小粒咖啡葉綠素合成基因CaPOR的全基因組鑒定及表達(dá)分析
2024-12-31 00:00:00張振偉董相書(shū)楊婧李學(xué)俊杞美軍蔣快樂(lè)楊永林王步天施學(xué)東邱俊超陳治華葛宇
中國(guó)農(nóng)業(yè)科技導(dǎo)報(bào) 2024年10期



關(guān)鍵詞:小粒咖啡;葉綠素;生物信息學(xué)分析;表達(dá)分析
咖啡是茜草科(Rubiaceae)咖啡屬(Coffea)多年生常綠灌木或小喬木[1]。目前已被分類(lèi)的咖啡屬種類(lèi)約有124種,主要栽培于熱帶和亞熱帶地區(qū)[2]。咖啡不僅是熱帶發(fā)展中國(guó)家的重要經(jīng)濟(jì)作物,也是世界上最受歡迎的飲料和重要的商業(yè)商品之一[3]。商業(yè)栽培的咖啡主要有小粒咖啡(Coffea arabica)和中粒咖啡(Coffea canephora)。其中小粒咖啡又稱(chēng)阿拉比卡咖啡,是咖啡屬中唯一的異源四倍體(2n=4x=44),由中粒咖啡和丁香咖啡(Coffea eugenoedes)或其他生態(tài)型較近的二倍體雜交而來(lái),原產(chǎn)于埃塞俄比亞中部高原[3]。我國(guó)咖啡主要種植在云南和海南省,其中云南省以小粒咖啡為主,主要種植在普洱、德宏、保山、西雙版納和臨滄,這5個(gè)產(chǎn)區(qū)的種植面積占云南咖啡種植面積的80%以上[4]。葉綠素(Chlorophyll,Chl)是一種脂溶性色素,廣泛存在于植物體的綠色組織中,尤其在果實(shí)的綠色果皮和植物體的葉片中含量極高,在植物進(jìn)行光合作用的過(guò)程中有著極其重要的作用[5]。因此,明確小粒咖啡葉片葉綠素合成的關(guān)鍵基因及葉綠素合成的調(diào)控機(jī)理,對(duì)提高小粒咖啡的有機(jī)物積累及咖啡豆品質(zhì)有重要意義。
植物葉綠素的生物合成是十分復(fù)雜的過(guò)程,由多種酶和基因共同調(diào)控[6]。擬南芥(Arabidopsisthaliana)的葉綠素生物合成主要由17種酶和27種編碼基因調(diào)控,其中任何酶的基因突變或通過(guò)抑制劑抑制其活性均有可能導(dǎo)致Chl含量變化,從而影響光合效率甚至導(dǎo)致植株死亡[7]。葉綠素的合成過(guò)程共15步[8],根據(jù)高等植物葉綠素合成的整個(gè)途徑,又可將葉綠素的合成分為2大步:第1步是由L-谷氨酰-tRNA合成原卟啉Ⅸ,其中編碼基因HEMA、HEMB、GSA(HEML)、HEMC、HEMD、HEME、HEMF 和HEMG 已從擬南芥[6]和水稻(Oryzasativa)[9]中分離獲得;第2步由原卟啉Ⅸ合成葉綠素,相關(guān)催化基因有CHLD、CHLH、CHLI、CHLM、CRDI、DVR、POR、CHLG、CAO[5,10]。POR (protochlorophyllide oxidoreductase)主要參與葉綠素生物合成途徑中的倒數(shù)第2個(gè)反應(yīng),對(duì)光合作用至關(guān)重要,負(fù)責(zé)催化光合體中的光合色素前體原葉綠素酸轉(zhuǎn)化為葉綠素a,是光誘導(dǎo)被子植物轉(zhuǎn)綠的關(guān)鍵酶[11-13]。此外,在對(duì)葉綠素合成相關(guān)基因的研究中發(fā)現(xiàn),可以通過(guò)調(diào)節(jié)光照強(qiáng)度或者光照顏色來(lái)提高植物體中POR 基因的表達(dá)水平,從而促進(jìn)葉綠素合成[14-16],或者通過(guò)控制土壤、基質(zhì)中的水分含量來(lái)降低POR 基因的表達(dá),從而抑制葉綠素合成[17]。
