強心湯對慢性心力衰竭大鼠線粒體的影響及機制
2025-02-11 00:00:00毛美玲盧健棋朱智德龐延謝麗鈺陳佳永吳鑫宇肖湘盧俊燊石煒琦
中國藥房 2025年2期
關鍵詞:模型

中圖分類號 R965 文獻標志碼 A 文章編號 1001-0408(2025)02-0160-06
DOI 10.6039/j.issn.1001-0408.2025.02.05
摘要 目的 探討強心湯對慢性心力衰竭(CHF)大鼠線粒體的影響及潛在機制。方法 采用結扎冠狀動脈左前降支的方法建立CHF模型。將造模成功的大鼠分為模型組,強心湯低、高劑量組(12.25、24.50 g/kg,以生藥量計),化學藥組(沙庫巴曲纈沙坦鈉片,10.42 mg/kg),另設不作處理的對照組,每組10 只。各組大鼠灌胃相應藥液或生理鹽水,每天2 次,連續28 d。末次給藥后,檢測大鼠血清中氨基末端腦利鈉肽前體(NT-proBNP)、三磷酸腺苷(ATP)和心肌組織中磷脂酸(PA)、心磷脂(CL)含量;觀察大鼠心肌組織病理損傷和膠原纖維化情況;檢測大鼠心肌細胞凋亡情況;觀察心肌組織超微結構;檢測心肌組織中線粒體融合蛋白1(Mfn1)、Mfn2、視神經萎縮蛋白1(OPA1)、動力相關蛋白1(Drp1)的表達情況。結果 與對照組比較,模型組大鼠血清中NT-proBNP 含量,心肌細胞凋亡率,心肌組織中S-OPA1、Drp1 蛋白的相對表達量均顯著升高(P<0.05);血清中ATP含量,心肌組織中PA、CL含量和Mfn1、Mfn2、L-OPA1 蛋白的相對表達量均顯著降低(P<0.05);心肌組織各層膜組織結構異常,心肌細胞變性壞死,且纖維化嚴重;心肌組織線粒體可見腫脹,嵴減少或消失,基質密度不均勻。經強心湯干預后,大鼠血清及心肌組織中上述定量指標水平(強心湯低劑量組CL含量除外)均顯著逆轉(P<0.05);心肌組織病理損傷明顯改善,纖維化情況明顯減輕;線粒體形態趨于正常且嵴增多,基質密度均勻。結論 強心湯可調節CHF大鼠的心肌線粒體功能和結構完整性,進而改善心肌能量代謝,減輕心肌纖維化;其作用機制可能與激活PA/Mfn/CL信號通路有關。
關鍵詞 強心湯;慢性心力衰竭;線粒體;能量代謝;纖維化;PA/Mfn/CL信號通路
慢性心力衰竭(chronic heart failure,CHF)是一種由微血管病變、低度全身性炎癥、氧化應激、線粒體障礙等多種因素聯合作用所導致的疾病,其典型病理變化為心肌重塑和纖維化[1]。研究表明,CHF患者心肌組織中三磷酸腺苷(adenosine triphosphate,ATP)含量降低和線粒體受損與其心功能下降密切相關[2];心肌細胞線粒體融合/分裂動態平衡對心肌能量代謝和收縮功能的維持具有積極作用,其動態平衡受損可能會導致CHF 加重[3]。研究指出,線粒體的結構和功能取決于自身外膜和內膜的分裂和融合,這些膜結構的重塑是由特定蛋白和脂質控制的,其中磷脂酸(phosphatidic acid,PA)、線粒體融合蛋白1(mitofusin 1,Mfn1)、Mfn2、心磷脂(cardiolipin,CL)與視神經萎縮蛋白1(optic atrophy protein 1,OPA1)在線粒體融合/分裂過程中扮演了重要角色,調控PA/Mfn/CL 信號通路對維持線粒體功能具有重要作用[4]。由此可見,調節心肌細胞線粒體穩態、保證心肌能量代謝成為了治療CHF的新思路。
