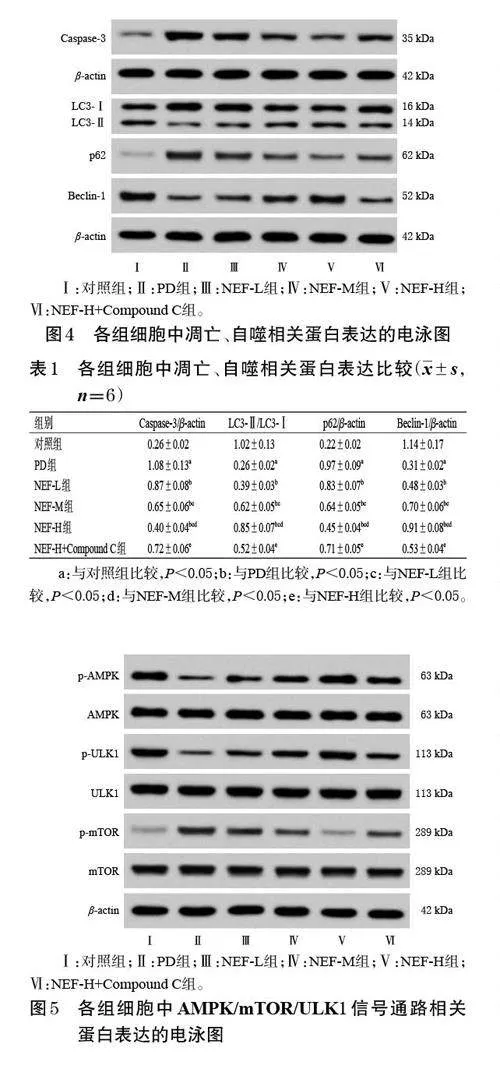
與對照組比較,PD 組細胞中LC3-Ⅱ/LC3-Ⅰ、Beclin-1 蛋白的表達水平均顯著降低,Caspase-3、p62 蛋白的表達水平均顯著升高(P<0.05);與PD組比較,NEFL、NEF-M、NEF-H組細胞中LC3-Ⅱ/LC3-Ⅰ、Beclin-1 蛋白的表達水平均顯著升高,Caspase-3、p62 蛋白的表達水平均顯著降低,且呈濃度依賴性(P<0.05);與NEFH組比較,NEF-H+Compound C 組細胞中LC3-Ⅱ/LC3-Ⅰ、Beclin-1 蛋白的表達水平均顯著降低,Caspase-3、p62 蛋白的表達水平均顯著升高(P<0.05)。結果見圖4、表1。
3.8 細胞中AMPK/mTOR/ULK1 信號通路相關蛋白表達比較
與對照組比較,PD 組細胞中AMPK、ULK1 蛋白的磷酸化水平均顯著降低,mTOR蛋白的磷酸化水平顯著升高(P<0.05);與PD 組比較,NEF-L、NEF-M、NEF-H組細胞中AMPK、ULK1 蛋白的磷酸化水平均顯著升高,mTOR蛋白的磷酸化水平均顯著降低,且呈濃度依賴性(P<0.05);與NEF-H組比較,NEF-H+Compound C組細胞中AMPK、ULK1 蛋白的磷酸化水平均顯著降低,mTOR蛋白的磷酸化水平顯著升高(P<0.05)。結果見圖5、表2。
4 討論
PD 發病率高,影響我國近2% 老年人的生活質量[11]。研究指出,PD的神經病理學特征是黑質中多巴胺能神經元的進行性丟失,伴有路易體和神經突的積累,其主要成分是聚集的α-突觸核蛋白(由SNCA 基因編碼),從而導致黑質紋狀體神經變性[11]。現有治療方法可以緩解PD 患者的早期癥狀,但不能阻止疾病進展或逆轉現有病情[11]。因此,尋找新的藥物對于PD 的臨床治療至關重要。
蓮子心是蓮種子的綠色胚,是一種可食用的中草藥,具有多種營養和藥用價值,可用于治療神經退行性疾病、失眠、高燒和心血管疾病[12]。NEF 是蓮子心所含的主要雙芐基異喹啉生物堿成分,具有抗心律失常、抗高血壓、抗癌、抗氧化、抗炎、治療神經退行性疾病等作用[4]。Wu 等[13]研究表明,NEF 可顯著提高SH-SY5Y 細胞的活力并降低其活性氧水平,抑制β淀粉樣蛋白和tau蛋白誘導的毒性,并增強線蟲的抗應激能力,具有神經保護作用。自噬是一種高度保守的分解代謝過程,可將異常蛋白和細胞器隔離到自噬體中,并將其遞送到溶酶體進行降解和回收,對細胞穩態調節、壓力適應和生存至關重要[14]。一項缺血性腦損傷研究指出,NEF可顯著抑制LC3-Ⅱ、Beclin-1 和p62 等自噬相關蛋白表達的上調,并阻止自噬體的形成[15]。本研究通過構建PD 細胞模型發現,細胞出現線粒體損傷并發生凋亡,自噬體明顯增加,說明經MPP+誘導后細胞發生了自噬和凋亡;經2.5、5.0、10.0 μmol/L 的NEF干預后,PD細胞中自噬體增多,自噬加重,凋亡率顯著降低,Caspase-3、p62 蛋白的表達顯著下調,LC3-Ⅱ/LC3-Ⅰ、Beclin-1 蛋白的表達顯著升高或上調,線粒體損傷有所改善,且上述作用具有濃度依賴性,提示NEF能夠促進細胞線粒體自噬進而緩解細胞凋亡。這一結果與文獻[15]有所不同,可能與所用模型不同、所涉疾病發生機制不同有關。然而,無論是促進自噬還是抑制自噬,兩項研究均證明了NEF對缺血性腦損傷和PD有一定的改善作用。