在咖啡的生長(zhǎng)過(guò)程中,地理環(huán)境、氣候條件等因素均會(huì)對(duì)咖啡的生長(zhǎng)發(fā)育造成嚴(yán)重影響。但目前我國(guó)小粒咖啡的研究方向以生長(zhǎng)、生理為主,關(guān)于分子水平的研究較少[18]。本研究通過(guò)對(duì)小粒咖啡的葉片進(jìn)行轉(zhuǎn)錄組測(cè)序,通過(guò)生物信息學(xué)手段挖掘小粒咖啡葉片葉綠素合成的關(guān)鍵基因,并明確小粒咖啡生長(zhǎng)發(fā)育過(guò)程中葉綠素合成相關(guān)基因的表達(dá)模式,為小粒咖啡葉片葉綠素合成的機(jī)理研究提供參考。
1 材料與方法
1.1 小粒咖啡葉綠素合成相關(guān)基因家族成員篩選及編碼蛋白的理化性質(zhì)分析
從NCBI 數(shù)據(jù)庫(kù)(https://www. ncbi. nlm. nih.gov/)中下載小粒咖啡的參考蛋白序列以及基因組數(shù)據(jù),分別構(gòu)建本地?cái)?shù)據(jù)庫(kù)。通過(guò)TAIR 網(wǎng)站(https://www.arabidopsis.org/)搜集到27 個(gè)葉綠素合成相關(guān)蛋白序列,利用本地BLASTP程序以及TBtools[19]的blast程序分別對(duì)小粒咖啡蛋白序列進(jìn)行比對(duì)篩選,設(shè)置Elt;10-10,取2個(gè)數(shù)據(jù)集的并集,去除重復(fù)序列,將篩選出的序列作為小粒咖啡葉綠素合成相關(guān)基因家族序列。篩選出的小粒咖啡蛋白序列,利用在線(xiàn)工具Expasy(https://www.expasy.org/)分析小粒咖啡葉綠素合成相關(guān)基因家族成員蛋白質(zhì)的氨基酸數(shù)量、分子量、理論等電點(diǎn)、不穩(wěn)定系數(shù)、脂肪指數(shù)和親水平均系數(shù)等理化性質(zhì)指標(biāo)。
1.2 小粒咖啡葉綠素合成相關(guān)基因系統(tǒng)發(fā)生樹(shù)的構(gòu)建
為研究小粒咖啡葉綠素合成相關(guān)基因與其他物種之間的親緣關(guān)系,將小粒咖啡和擬南芥的葉綠素合成相關(guān)基因序列運(yùn)用MEGA7中muscle程序進(jìn)行多序列比對(duì),比對(duì)結(jié)果經(jīng)修剪后用NJ(neighbour-joining,)法構(gòu)建系統(tǒng)發(fā)生樹(shù),并用Chiplot(https://www.chiplot.online/)[20]網(wǎng)站對(duì)系統(tǒng)發(fā)生樹(shù)進(jìn)行美化。
1.3 不同品種小粒咖啡葉片葉綠素合成相關(guān)基因表達(dá)模式分析