中醫將CHF歸納為本虛標實證,氣虛、陽虛為本,血瘀、水飲為標。廣西中醫藥大學第一附屬醫院心內科盧健棋教授所創的強心湯以多靶點、多成分的作用優勢行益氣溫陽、活血利水之功,在治療CHF方面取得了良好的效果[5],但該方影響線粒體功能的上游調控機制尚不完全明確。基于此,本研究擬從PA/Mfn/CL 信號通路出發,探討強心湯對CHF大鼠心肌線粒體功能和能量代謝的影響及潛在機制,從而為強心湯的臨床應用提供實驗依據。
1 材料
1.1 主要儀器
本研究所用主要儀器包括MQX200 型酶標儀(美國BioTek 公司)、Tecnai G20 TWIN 型透射電鏡(美國FEI公司)、BX53 型熒光顯微鏡(日本Olympus 公司)、Flexacam C1 型顯微鏡(德國Leica 公司)、ChemiDocTMXRS+型超高靈敏度化學發光成像系統(美國Bio-Rad公司)、HMS-901B型制膠儀(深圳市博大精科生物科技有限公司)、DYCZ-40 型電轉儀(北京六一生物科技有限公司)等。
1.2 主要藥品與試劑
強心湯由黃芪、黨參、炮附子、桂枝、川芎、丹參、葶藶子、茯苓、白術、柏子仁、玉竹、炙甘草組成,各藥材飲片( 批號分別為230501、231225、230315、230807、220424、230814、221203、230617、230116、231002、220214、220723)均由廣西中醫藥大學第一附屬醫院提供,經該院心內科潘朝鋅主任中醫師鑒定為真品。
沙庫巴曲纈沙坦鈉片(批號090502,規格100 mg)購自北京諾華制藥有限公司;ATP 含量測定試劑盒(批號BC0305)購自北京索萊寶科技有限公司;兔源Mfn1、Mfn2 多克隆抗體(批號分別為NBP1-51841SS、NBP2-66383SS)均購自美國Novus Biologicals 公司;兔源動力相關蛋白1(dynamin-related protein 1,Drp1)單克隆抗體(批號ab184247)購自英國Abcam公司;鼠源β-肌動蛋白(β-actin)單克隆抗體(批號T0022)購自美國Affinity 公司;兔源OPA1 單克隆抗體、辣根過氧化物酶標記的羊抗鼠免疫球蛋白G 二抗(批號分別為27733-1-ap、SA00001-1)均購自武漢三鷹生物技術有限公司;辣根過氧化物酶標記的羊抗兔免疫球蛋白G 二抗(批號A0208)購自上海碧云天生物技術股份有限公司;ECL底物液(批號G2014)購自武漢賽維爾生物科技有限公司;PA、CL酶聯免疫吸附測定(ELISA)試劑盒(批號分別為MM-926522O2、MM-71760R2)均購自江蘇酶免實業有限公司;大鼠氨基末端腦利鈉肽前體(N-terminal probrainnatriuretic peptide,NT-proBNP)ELISA 試劑盒(批號E-EL-R3023)購自武漢伊萊瑞特生物科技股份有限公司。
1.3 實驗動物