AMPK是能量代謝、細胞生長和自噬的重要調節因子[16]。AMPK作為關鍵的能量傳感器,可通過磷酸化來激活ULK1 蛋白,以維持細胞能量穩態并促進自噬,而mTOR是細胞生長的中樞調節劑和自噬的關鍵制動器,具有抑制ULK1 激活的能力[16]。研究報道,AMPK被激活后,ULK1 蛋白可以被磷酸化的AMPK直接激活,也可以通過mTOR的失活間接激活,進而導致顆粒細胞自噬[17]。據報道,激活AMPK/mTOR/ULK1 信號通路可提高LC3-Ⅱ/LC3-Ⅰ、Beclin-1 水平,降低p62 水平,從而促進自噬[8]。Zaki 等[18]研究發現,羅氟司特可通過上調p-AMPK、p-ULK1、Beclin-1 蛋白的表達和LC3- Ⅱ/LC3-Ⅰ,以及下調p62、p-mTOR蛋白的表達來恢復細胞的自噬功能。本研究結果顯示,經NEF處理后,PD模型細胞中AMPK、ULK1 蛋白的磷酸化水平均顯著升高,mTOR蛋白的磷酸化水平均顯著降低,與上述研究的結果基本一致。為明確NEF 的作用是否與AMPK/mTOR/ULK1信號通路有關,本研究在高濃度NEF 的基礎上聯用了AMPK抑制劑Compound C,結果顯示,該抑制劑逆轉了NEF對PD模型細胞各定量指標的改善作用。
綜上所述,NEF通過上調AMPK蛋白的磷酸化水平而使其活化,活化的AMPK蛋白可使下游ULK1 蛋白的磷酸化水平升高、mTOR蛋白的磷酸化水平降低,進而促進PD模型細胞線粒體自噬,抑制細胞凋亡,從而發揮神經細胞保護作用。本研究為未來NEF治療PD提供了可參考的實驗依據,但本研究僅通過體外細胞實驗初步證實了NEF與PD細胞自噬的關系,該成分能否在體內也發揮相同的作用及具體機制尚需相關動物實驗進一步驗證。
參考文獻
[ 1 ] LIZAMA B N,CHU C T. Neuronal autophagy and mitophagy
in Parkinson’s disease[J]. Mol Aspects Med,
2021,82:100972.
[ 2 ] MINCHEV D,KAZAKOVA M,SARAFIAN V. Neuroinflammation
and autophagy in Parkinson’s disease-novel
perspectives[J]. Int J Mol Sci,2022,23(23):14997.
[ 3 ] NECHUSHTAI L,FRENKEL D,PINKAS-KRAMARSKI
R. Autophagy in Parkinson’s disease[J]. Biomolecules,
2023,13(10):1435.
[ 4 ] LI S J,ZHANG Y Y,ZHANG J,et al. Neferine exerts
ferroptosis-inducing effect and antitumor effect on thyroid
cancer through Nrf2/HO-1/NQO1 inhibition[J]. J Oncol,
2022,2022:7933775.
[ 5 ] LI T,ZHAI Y X,ZHENG T Y,et al. Neferine exerts antiinflammatory
activity in BV-2 microglial cells and protects
mice with MPTP-induced Parkinson’s disease by inhibiting
NF-κB activation[J]. Mol Med Rep,2023,28
(6):235.