分別選取20個(gè)品種的小粒咖啡成熟時(shí)期葉片為試驗(yàn)材料,提取RNA 樣品,采用IlluminaHiSeq進(jìn)行RNA-seq測(cè)序。測(cè)序結(jié)果與上一階段中所發(fā)掘的小粒咖啡葉片葉綠素合成相關(guān)基因進(jìn)行比對(duì)篩選,采用FPKM(fragments per kilobase ofexon model per million mapped fragments)法分析轉(zhuǎn)錄組數(shù)據(jù)中小粒咖啡葉片葉綠素合成相關(guān)基因的表達(dá),通常認(rèn)為FPKMgt;15 為高表達(dá)基因[21]。因此,通過(guò)對(duì)轉(zhuǎn)錄組數(shù)據(jù)中每個(gè)小粒咖啡品種基因的FPKM 求平均值,并選取至少有1 個(gè)品種FPKMgt;15的葉綠素合成相關(guān)基因,利用TBtools繪制表達(dá)量熱圖。
1.4 小粒咖啡CaPOR 基因保守基序、結(jié)構(gòu)域以及基因結(jié)構(gòu)分析
利用MEME(https://meme-suite. org/meme/tools/meme)在線(xiàn)分析軟件對(duì)小粒咖啡CaPOR 和擬南芥AtPOR 基因編碼蛋白的保守基序進(jìn)行分析,通過(guò)NCBI batch CD search Tool(https://www.ncbi. nlm. nih. gov/Structure/bwrpsb/bwrpsb. cgi)[22-24]在線(xiàn)軟件對(duì)小粒咖啡CaPOR 及擬南芥基因家族成員的蛋白結(jié)構(gòu)域進(jìn)行分析,利用TBtools構(gòu)建小粒咖啡以及擬南芥POR 基因家族成員保守基序、結(jié)構(gòu)域以及基因結(jié)構(gòu)可視化圖。
1.5 小粒咖啡CaPOR 基因順式作用元件分析
利用TBtools分離小粒咖啡CaPOR 基因起始密碼子上游2 kb序列,將分離的序列在PlantCARE(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)[25]上進(jìn)行順式作用元件分析,將無(wú)效信息剔除后利用TBtools進(jìn)行可視化處理。
1.6 小粒咖啡CaPOR 基因的染色體分布
利用TBtools 導(dǎo)入篩選出的38 個(gè)小粒咖啡CaPOR 基因組數(shù)據(jù)及注釋文件,繪制CaPOR 基因所在染色體的密度條帶,并定位CaPOR 基因的分布。
1.7 小粒咖啡CaPOR 基因的同源關(guān)系及同源性比對(duì)
利用從TAIR 網(wǎng)站(https://www. arabidopsis.org/)下載的擬南芥全基因組數(shù)據(jù)及注釋文件,NCBI 數(shù)據(jù)庫(kù)(https://www.ncbi.nlm.nih.gov/)下載小粒咖啡、中粒咖啡、丁香咖啡的全基因組數(shù)據(jù)及注釋文件。以小粒咖啡CaPOR 基因?yàn)閰⒄眨肨Btools 分析2個(gè)親本咖啡與小粒咖啡CaPOR 基因的共線(xiàn)性關(guān)系,并將結(jié)果可視化。
1.8 小粒咖啡CaPOR 基因密碼子偏好分析