本研究所用動物為SPF 級健康雄性SD 大鼠,共60只,體重為(180±20)g,購自湖南斯萊克景達實驗動物有限公司,動物生產許可證號為SCXK(湘)2019-0004。所有大鼠均飼養于廣西中醫藥大學科學實驗中心[溫度(25±2)℃、相對濕度(50±10)%,每12 h 明暗交替]內,自由攝食、飲水,適應1 周后進行實驗。本研究方案經廣西中醫藥大學實驗動物福利倫理委員會批準(批準編號為DW20231115-257)。
2 方法
2.1 分組與造模
隨機選取10 只健康大鼠作為對照組,其余50 只大鼠采用結扎冠狀動脈左前降支的方法建立CHF模型[6],具體操作如下:用2% 異氟烷麻醉大鼠,行氣管插管后連接小動物呼吸機,開胸暴露左冠狀動脈并結扎左前降支,若觀察到心室壁變為蒼白色且心電圖監測結果示ST段抬高,表明造模成功。隨后,將大鼠胸腔逐層縫合,撤掉氣管插管和呼吸機,術后連續給予青霉素3 d 預防感染。對照組大鼠不作處理。造模過程中,共有10 只大鼠死亡,將存活且造模成功的40 只大鼠隨機分為模型組,強心湯低、高劑量組,化學藥組(沙庫巴曲纈沙坦鈉片),每組10 只。
2.2 藥液配制與分組干預
強心湯含黃芪30 g、黨參15 g、炮附子10 g、桂枝10 g、川芎10 g、丹參10 g、葶藶子10 g、茯苓10 g、白術10 g、柏子仁12 g、玉竹10 g、炙甘草6 g。取處方量炮附子先煮1 h 去除毒性,然后與其他藥材共同煎煮,制成質量濃度分別為1.489 6、2.979 2 kg/L 的強心湯藥液(以生藥量計)。將沙庫巴曲纈沙坦鈉片碾碎后,溶解于水中,制成質量濃度為1.02 g/L的藥液。
于造模成功第2 天,強心湯低、高劑量組大鼠按12.25、24.50 g/kg 的劑量(以生藥量計,其中低劑量為臨床等效劑量)灌胃相應藥液,化學藥組大鼠按10.42 mg/kg的劑量(根據臨床等效劑量換算而得)灌胃沙庫巴曲纈沙坦藥液[7],對照組和模型組大鼠灌胃生理鹽水,每天2次,連續28 d。
2.3 取材
末次給藥后,以0.5% 戊巴比妥鈉(50 mg/kg)腹腔注射麻醉大鼠,于腹主動脈采血,離心后取上層血清,備用;繼續分離其心肌組織,取每組6 只大鼠的心肌組織,保存于低溫冰箱中,備用;另取每組1 只大鼠的心肌組織固定于戊二醛中,用于透射電鏡觀察;取每組剩余3 只大鼠的心肌組織,固定于4% 多聚甲醛中,用于病理觀察和TUNEL實驗。
2.4 大鼠血清中NT-proBNP、ATP 和心肌組織中PA、CL含量檢測
取“2.3”項下每組6 只大鼠的血清樣品適量,根據試劑盒說明書方法操作,采用ELISA 法檢測血清中NTproBNP含量,采用微量法檢測血清中ATP 含量。取“2.3”項下每組6 只大鼠凍存的心肌組織適量,加入磷酸鹽緩沖液于冰浴中充分研磨后,以3 000 r/min 離心10min,取上清液,根據試劑盒說明書方法操作,采用ELISA 法檢測心肌組織中CL 含量;取“2.3”項下每組6只大鼠凍存的心肌組織適量,用丁醇-甲醇-水(5∶25∶70,V/V/V)溶液抽提后,根據試劑盒說明書方法操作,采用ELISA法檢測大鼠心肌組織中PA含量。
2.5 大鼠心肌組織病理損傷觀察
分別采用蘇木精-伊紅(HE)、Masson 染色法觀察。取“2.3”項下各組大鼠固定于4% 多聚甲醛中的心肌組織適量,經脫水、透明、浸蠟、包埋、切片、烤片、脫蠟后,取部分切片,加蘇木精、伊紅染液等進行HE染色;另取部分切片,加重鉻酸鉀、蘇木精染液等進行Masson 染色;然后均用中性樹膠封片,采用顯微鏡觀察心肌組織病理變化和膠原纖維化情況,并拍照。
