哈茨木霉菌對煙草的促生及其黑脛病的誘導抗性評價
2024-12-31 00:00:00常峻嘉蓋佳鑫陶剛莫轉龍海
中國農業科技導報 2024年10期

關鍵詞:哈茨木霉;促生作用;煙草黑脛病;誘導抗性;生物防治
煙草是我國的重要產業,為國家的財政收入做出了巨大貢獻。同時,煙葉中提取的煙堿可用于化工、制造醫藥和農藥等領域。煙草黑脛病是由煙草疫霉菌(Phytophthora nicotianae)引起、可通過土壤傳播的一種嚴重病害,在煙草整個生長階段均可發生,最常見的癥狀為莖基部呈黑色[1],能導致煙草產量下降甚至絕收[2]。施用化學農藥會造成嚴重的環境污染[3],同時引發病菌產生耐藥性或抗藥性,通過生物防治解決煙草病害是重要方式[4],也是現代農業作物病害綠色防控的重要方向之一,為解決現代農業生產中農藥殘留和環境污染問題提供了有效途徑。
木霉菌(Trichoderma spp.)是一類在全世界廣泛分布的真菌資源,其種類超過400種,截至2020年,有效命名的木霉種類有375種[5]。木霉菌也是一類重要的生防真菌,在2004年被列為無毒的植物共生真菌[6]。因其生長速度快、生存能力強且具有很強的植物病害生物防治能力等特點在農業實踐中得到廣泛應用。木霉能夠促進不溶性化合物質溶解、微量元素的利用及植物根系生長和對營養物質的吸收,從而促進植物生長發育[7]。研究表明,多種木霉在煙草黑脛病的生物防治中效果顯著,如哈茨木霉(Trichoderma harzianum)、棘孢木霉(T. asperellum)、長枝木霉(T. longibrachiatum)[8-10]。同時,木霉可誘導宿主植物產生抗性。薛春生等[11]研究表明,哈茨木霉TR409可誘導辣椒植株提高抗病性,TR409處理后辣椒植株的苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)、過氧化物酶(peroxisome,POD)、多酚氧化酶(polyphenoloxidase,PPO)和超氧化物歧化酶(superoxidedismutase,SOD)活性較對照組顯著提升。匡志豪等[12]在哈茨木霉對煙草黑脛病的誘導抗性研究中發現,處理組根系的POD、過氧化氫酶(catalase,CAT)等抗氧化酶活性均顯著提高。
研究表明,在植物的病害侵染反應中存在系統獲得抗性(systemic acquired resistance, SAR)和誘導系統抗性(Induced systemic resistance,ISR)2種關鍵系統抗性[1314],其中水楊酸是誘發SAR 的關鍵信號分子,而茉莉酸和乙烯是誘發ISR反應的關鍵信號分子。NPR1、PDF1.2、EFE 和NtSGT1這4個基因是水楊酸、茉莉酸和乙烯代謝合成途徑的關鍵基因[15]。EFE 基因編碼的乙烯合成酶是植物在響應脅迫、抵御病害過程中的關鍵信號物質;NtSGT1 基因通過激活半乳糖苷酶誘導煙草產生抗性[16]。