通過(guò)生信云在線(xiàn)網(wǎng)站(https://ngs.sangon.com/cloudtool)分析密碼子第1、2、3位堿基的GC含量,記作GC1、GC2、GC3,并計(jì)算GC1 和GC2 的平均值,記作GC12。以GC3 為橫軸,GC12 為縱軸作圖[26]。
通過(guò)生信云在線(xiàn)網(wǎng)站計(jì)算有效密碼子數(shù)(effective number of codon,ENC),并根據(jù)ENC 和GC3進(jìn)行ENC-plot分析[27]。
通過(guò)MEGA 7計(jì)算密碼子第3位堿基的A、T、G、C含量,分別記作A3、T3、G3、C3,以G3/(G3+C3)為橫軸、A3/(A3+T3)為縱軸進(jìn)行偏倚分析[28]。
1.9 小粒咖啡葉片葉綠素含量測(cè)定
稱(chēng)取3份小粒咖啡葉片各0.2 g,剪碎放入研體,加少量石英砂和碳酸鈣及3 mL提取液(95%乙醇),研成勻漿,再加5 mL提取液,繼續(xù)研磨至組織變白,室溫靜置3 min后轉(zhuǎn)移到10 mL離心管中,4 000 r·min-1離心5 min,取上清液,用提取液定容至25 mL。以95%乙醇為空白,分別在波長(zhǎng)665、649和470 nm下測(cè)定吸光度。所得結(jié)果代入下列公式,計(jì)算單位鮮重組織中葉綠素a(Chl a)、葉綠素b(Chl b)、葉綠素(Chl)、類(lèi)胡蘿卜素(Car)和葉綠體色素含量。
1.10 小粒咖啡CaPOR 基因熒光定量PCR 驗(yàn)證
對(duì)同一品種分別采集3株不同植株的嫩葉與成熟葉片,設(shè)置3個(gè)生物學(xué)重復(fù)。分別將這2個(gè)時(shí)期的葉片液氮研磨后,置于1.5 mL離心管中,利用RNA快速提取試劑盒上提取RNA,將提取質(zhì)量合格的RNA置于-80 ℃保存?zhèn)溆谩?/p>
利用NCBI設(shè)計(jì)引物,并以24S[29]作為內(nèi)參基因,進(jìn)行熒光定量PCR,并進(jìn)行3次重復(fù)以確保數(shù)據(jù)的準(zhǔn)確性,結(jié)果通過(guò)2-△△CT法[30]計(jì)算基因的相對(duì)表達(dá)量,所用引物如表1所示。
2 結(jié)果與分析
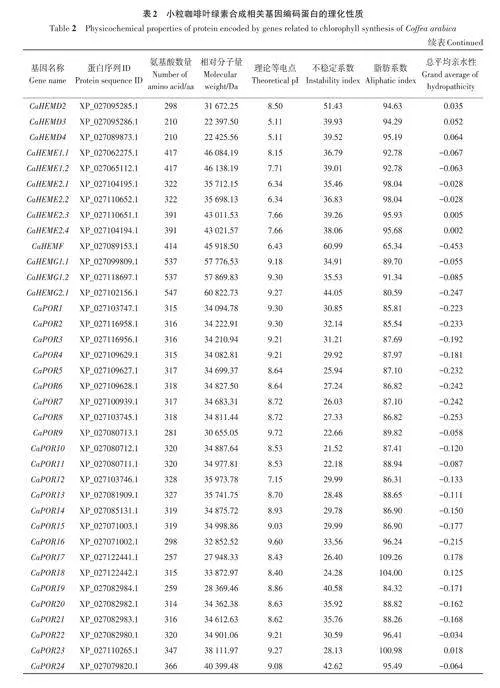
2.1 小粒咖啡葉綠素合成相關(guān)基因家族成員篩選及編碼蛋白的理化性質(zhì)分析
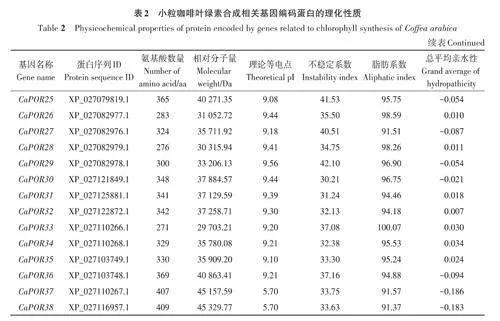
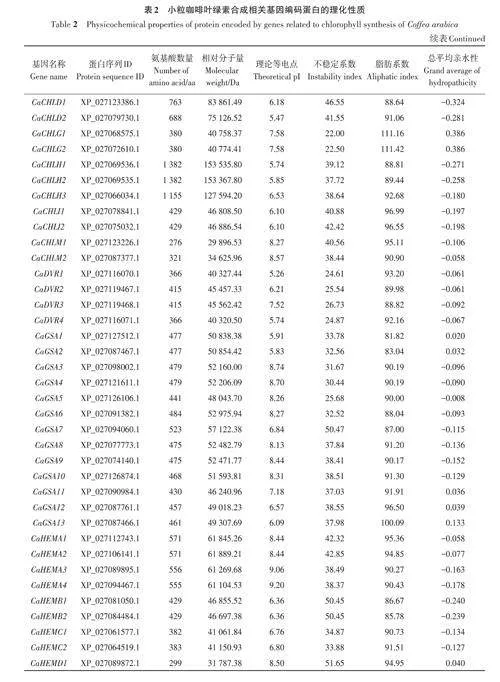
基于擬南芥葉綠素合成相關(guān)基因的27個(gè)參考序列,共篩選出小粒咖啡葉綠素合成相關(guān)基因92個(gè),依據(jù)比對(duì)重合度可分為17個(gè)基因家族。