2.6 大鼠心肌細胞凋亡情況檢測
采用TUNEL法檢測。取“2.5”項下各組大鼠心肌組織剩余切片,于60 ℃下烘片3 h,脫蠟后以水浸洗,滴加稀釋好的蛋白酶K溶液,于室溫孵育15 min;以磷酸鹽緩沖液清洗3 次,滴加平衡緩沖液平衡15 min,吸掉多余液體后,滴加工作液,經4′,6- 二脒基-2- 苯基吲哚(DAPI)復染后,用中性樹膠封片,采用熒光顯微鏡觀察,并在400 倍鏡下統計細胞總數以及凋亡細胞(紅色)數,然后計算細胞凋亡率(細胞凋亡率=凋亡細胞數/細胞總數×100%)。
2.7 大鼠心肌組織超微結構觀察
采用透射電鏡觀察。取“2.3”項下各組大鼠固定于戊二醛中的心肌組織,用0.1 mol/L 磷酸鹽緩沖液漂洗15 min×3 次,再用1% 鋨酸溶液于室溫下固定2 h,于60 ℃下聚合48 h 后包埋,切片(厚約60~80 nm),再進行鈾鉛雙染色;次日,采用透射電鏡觀察心肌組織超微結構,并拍照。
2.8 大鼠心肌組織中Mfn1、Mfn2、OPA1、Drp1 蛋白表達檢測
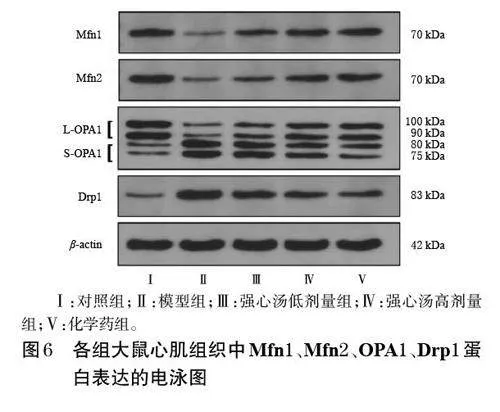
采用Western blot 法檢測。隨機選取“2.3”項下每組3 只大鼠凍存的心肌組織適量,加入RIPA 裂解液裂解后,采用BCA法測定蛋白濃度。將蛋白樣品變性后,進行電泳分離并轉移至聚偏二氟乙烯膜上,以脫脂奶粉于室溫下封閉2 h;洗膜后,分別加入Mfn1、Mfn2、OPA1、Drp1、β-actin 一抗(稀釋比例分別為1∶5 000、1∶5 000、1∶2 000、1∶1 000、1∶5 000),孵育過夜;洗膜后,加入相應二抗(稀釋比例均為1∶10 000),孵育2 h;洗膜后,以ECL底物液顯影,并于超高靈敏度化學發光成像系統下成像。采用Image J 軟件分析各目的蛋白的條帶灰度值,以β-actin為內參,計算目的蛋白的相對表達量。
2.9 統計學方法
使用SPSS 26.0 軟件對數據進行統計分析。實驗數據以x±s 表示,多組間比較采用單因素方差分析,進一步兩兩比較采用LSD-t 檢驗(方差齊)或Tamhane’s T2檢驗(方差不齊)。檢驗水準α=0.05。
3 結果
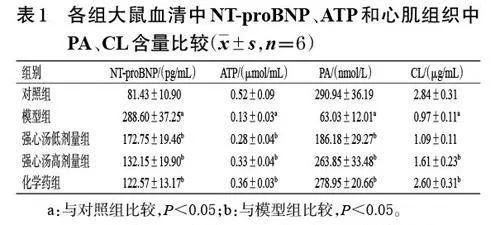
3.1 強心湯對CHF大鼠血清中NT-proBNP、ATP和心肌組織中PA、CL含量的影響
與對照組比較,模型組大鼠血清中NT-proBNP 含量顯著升高,ATP 含量顯著降低;心肌組織中PA、CL含量均顯著降低(P<0.05)。與模型組比較,各藥物組大鼠NT-proBNP 含量均顯著降低,ATP、PA、CL(強心湯低劑量組除外)含量均顯著升高(P<0.05)。結果見表1。
3.2 強心湯對CHF大鼠心肌組織病理變化的影響
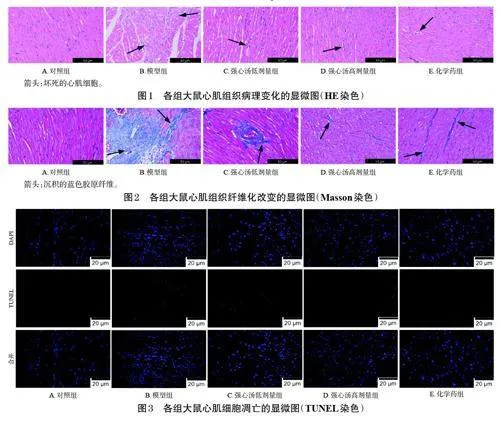