目前利用木霉菌防治煙草病害的研究報道較多,但木霉對不同煙草品種抗病防御酶活性的影響和誘導抗病相關基因表達的影響還缺乏系統研究。因此,本研究以紅花大金元和K326煙草品種為研究對象,對5株木霉菌進行平板對峙拮抗試驗,篩選對煙草疫霉菌具有強拮抗作用的木霉菌株;并進一步采用溫室盆栽試驗,篩選對煙草黑脛病具有生物防治效果的菌株,分析該菌株處理下煙草植株抗病相關防御酶的活性及調控誘導信號分子代謝途徑關鍵基因的表達變化,探究木霉-煙草-煙草疫霉菌三者間的相互作用,為研究植物病害生物防治和木霉誘導植物抗病機制提供優秀菌株資源和理論基礎。
1 材料與方法
1.1 試驗材料
1.1.1 供試木霉菌株與病原菌株 供試哈茨木霉ACCC 32808(TH32808)、哈茨木霉ACCC 32809、鉤狀木霉(T. hamatum)ACCC 31649、深綠木霉(T. atroviride)ACCC 32826、非洲哈茨木霉(T.afroharzianum)ACCC 32969由中國農業科院農業資源與農業區劃研究所顧金剛課題組保存。煙草疫霉菌由云南煙草科學研究院方敦煌老師提供。
1.1.2 煙草品種 供試煙草品種為K326和紅花大金元(Honghuadajinyuan,H),由河南農業大學煙草學院殷全玉老師提供。
1.1.3 培養基 PDA(potato dextrose agar)培養基購買于Oxoid(英國)公司。
燕麥培養基:燕麥60 g煮1 h后過濾,取上清加入蔗糖20 g、瓊脂粉16 g,蒸餾水定容至1 000 mL。
1.2 試驗方法
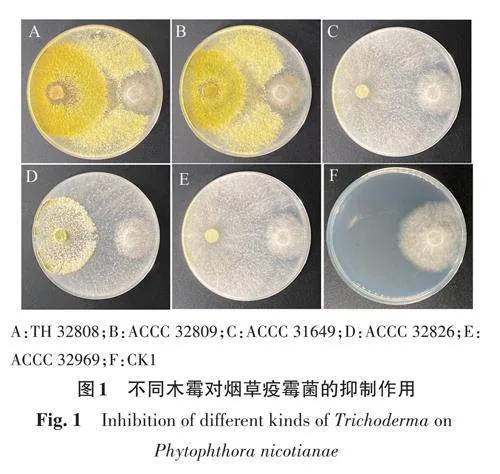
1.2.1 木霉菌株與病原菌的平板拮抗試驗 將5株木霉菌在PDA 培養基上培養7 d;煙草疫霉菌于燕麥培養基上活化7 d,備用。用直徑為5 mm的打孔器取煙草疫霉菌和5株木霉菌菌餅各1塊,分別接種于90 mm培養皿,距中心2.5 cm的兩側,使木霉與煙草疫霉間的距離為5 cm。以單獨接種煙草疫霉菌餅為對照(CK1),每處理5次重復,用封口膜封口,28 ℃黑暗倒置培養5 d后,計算抑菌率(%),公式如下。
1.2.2 煙草苗的培育 將煙草種子經10% H2O2消毒30 min后,用無菌水清洗4~5次,用槍頭吸至裝有營養土的32 孔育苗缽(55 mm×27 mm ×60 mm)中,待煙草苗齡為40 d時備用。
1.2.3 孢子懸浮液制備 木霉孢子懸浮液:將活化的木霉菌餅轉接至PDA 平板上,在黑暗條件下,28 ℃倒置培養1周,向培養好的木霉平板中加入10 mL無菌水,用槍頭將菌絲和孢子洗下,倒入加有玻璃珠的三角瓶中,將菌絲打碎后過4層紗布過濾,得到的液體即為木霉孢子懸浮液,調節TH32808孢子懸浮液至1×108個孢子·mL?1。
煙草疫霉菌孢子懸浮液:參考羅秀娟等[17]的方法制備煙草疫霉游動孢子懸浮液。將活化的煙草疫霉菌菌餅轉接至燕麥培養基平板,黑暗條件下,28 ℃倒置培養2周,向平板中加入0.1%硝酸鉀,培養2~3 d后洗至三角瓶中,將菌絲搗碎過4層紗布過濾,放入4 ℃冰箱20~30 min后,即為煙草疫霉孢子懸浮液,將其調整至1×105個孢子·mL?1。
1.2.4 木霉菌對煙草植株的促生作用 取40 d的紅花大金元和K326 煙苗進行移栽,盆栽基質為營養土∶珍珠巖∶蛭石=4∶1∶1(體積比)。緩苗7 d后,用TH32808孢子懸浮液10 mL對煙草苗進行灌根處理,以不接種的煙草苗為對照(CK2),每處理5次重復。將處理和對照的煙草苗均置于溫室自然光下培養,30 d后記錄煙草的株高、莖粗、鮮重、干重,計算促生率(%)。