由表2可知,小粒咖啡葉綠素合成相關(guān)蛋白質(zhì)的氨基酸數(shù)量差異較大,其中CaHEMD3和CaHEMD4的氨基酸數(shù)量最少,只有210 個(gè),CaCHLH1 和CaCHLH2的氨基酸數(shù)量最多,均高達(dá)1 382個(gè),大多數(shù)蛋白質(zhì)的氨基酸數(shù)量在600以?xún)?nèi);相對(duì)分子量為22 397.5~153 535.84 Da;理論等電點(diǎn)為5.11~9.72,其中大于8的有58個(gè),呈堿性,小于6.5的有21個(gè),呈酸性;不穩(wěn)定系數(shù)為21.52~60.99,其中有70個(gè)蛋白質(zhì)的不穩(wěn)定系數(shù)小于40,具有較好的穩(wěn)定性,有22個(gè)蛋白質(zhì)的不穩(wěn)定系數(shù)大于40;脂肪系數(shù)為65.34~111.42,其中有85個(gè)蛋白質(zhì)的脂肪系數(shù)小于100,較親水;總平均親水性為-0.453~0.386,其中有23 個(gè)蛋白質(zhì)的總平均親水性為正值,表現(xiàn)為疏水蛋白,有69個(gè)蛋白質(zhì)的總平均親水性為負(fù)值,表現(xiàn)為親水蛋白。
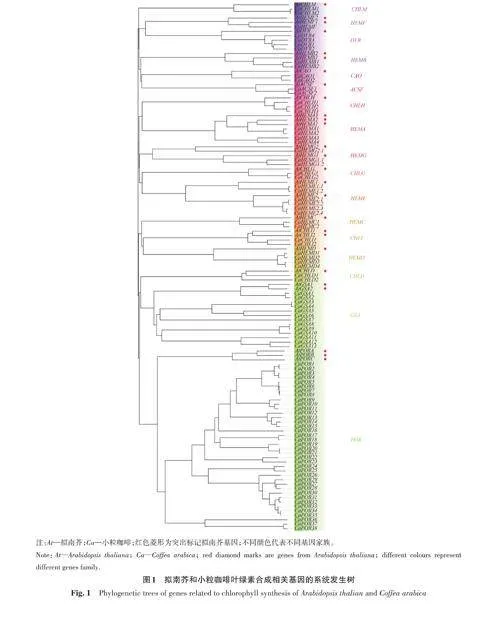
2.2 小粒咖啡葉綠素合成相關(guān)基因系統(tǒng)發(fā)生樹(shù)的分析
為探究小粒咖啡葉片葉綠素合成相關(guān)基因與擬南芥葉綠素合成相關(guān)基因的親緣關(guān)系,利用NJ法構(gòu)建系統(tǒng)發(fā)育樹(shù),如圖1所示。根據(jù)葉綠素合成途徑中起關(guān)鍵作用的基因,可將整個(gè)系統(tǒng)發(fā)育樹(shù)分為CHLM、HEMF、DVR、HEMB、CAO、ACSF、CHLH、HEMA、HEMG、CHLG、HEME、HEMC、CHLI、HEMD、CHLD、GSA、POR 共17 個(gè)分支。雖然每個(gè)小家族之間的親緣關(guān)系較遠(yuǎn),但每個(gè)小家族內(nèi)關(guān)系較近,且可信度較高,平均可信度都在80以上。由此表明,同一家族內(nèi)小粒咖啡與擬南芥的葉綠素合成相關(guān)基因具有極高的相似性。其中POR 基因家族成員較多,共38個(gè),占41.3%;其次是GSA 基因家族,共13個(gè),占14.1%;而其他葉綠素合成相關(guān)基因的數(shù)量較少,均不超過(guò)5個(gè)。
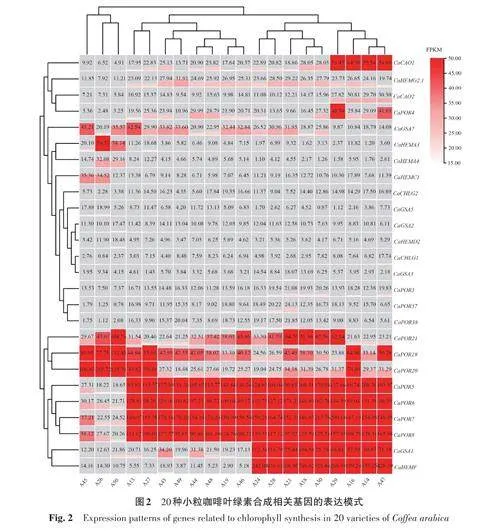
2.3 不同品種小粒咖啡葉綠素合成相關(guān)基因表達(dá)模式分析
分析20種小粒咖啡的成熟葉片中CaPOR 基因的表達(dá)模式,結(jié)果(圖2)表明,以FPKM≥15為篩選標(biāo)準(zhǔn),篩選出CaPOR5、CaPOR6、CaPOR7、CaPOR8、CaPOR19、CaPOR20、CaPOR21 共7個(gè)在小粒咖啡葉綠素合成中較穩(wěn)定表達(dá)的基因。其中CaPOR5、CaPOR6、CaPOR7、CaPOR8 基因在A(yíng)45、A26、A50中的表達(dá)量較低,而在其他17個(gè)小粒咖啡品種中表達(dá)量極高。這可能是由于這3個(gè)小粒咖啡品種中出現(xiàn)了新的變異,導(dǎo)致這4基因的表達(dá)被其他基因所替代,因此在這3個(gè)小粒咖啡品種中的表達(dá)量較低。
2.4 小粒咖啡CaPOR 基因保守基序、結(jié)構(gòu)域以及基因結(jié)構(gòu)分析