對照組大鼠心肌組織各層膜結構完整,細胞核染色正常,未見明顯病理損傷。模型組大鼠心肌組織病理損傷嚴重,心內膜、中膜和外膜結構異常,心肌細胞排列紊亂,細胞間隙增寬,膜破裂、腫脹且部分變性壞死,大量膠原纖維沉積,纖維化嚴重。各藥物組大鼠心肌組織病理損傷明顯改善,細胞壞死和纖維化情況明顯減輕。結果見圖1、圖2。
3.3 強心湯對CHF大鼠心肌細胞凋亡的影響
與對照組比較,模型組大鼠心肌細胞凋亡增多,凋亡率顯著升高(P<0.05);與模型組比較,各藥物組大鼠心肌細胞凋亡有所減少,凋亡率均顯著降低(P<0.05)。結果見圖3、圖4。
3.4 強心湯對CHF大鼠心肌組織超微結構的影響
對照組大鼠心肌組織線粒體形態規則,基質密度均勻;肌纖維排列整齊,肌絲清晰,Z 線和M線結構明顯。模型組大鼠心肌組織肌纖維紊亂,肌絲結構模糊,Z線和M線不清晰;線粒體腫脹、嵴減少或消失,基質密度不均勻。各藥物組大鼠心肌組織肌纖維排列整齊,肌絲結構清晰,Z 線和M線明顯;線粒體形態趨于正常,嵴增多,基質密度均勻。結果見圖5。
3.5 強心湯對CHF 大鼠心肌組織中Mfn1、Mfn2、OPA1、Drp1 蛋白表達的影響
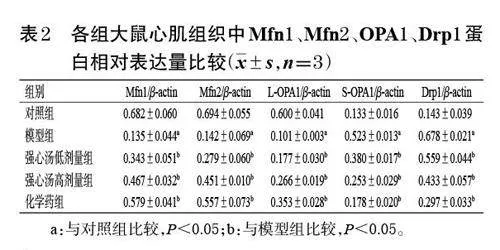
與對照組比較,模型組大鼠心肌組織中Mfn1、Mfn2、L-OPA1 蛋白的相對表達量均顯著降低,S-OPA1、Drp1 蛋白的相對表達量均顯著升高(P<0.05);與模型組比較,各藥物組大鼠心肌組織中Mfn1、Mfn2、LOPA1蛋白的相對表達量均顯著升高,S-OPA1、Drp1 蛋白的相對表達量均顯著降低(P<0.05)。結果見圖6、表2。
4 討論
中醫認為,心臟的正常運作和生命活動的維持需要靠心氣推動血液運行于周身,心氣不足則無力運行血液以致形神不得榮養;陰陽同源,氣虛日久則損及陽氣,陽虛加之“血不利則為水”,水飲漸生,如此循環,則變為CHF[8]。因此,中醫治療CHF 應以補益心氣為基礎,溫補、調和心陰心陽,輔以利水、化瘀,使邪有出路,正氣得到扶助。
心臟中線粒體含量非常豐富。線粒體可通過氧化磷酸化產生高達90% 的ATP,為心肌收縮和舒張提供能量;當細胞線粒體受到內部DNA突變影響或外部刺激時,線粒體外膜通透性將發生改變,從而釋放細胞凋亡因子以促進細胞凋亡,最終影響線粒體的能量代謝[9]。NT-proBNP 是臨床評估心臟功能的重要指標[10]。本研究結果顯示,模型大鼠血清中NT-proBNP 含量顯著升高,同時,HE、Masson 染色結果顯示,其心肌組織病理損傷、纖維化嚴重。經強心湯干預后,模型大鼠血清中NTproBNP含量顯著降低,心肌組織病理損傷和纖維化情況均得到改善,細胞凋亡率降低。這表明強心湯能夠在一定程度上發揮拮抗CHF的作用。
PA主要在內質網中生成,CL則主要由PA合成。當PA從內質網轉運到線粒體內膜后,其可通過多種酶促反應轉化為CL;隨后,部分CL 又被轉運到線粒體外膜上,經磷脂酶D誘導轉化為PA;兩者的相互轉化可參與控制線粒體的融合和分裂,并協調上述動態過程之間的平衡,從而維持線粒體穩態[11―12]。Mfn1、Mfn2 是絲裂霉素家族的重要成員,當心肌缺血缺氧時,PA能夠刺激線粒體外膜上的Mfn1 和Mfn2 與另一線粒體的七肽重復序列1 結構域相融合,形成七肽重復序列2 結構域;而位于七肽重復序列2 結構域的兩個半胱氨酸可因氧化型谷胱甘肽水平升高而被氧化,導致Mfn1 和Mfn2 之間形成二硫鍵,從而促進兩個線粒體的外膜發生融合[13]。