[ 6 ] KINET R,DEHAY B. Pathogenic aspects and therapeutic
avenues of autophagy in Parkinson’s disease[J]. Cells,
2023,12(4):621.
[ 7 ] HAN D,KIM D,KIM H,et al. Methylsulfonylmethane
ameliorates metabolic-associated fatty liver disease by
restoring autophagy flux via AMPK/mTOR/ULK1 signaling
pathway[J]. Front Pharmacol,2023,14:1302227.
[ 8 ] WANG X C,JIA J P. Magnolol improves Alzheimer’s
disease-like pathologies and cognitive decline by promoting
autophagy through activation of the AMPK/mTOR/
ULK1 pathway[J]. Biomed Pharmacother,2023,161:
114473.
[ 9 ] 彭秀華,柳明杰. MiR-486-5p 靶向TRPM2 對帕金森病
模型細胞自噬和凋亡的影響[J]. 中國老年學雜志,2021,
41(1):115-122.
PENG X H,LIU M J. Effect of miR-486-5p targeting
TRPM2 on autophagy and apoptosis in Parkinson’s disease
model cells[J]. Chin J Gerontol,2021,41(1):115-122.
[10] 時延龍,周鵬,王雪凱,等. 銀杏內酯K 調節AMPK/
mTOR/ULK1 信號通路對乳腺癌細胞自噬和凋亡的影
響[J]. 西部醫學,2024,36(1):42-46.
SHI Y L,ZHOU P,WANG X K,et al. Influences of
ginkgolide K on autophagy and apoptosis in breast cancer
cells by regulating AMPK/mTOR/ULK1 signaling pathway[
J]. Med J West China,2024,36(1):42-46.
[11] MA Z W,LIANG H,HU B C,et al. Autophagy-regulating
miRNAs:novel therapeutic targets for Parkinson’s disease:
review[J]. Int J Mol Med,2023,51(6):50.
[12] WANG M Y,ZHANG S S,AN M F,et al. Neferine ameliorates
nonalcoholic steatohepatitis through regulating
AMPK pathway[J]. Phytomedicine,2023,114:154798.
[13] WU M C,GAO Y H,ZHANG C,et al. Liensinine and
neferine exert neuroprotective effects via the autophagy
pathway in transgenic Caenorhabditis elegans[J]. BMC
Complement Med Ther,2023,23(1):386.
[14] TU H Y,YUAN B S,HOU X O,et al. α-synuclein suppresses
microglial autophagy and promotes neurodegeneration
in a mouse model of Parkinson’s disease[J]. Aging
Cell,2021,20(12):e13522.
[15] SENGKING J,OKA C,WICHA P,et al. Neferine protects
against brain damage in permanent cerebral ischemic
rat associated with autophagy suppression and AMPK/
mTOR regulation[J]. Mol Neurobiol,2021,58(12):6304-
6315.
[16] YANG C L,DENG Z W,ZENG Q H,et al. BMAL1 involved
in autophagy and injury of thoracic aortic endothelial
cells of rats induced by intermittent heat stress through
the AMPK/mTOR/ULK1 pathway[J]. Biochem Biophys
Res Commun,2023,661:34-41.
[17] LIN M L,HUA R,MA J,et al. Bisphenol A promotes autophagy
in ovarian granulosa cells by inducing AMPK/
mTOR/ULK1 signalling pathway[J]. Environ Int,2021,
147:106298.
[18] ZAKI E S,SAYED R H,SAAD M A,et al. Roflumilast
ameliorates ovariectomy-induced depressive-like behavior
in rats via activation of AMPK/mTOR/ULK1-dependent
autophagy pathway[J]. Life Sci,2023,327:121806.
(收稿日期:2024-07-04 修回日期:2024-12-24)
(編輯:鄒麗娟)
猜你喜歡
中老年保健(2022年5期)2022-08-24 02:34:58
中老年保健(2022年4期)2022-08-22 02:59:22
保健醫苑(2022年6期)2022-07-08 01:25:28
中老年保健(2021年6期)2021-11-30 08:25:27
中國民間療法(2021年10期)2021-07-22 02:32:40
成都醫學院學報(2021年2期)2021-07-19 08:35:14
生物學通報(2021年4期)2021-03-16 05:41:26
活力(2019年22期)2019-03-16 12:47:04
醫學研究雜志(2015年5期)2015-06-10 06:43:26
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