1.2.5 木霉對煙草黑脛病的防治 取40 d的紅花大金元煙苗進行移栽,盆栽基質為營養土∶珍珠巖∶蛭石=4∶1∶1(體積比)。緩苗7 d后,采用灌根處理接種TH32808 孢子懸浮液10 mL;2 d 后,接種煙草疫霉菌孢子懸浮液5 mL;以單獨接種煙草疫霉菌孢子懸浮液5 mL為對照(CK3),每處理6次重復。參照GB/T 23222—2008[18]中煙草黑脛病分級標準,以株為單位,統計煙草植株15 d內的病害情況,計算發病率(%)、病情指數、防治效果(%)。
1.2.6 煙草防御酶活性的測定 煙草苗移栽與灌根方法同1.2.4,共設計4個處理,以不接種菌作為對照(CK4);分別設置單獨接種TH32808孢子懸浮液10 mL(TH32808);接種TH32808孢子懸浮液10 mL,2 d 后接種煙草疫霉孢子懸浮液5 mL(TH32808+P);單獨接種煙草疫霉孢子懸浮液5 mL(P)。分別在處理2、4、6 d時取頂部3片葉各1 g,并將其混勻,液氮研磨后用于煙草各防御酶活性的測定。酶活性測定參考索萊寶活性檢測試劑盒說明書。
1.2.7 TH32808 誘導煙草抗性相關基因表達測定 煙草苗移栽和灌根方法同1.2.4。接種TH32808孢子懸浮液10 mL,2 d后接種煙草疫霉孢子懸浮液5 mL;以不接種菌的煙苗為對照(CK5)。分別于處理1、2、3、4 d 時取幼苗頂部3片葉,將其混勻液氮研磨后,取樣50~100 mg,采用Vazyme RC401 試劑盒提取葉片總RNA,再利用Vazyme逆轉錄試劑盒合成cDNA,置于?20 ℃備用。利用ChamQ SYBR qPCR Master mix(Vazyme)在ViiATM7(美國ABI公司)上進行qRT-PCR檢測。PCR引物見表1,由北京擎科生物科技股份有限公司合成。PCR 體系為20 μL,包括10 μL 2×ChamQ SYBR qPCR Master Mix,0.4 μL 正向引物,0.4 μL 反向引物,0.4 μL 50×ROX Reference Dye1,1 μL Template DNA/cDNA,7.8 μL ddH2O。PCR程序:95 ℃預變性 30 s;95 ℃變性 10 s,60 ℃退火20 s,循環40次;60 ℃延伸60 s。參照Schmittgen等[19]的方法,以β-actin 為內參基因計算相對基因的表達水平,3次重復。
1.3 數據統計與分析
采用Excel軟件統計數據,通過DPS 7.05軟件進行數據分析,并用最小顯著性差異(leastsignificant difference,LSD)法進行差異顯著性檢驗。
2 結果與分析
2.1 木霉菌對病原菌的抑制效果
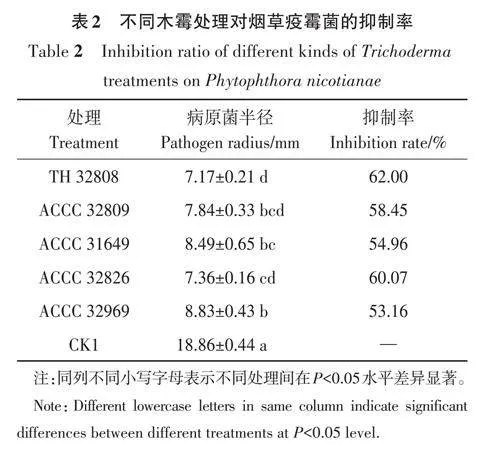
由圖1可知,5株木霉菌處理下病原菌的菌落半徑與CK差異較大,且木霉菌在抑制病原菌生長的同時,對病原菌的產孢也會有不同程度的抑制。將煙草疫霉菌與5株木霉菌對峙培養5 d后,其菌落直徑均顯著小于CK,其中TH32808對煙草疫霉菌的抑制效果最好,抑菌率為62.00%(表2)。