OPA1可被切割成數個多肽片段,其中分子量較高的被稱為LOPA1,分子量較低的被稱為S-OPA1;S-OPA1 可促進線粒體裂變,而L-OPA1 可直接與線粒體內膜中含有CL的脂質體相互作用,從而介導線粒體內膜的融合[14]。相關研究發現,在線粒體內膜融合過程中,OPA1 與Mfn1 可能存在相互依賴的關系,但與Mfn2 可能不存在相互依賴的關系[15]。由此可知,線粒體融合是PA 刺激Mfn1、Mfn2 介導的線粒體外膜融合和CL與OPA1 相互作用介導的線粒體內膜融合兩個生物過程。除參與線粒體融合以外,PA和CL在線粒體分裂過程中也發揮了相應的作用,這與PA和CL特殊的脂質體結構有關[16]。Drp1 是一種可溶性蛋白,在線粒體分裂過程中,一部分Drp1 能夠特異性識別PA的酰基鏈,并優先與PA飽和酰基鏈結合;另一部分Drp1 能穿透線粒體的脂質雙層膜,與PA和磷脂酰膽堿結合,這種特殊的脂質結合作用可抑制Drp1誘導的線粒體分裂[17―18]。本研究結果顯示,與對照組比較,模型組大鼠心肌組織中PA、CL含量和Mfn1、Mfn2、L-OPA1 蛋白的相對表達量均顯著降低,S-OPA1、Drp1蛋白的相對表達量均顯著升高,表明CHF大鼠心肌組織線粒體內、外膜融合減少,而分裂增加。經強心湯干預后,模型大鼠心肌組織中PA、CL含量和Mfn1、Mfn2、LOPA1蛋白的相對表達量均顯著升高,S-OPA1、Drp1 蛋白的相對表達量均顯著降低,進一步結合透射電鏡和ATP檢測結果發現,強心湯可恢復心肌組織受損線粒體結構和功能,增強心肌能量代謝。
綜上所述,強心湯可調節CHF大鼠的心肌線粒體功能和結構完整性,進而改善心肌能量代謝、減輕心肌纖維化,其作用機制可能與激活PA/Mfn/CL 信號通路有關。盡管本研究已從作用途徑、表型等方面盡可能充分闡釋強心湯的作用途徑,然而仍存在一定的局限性,如并未在體外實驗中同步驗證,且僅正向驗證了該通路,未設置靶標敲低組,以進一步反向驗證該機制,故有待深入研究。
參考文獻
[ 1 ] BORLAUG B A,PAULUS W J. Heart failure with preserved
ejection fraction:pathophysiology,diagnosis,and
treatment[J]. Eur Heart J,2011,32(6):670-679.
[ 2 ] CHISTIAKOV D A,SHKURAT T P,MELNICHENKO A
A,et al. The role of mitochondrial dysfunction in cardiovascular
disease:a brief review[J]. Ann Med,2018,50
(2):121-127.
[ 3 ] SCHIRRMACHER V. Mitochondria at work:new insights
into regulation and dysregulation of cellular energy supply
and metabolism[J]. Biomedicines,2020,8(11):526.