利用MEME在線(xiàn)分析軟件對(duì)小粒咖啡以及擬南芥POR蛋白序列進(jìn)行保守基序分析,結(jié)果(圖3)表明,共鑒定出6個(gè)保守基序。由圖4可知,小粒咖啡以及擬南芥的41個(gè)POR 基因的Motif種類(lèi)和分布較為相似,均具有motif 1、motif 2、motif 5,且這3 個(gè)motif 的順序也完全一致;只有CaPOR17 無(wú)motif 3,且其余成員的motif 3均位于開(kāi)頭位置。由此表明,擬南芥的和小粒咖啡的POR 基因家族成員的motif分布較為相似,具有較高的保守性。
進(jìn)一步分析小粒咖啡和擬南芥POR 基因的保守結(jié)構(gòu)域(圖4C)發(fā)現(xiàn),擬南芥以PLN00015(葉綠素還原酶)結(jié)構(gòu)域?yàn)橹鳎×?Х纫詒etinol-DH_like_SDR_c_like 結(jié)構(gòu)域和NADB_Rossmann superfamily結(jié)構(gòu)域?yàn)橹鳎渲挟?dāng)缺少motif 4時(shí), CaPOR 基因都具有retinol-DH_like_SDR_c_like結(jié)構(gòu)域。
分析小粒咖啡CaPOR 基因的內(nèi)含子/外顯子分布,結(jié)果(圖4D)表明,不同CaPOR 基因的內(nèi)含子和外顯子的數(shù)量及結(jié)構(gòu)存在差異。其中CaPOR12、CaPOR13、CaPOR14、CaPOR15 和CaPOR36 這5 個(gè)基因均只有1 個(gè)外顯子,而CaPOR36 只有1個(gè)內(nèi)含子。
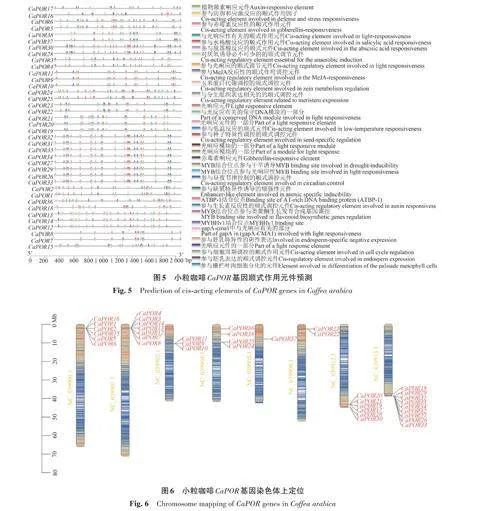
2.5 小粒咖啡CaPOR 基因順式作用元件分析
對(duì)小粒咖啡CaPOR 基因上游2 kb區(qū)域的啟動(dòng)子區(qū)進(jìn)行分析,得到33種共1 025個(gè)具有特異性功能的順式作用元件(圖5)。其中與生長(zhǎng)發(fā)育相關(guān)的元件種類(lèi)有11種,共250個(gè),占24.39%;與光反應(yīng)相關(guān)的元件有10種,共464個(gè),占45.27%;;激素響應(yīng)類(lèi)元件有10種,共154個(gè),占15.02%;生物與非生物脅迫類(lèi)元件有6 種,共157 個(gè),占15.32%。表明小粒咖啡CaPOR 基因大多參與光響應(yīng),同時(shí)也與植物的生長(zhǎng)發(fā)育過(guò)程密切相關(guān)。
2.6 小粒咖啡CaPOR 基因的染色體分布
對(duì)小粒咖啡CaPOR 基因進(jìn)行染色體定位,有38個(gè)CaPOR 基因定位于8條染色體上,其中染色體NC_039913.1上定位到的CaPOR 基因最多,有9 個(gè);染色體NC_039905.1 上的基因最少,只有CaPOR37(圖6)。CaPOR 基因多聚集在染色體的末端,有利于穩(wěn)定遺傳,且同一染色體上的CaPOR 基因位置較為相近,可能出現(xiàn)了復(fù)制的現(xiàn)象。
2.7 小粒咖啡CaPOR 基因的同源關(guān)系及同源性比對(duì)
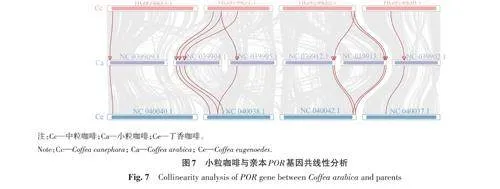