2.2 木霉對煙草植株的促生作用
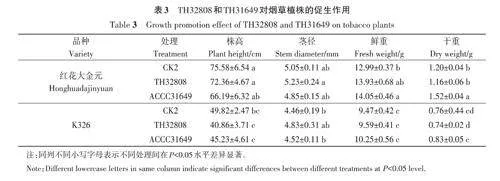
采用木霉菌株TH32808 和ACCC31649 的孢子懸浮液灌根處理30 d,結果(表3)表明,TH32808對紅花大金元和K326煙草植株的莖徑、鮮重有促進作用,分別較CK2增加3.56%、8.30%和7.24%、1.27%,說明不同木霉菌株對同一煙草品種或同一木霉菌株對不同煙草品種的促生作用存在差異。
2.3 木霉對煙草黑脛病的生物防治效果
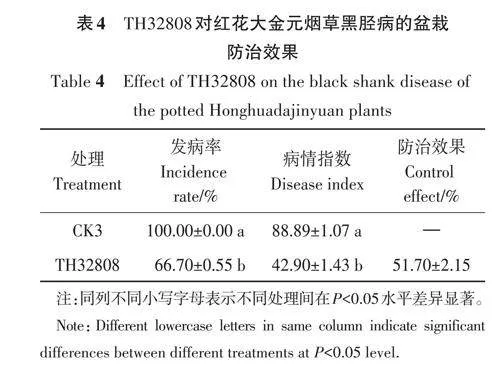
K326煙草品種在試驗過程中由于多重因素導致死亡,因此僅對紅花大金元煙草品種進行分析。由表4可知,單獨接種煙草疫霉菌(CK3)時,其黑脛病的發病率為100.00%,病情指數為88.89;而TH32808孢子懸浮液灌根處理后,煙草植株的發病率降為66.70%,病情指數降至42.90,與CK3相比,防治效果為51.70%。
2.4 哈茨木霉ACCC 32808 對煙草植株防御酶活性的影響
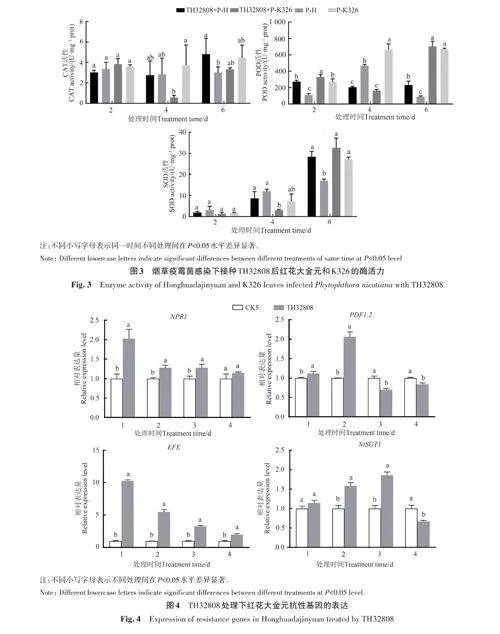
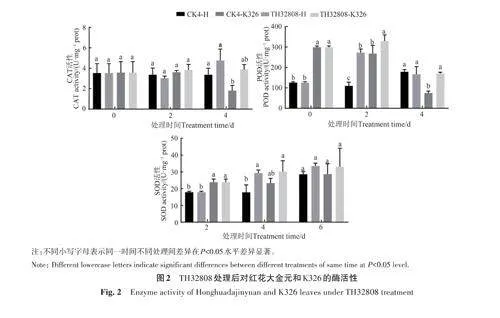
紅花大金元和K326 的過氧化氫酶(CAT)活性在接種TH32808的第4天時達到峰值,分別為4.75和3.87 U·mg?1 prot,是CK4的1.42與2.15倍;過氧化物酶(POD)在接種第2天達到峰值,分別為272.52 和329.15 U·mg?1 prot,是CK4 的2.49 和2.29 倍,在第4 天時活性降低;超氧化物歧化酶(SOD)在接種TH32808后活性呈不斷增加趨勢,在第4 天時為33.48 和32.95 U·mg?1 prot,是CK4的1.17和1.15倍(圖2)。
在接種煙草疫霉菌2 d后接種TH32808處理,紅花大金元與K326的抗病相關酶活性呈現的變化趨勢有些差別。紅花大金元的CAT與SOD活性在第6天最高,分別為4.81與28.43 U·mg?1 prot,是僅接種煙草疫霉菌處理的1.60與1.67倍,而POD無明顯變化規律;K326的POD和SOD在第6天活性最高,分別為702.93和 32.68 U·mg?1 prot,是僅接種煙草疫霉菌處理的1.05和1.20倍,而CAT無明顯變化規律(圖3)。
2.5 哈茨木霉32808 誘導煙草植株抗性基因的表達分析
由圖4可知,接種TH32808后,紅花大金元葉片中NPR1 基因的表達量在第1天時顯著上調,為CK5的2.03倍,第2天表達量有所降低,但仍顯著高于CK5,至第4天時仍高于CK5;PDF1.2 基因的表達量在第1天時顯著高于CK5,第2天繼續上調,為CK5的2.06倍,但在第3天開始低于CK5;EFE基因在第1天顯著上調表達,為CK5的10.31倍,隨后表達量逐漸降低,但均顯著高于CK5;NtSGT1 基因在前3 d的表達量呈逐漸增加趨勢,在第2和第3天顯著高于CK5,其中第3天為CK5的1.59倍。