[ 4 ] ROY M,REDDY P H,IIJIMA M,et al. Mitochondrial division
and fusion in metabolism[J]. Curr Opin Cell Biol,
2015,33:111-118.
[ 5 ] 毛美玲,盧健棋,謝麗鈺,等. 基于網絡藥理學、分子對接
探討強心湯治療慢性心力衰竭的潛在作用機制[J]. 中醫
雜志,2023,64(20):2132-2137.
MAO M L,LU J Q,XIE L Y,et al. Potential mechanism
of action of Qiangxin decoction for chronic heart failure
based on network pharmacology and molecular docking
[J]. J Tradit Chin Med,2023,64(20):2132-2137.
[ 6 ] 王飛燕,李學文,仝凌. 影響大鼠心肌梗死模型存活率的
因素分析[J]. 中西醫結合心腦血管病雜志,2008,6(7):
862-863.
WANG F Y,LI X W,TONG L. Analysis of factors affecting
the survival rate of rat myocardial infarction model[J].
Chin J Integr Med Cardio Cerebrovasc Dis,2008,6(7):
862-863.
[ 7 ] 黃繼漢,黃曉暉,陳志揚,等. 藥理試驗中動物間和動物
與人體間的等效劑量換算[J]. 中國臨床藥理學與治療
學,2004,9(9):1069-1072.
HUANG J H,HUANG X H,CHEN Z Y,et al. Dose conversion
among different animals and healthy volunteers in
pharmacological study[J]. Chin J Clin Pharmacol Ther,
2004,9(9):1069-1072.
[ 8 ] 盧健棋,唐梅玲,朱智德,等. 以中醫思維認識心力衰竭
[J]. 中醫學報,2021,36(8):1600-1603.
LU J Q,TANG M L,ZHU Z D,et al. Understanding heart
failure from TCM thinking[J]. Acta Chin Med,2021,36
(8):1600-1603.
[ 9 ] FAN H L,HE Z J,HUANG H F,et al. Mitochondrial
quality control in cardiomyocytes:a critical role in the progression
of cardiovascular diseases[J]. Front Physiol,
2020,11:252.
[10] ZHU F,ARSHI B,LEENING M J G,et al. Sex-specific
added value of cardiac biomarkers for 10-year cardiovascular
risk prediction[J]. Eur J Prev Cardiol,2022,29
(11):1559-1567.
[11] KAMEOKA S,ADACHI Y,OKAMOTO K,et al. Phosphatidic
acid and cardiolipin coordinate mitochondrial dynamics[
J]. Trends Cell Biol,2018,28(1):67-76.
[12] NELSON R K,FROHMAN M A. Physiological and
pathophysiological roles for phospholipase D[J]. J Lipid
Res,2015,56(12):2229-2237.
[13] TILOKANI L,NAGASHIMA S,PAUPE V,et al. Mitochondrial
dynamics:overview of molecular mechanisms
[J]. Essays Biochem,2018,62(3):341-360.
[14] MACVICAR T,LANGER T. OPA1 processing in cell
death and disease:the long and short of it[J]. J Cell Sci,
2016,129(12):2297-2306.
[15] MATTIE S,RIEMER J,WIDEMAN J G,et al. A new mitofusin
topology places the redox-regulated C terminus in
the mitochondrial intermembrane space[J]. J Cell Biol,
2018,217(2):507-515.
[16] FROHMAN M A. Role of mitochondrial lipids in guiding
fission and fusion[J]. J Mol Med,2015,93(3):263-269.
[17] FRANCY C A,ALVAREZ F J,ZHOU L,et al. The
mechanoenzymatic core of dynamin-related protein 1
comprises the minimal machinery required for membrane
constriction[J]. J Biol Chem,2015,290(18):11692-11703.
[18] STEPANYANTS N,MACDONALD P J,FRANCY C A,
et al. Cardiolipin’s propensity for phase transition and its
reorganization by dynamin-related protein 1 form a basis
for mitochondrial membrane fission[J]. Mol Biol Cell,
2015,26(17):3104-3116.
(收稿日期:2024-06-13 修回日期:2024-10-08)
(編輯:唐曉蓮)
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