對(duì)小粒咖啡CaPOR 基因與2個(gè)親本(中粒咖啡和丁香咖啡)進(jìn)行共線(xiàn)性分析,結(jié)果(圖7)表明,在小粒咖啡中分布于6 條染色體上的9 個(gè)CaPOR 基因與中粒咖啡的8個(gè)基因存在共線(xiàn)性關(guān)系;有10個(gè)CaPOR 基因與丁香咖啡的10個(gè)基因存在共線(xiàn)性關(guān)系;在中粒咖啡和丁香咖啡中,有1個(gè)POR 基因與小粒咖啡的2個(gè)CaPOR 基因具有共線(xiàn)性關(guān)系,且這2個(gè)基因分別位于2條相鄰染色體的末端。由此推斷,小粒咖啡在進(jìn)化過(guò)程中,CaPOR 基因可能發(fā)生復(fù)制現(xiàn)象,且與親本具有共線(xiàn)性關(guān)系的CaPOR 基因具有較保守的結(jié)構(gòu)。
2.8 小粒咖啡CaPOR 基因密碼子偏好分析
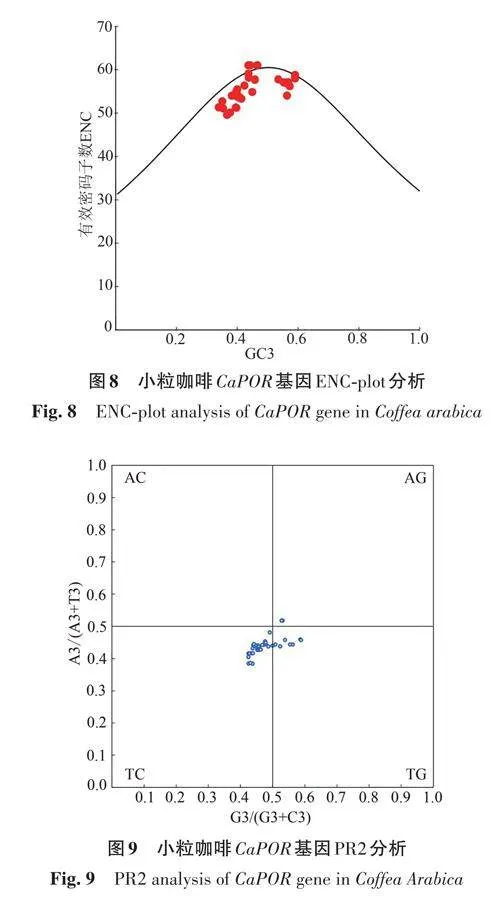
ENC-plot 分析(圖8)顯示,小粒咖啡的CaPOR 基因密碼子大部分沿標(biāo)準(zhǔn)曲線(xiàn)分布,也有部分基因距標(biāo)準(zhǔn)曲線(xiàn)較遠(yuǎn),說(shuō)明CaPOR 基因密碼子偏好性主要受堿基突變的影響,部分基因受自然選擇的影響。
PR2-plot 偏倚分析(圖9)顯示,小粒咖啡CaPOR 基因大部分位于左下角,表明小粒咖啡CaPOR 基因密碼子的第3位堿基中T、C的頻率高于A(yíng)、G;且大多數(shù)基因偏離中心,說(shuō)明小粒咖啡CaPOR基因受堿基突變、自然選擇等多種因素共同影響。
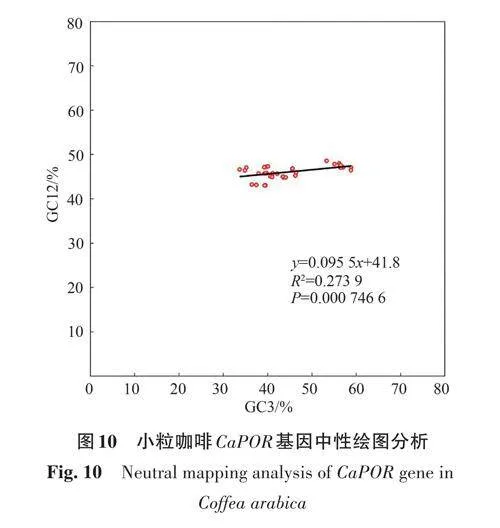
中性繪圖分析(圖10)顯示,小粒咖啡CaPOR基因密碼子的GC12 為43.04%~48.57%,GC3 為33.69%~58.82%,GC12 與GC3 的相關(guān)系數(shù)為0.273 9,斜率為0.095 5,Plt;0.05,二者間顯著相關(guān),表明小粒咖啡CaPOR 基因密碼子使用偏好性主要受堿基突變影響。
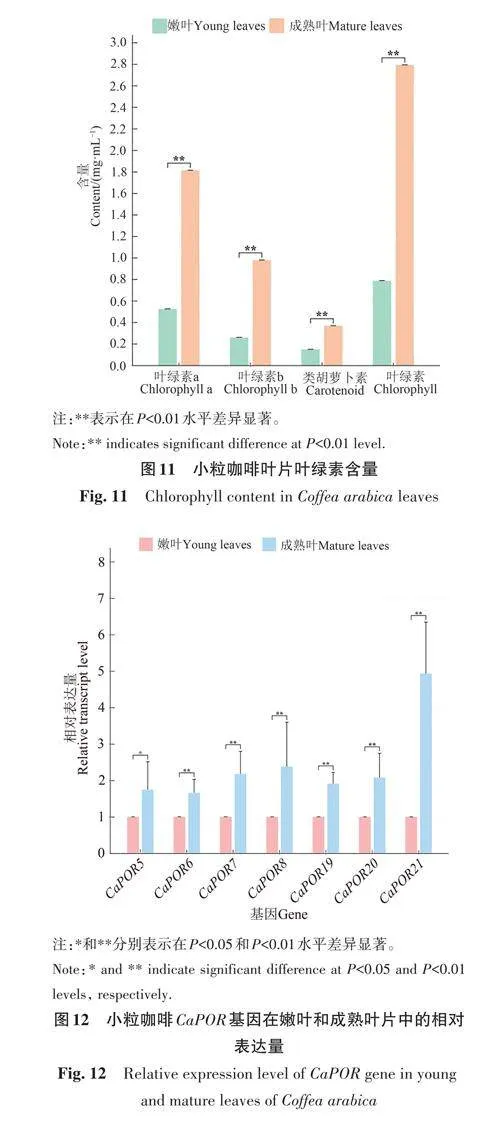
2.9 小粒咖啡葉片葉綠素含量分析
對(duì)小粒咖啡嫩葉期和成熟期葉片的葉綠素a、葉綠素b 和類(lèi)胡蘿卜素含量進(jìn)行測(cè)定,結(jié)果(圖11)表明,成熟葉片的葉綠素a、葉綠素b和類(lèi)胡蘿卜素含量均極顯著高于嫩葉。在嫩葉和成熟葉中均表現(xiàn)為葉綠素含量較高,類(lèi)胡蘿卜素含量較低;在葉綠素中又主要以葉綠素a 為主,葉綠素b含量較低。
2.10 小粒咖啡CaPOR 基因的表達(dá)分析
進(jìn)一步對(duì)7個(gè)在成熟葉片中具有極高的表達(dá)量的CaPOR 基因進(jìn)行實(shí)時(shí)熒光定量PCR驗(yàn)證,結(jié)果(圖12)表明,這7個(gè)基因在成熟葉片中的表達(dá)量顯著或極顯著高于嫩葉,其中CaPOR5、CaPOR6、CaPOR7、CaPOR8、CaPOR19、CaPOR20基因在成熟葉片中的表達(dá)量均約為嫩葉的2倍,CaPOR21 基因在成熟葉片中的表達(dá)量約為嫩葉的5倍。
3 討論