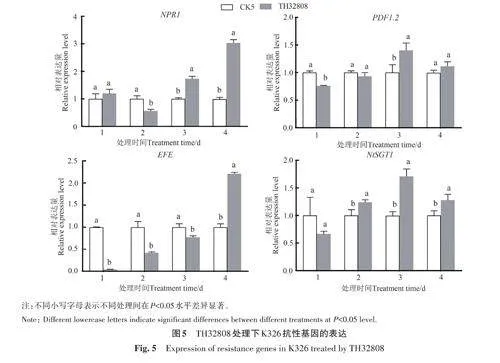
由圖5 可知,K326 經TH32808 處理后,葉片中抗病相關基因的表達與紅花大金元存在差異。NPR1 基因在第1和第2天的表達量低于CK5;在第3天時顯著高于CK5;在第4天時為CK5的3.04倍。PDF1.2 基因在第1 和第2 天的表達量低于CK5;在第3天時表達量顯著上調,為CK5的1.40倍;在第4 天時仍然高于CK5,但差異不顯著。EFE 基因的表達量呈逐漸上升趨勢,與紅花大金元趨勢相反,在第4天時表達量顯著上調,為CK5的2.21倍。NtSGT1 基因的表達量變化趨勢與紅花大金元相似,在第3 天時表達量顯著上調,為CK5的1.72倍;在第4天時表達量有所下降,但仍顯著高于CK5。
3 討論
本研究分析了哈茨木霉TH32808 對煙草的促生作用,并進一步探討了對煙草黑脛病害的生物防治及其誘導抗性機制,具有重要的理論意義和實踐應用價值。
木霉對植物的促生機制主要包括以下幾個方面。①產生植物生長素。生長素是一種可以促進植物生長的化學物質,Guzmán等[20]研究發現,在深綠木霉中有10個與生長素合成及轉運相關的基因;Vinale等[21]在木霉菌的次級代謝產物中分離出可促進番茄和油菜生長的類植物生長素6-PP,這類植物生長素還可以減少植物病害的發生。②提高植物養分利用率。唐磊等[22]證明,棘孢木霉可通過提高土壤中水解氮的比例,促進青蒿生長。哈茨木霉T-22可分解土壤中的復雜化合物,促進植物對養分的吸收利用,從而有利于植物生長發育[23]。③影響植物根際微生態。康萍芝等[24]研究表明,木霉制劑可顯著增加番茄根際土壤中益生菌的數量,從而促進番茄生長,同時降低枯萎病的發病率。本研究表明,哈茨木霉TH32808對2個供試品種煙草有一定促生作用,但效果不顯著,這可能與木霉的接種劑量有關,還需進一步深入研究。
木霉可通過競爭作用、重寄生、抗性作用和協同拮抗作用等機制來達到抑制和防治病害的作用。在木霉?植物?病原菌三者的互作關系中,木霉可誘導植物產生抗病性,對保證植物健康生長具有重要作用,其中誘導植物中防御酶活性增強是一種重要機制[25]。研究表明,煙草植株天然具備一定的抗性,且不同品種間存在抗性差異,如紅花大金元對煙草疫霉病的抗性明顯優于K326[26],而施用哈茨木霉可誘導煙草植株防御酶活性增強[12]。本研究表明,施用TH32808能夠顯著提高煙草葉片CAT、 POD和 SOD活性,增強煙草對病害的抵抗力,但不同煙草品種間存在差異;同時,在接種煙草疫霉菌2 d后再接種TH32808,煙草植株酶活性的變化趨勢會因煙草品種的不同出現差別。哈茨木霉定殖于煙草植株后會誘導植物抗性相關基因的表達。在本研究中,NPR1、PDF1.2、EFE 和NtSGT1 這4 個關鍵基因在TH32808 處理后第1~4天均出現上調表達趨勢。在K326品種中,水楊酸、茉莉酸和乙烯途徑的相關基因上調表達的時間比紅花大金元品種相對滯后,表明不同煙草品種在相同基因誘導表達中存在差異。綜上,哈茨木霉TH32808能顯著誘導2個供試品種植株中關鍵信號分子水楊酸、茉莉酸和乙烯的關鍵基因的表達,以此來增強煙草自身抵抗外界病害的能力,從而發揮對煙草黑脛病的防治作用。