POR是葉綠素生物合成的關(guān)鍵酶,可催化原葉綠素酸脂在光照條件下還原為葉綠素酸酯[31]。當(dāng)植物體中缺乏某一葉綠素時(shí),其POR 基因表達(dá)水平也會(huì)下調(diào),這表明植物體中葉綠素含量與POR 基因的表達(dá)息息相關(guān)[32]。本研究利用生物信息學(xué)以擬南芥葉綠素合成相關(guān)的27個(gè)基因?yàn)閰⒄眨埠Y選出小粒咖啡葉綠素合成相關(guān)基因92個(gè)。對(duì)小粒咖啡多個(gè)品種的成熟葉片進(jìn)行轉(zhuǎn)錄組測(cè)序,結(jié)合轉(zhuǎn)錄組數(shù)據(jù)發(fā)現(xiàn),大多數(shù)小粒咖啡CaPOR 基因在成熟葉片中不表達(dá)或是表達(dá)量較低,其中CaPOR5、CaPOR6、CaPOR7、CaPOR8、CaPOR19、CaPOR20、CaPOR21 基因的FPKM 值均大于15,推斷這7個(gè)基因可能對(duì)小粒咖啡葉片的葉綠素合成起重要作用,且這7個(gè)基因均包含retinol-DH_like_SDR_c_like 結(jié)構(gòu)域,其蛋白氨基酸數(shù)量為310~320,多為堿性、穩(wěn)定性較好的親水蛋白,僅CaPOR19 基因編碼蛋白的氨基酸數(shù)量為259,且不穩(wěn)定系數(shù)大于40,表現(xiàn)為較不穩(wěn)定。研究表明,POR 基因在蘿卜(Raphanussativus)[33]、水稻[34]、安吉白茶(Camellia sinensis)[35]和番茄(Solanum lycopersicum)[36]的葉綠素合成過(guò)程中起重要作用。本研究也發(fā)現(xiàn),這7個(gè)CaPOR基因在小粒咖啡葉片不同時(shí)期的表達(dá)量具有顯著差異,在小粒咖啡葉片發(fā)育過(guò)程中,均表現(xiàn)為后期的表達(dá)量顯著高于前期,與成熟葉片的色素含量顯著高于嫩葉相一致。由此表明,這7個(gè)CaPOR基因在小粒咖啡葉片的生長(zhǎng)發(fā)育過(guò)程中與葉綠素合成關(guān)系密切,可能是負(fù)責(zé)小粒咖啡葉片葉綠素合成的關(guān)鍵基因。其他CaPOR 基因在小粒咖啡葉片的生長(zhǎng)發(fā)育過(guò)程中表達(dá)量較低或是不表達(dá),可能參與葉片生長(zhǎng)的其他生物功能過(guò)程。
本研究進(jìn)一步分析了CaPOR 基因的保守基序、結(jié)構(gòu)域、順式作用元件等,發(fā)現(xiàn)小粒咖啡CaPOR 基因在結(jié)構(gòu)上具有極高的相似性,在染色體上聚集成簇分布,推測(cè)可能具有相似的功能或表達(dá)模式[37]。小粒咖啡CaPOR 基因與2個(gè)親本間存在共線(xiàn)性關(guān)系,親本的1個(gè)基因?qū)?yīng)小粒咖啡2條染色體上的2個(gè)基因,推測(cè)小粒咖啡CaPOR 基因在遺傳過(guò)程中出現(xiàn)復(fù)制現(xiàn)象,重復(fù)基因有可能導(dǎo)致基因冗余或出現(xiàn)新的功能[38]。CaPOR 基因密碼子偏好性分析發(fā)現(xiàn),主要受堿基突變的影響,同時(shí)也受自然選擇的影響[3940]。
