產(chǎn)褐黃色鏈霉菌HEBRC45958 菌株防控番茄棒孢葉斑病研究
2024-12-31 00:00:00黃大野余志斌萬(wàn)中義楊丹李金萍曹春霞
中國(guó)農(nóng)業(yè)科技導(dǎo)報(bào) 2024年11期





摘要:為明確產(chǎn)褐黃色鏈霉菌HEBRC45958菌株對(duì)番茄棒孢葉斑病的防控效果,采用平板對(duì)峙培養(yǎng)結(jié)合光學(xué)顯微鏡和掃描電鏡觀察確定HEBRC45958離體抑菌活性,并測(cè)定其產(chǎn)鐵載體能力和水解酶活性。同時(shí)采用盆栽試驗(yàn)確定HEBRC45958發(fā)酵液對(duì)番茄棒孢葉斑病防控效果。結(jié)果表明,對(duì)峙培養(yǎng)條件下 HEBRC45958對(duì)多主棒孢的抑菌率為52.65%。掃描電鏡下觀察,與對(duì)照相比,HEBRC45958處理后多主棒孢菌絲發(fā)生扭曲和破裂,透射電鏡下觀察,多主棒孢細(xì)胞膜和細(xì)胞壁距離增加。HEBRC45958菌株能產(chǎn)生淀粉酶、纖維素酶、幾丁質(zhì)酶和嗜鐵素。盆栽試驗(yàn)結(jié)果表明,HEBRC45958發(fā)酵液對(duì)番茄棒孢葉斑病的防效為100%,具有較好的生防應(yīng)用前景。
關(guān)鍵詞:產(chǎn)褐黃色鏈霉菌;番茄;多主棒孢;防效
doi:10.13304/j.nykjdb.2023.0416
中圖分類號(hào):S476 文獻(xiàn)標(biāo)志碼:A 文章編號(hào):10080864(2024)11013607
由多主棒孢(Corynespora cassiicola)引起的番茄棒孢葉斑病是番茄的一種重要病害,其寄主廣泛,可侵染380個(gè)屬內(nèi)的500多種植物,包括黃瓜、辣椒、橡膠和一品紅等經(jīng)濟(jì)作物[1]。該病害最早在美國(guó)佛羅里達(dá)地區(qū)爆發(fā),隨后在日本和韓國(guó)等地也相繼發(fā)生。由于其寄主廣泛、侵染力強(qiáng)、傳播途徑多樣,加之近些年我國(guó)設(shè)施蔬菜面積不斷擴(kuò)大,高溫高濕等環(huán)境為番茄棒孢葉斑病的發(fā)生提供了條件,在我國(guó)山東、海南和北京等地區(qū)大面積發(fā)生,嚴(yán)重影響了番茄生產(chǎn)[23]。目前棒孢葉斑病的防控主要依靠化學(xué)殺菌劑,然而,隨著殺菌劑的大量使用,多主棒孢出現(xiàn)了嚴(yán)重的抗藥性,如對(duì)多菌靈、啶酰菌胺和甲氧基丙烯酸酯類殺菌劑均產(chǎn)生嚴(yán)重的抗藥性[4-8];另外,化學(xué)殺菌劑的大量使用會(huì)造成農(nóng)藥殘留和環(huán)境污染,亟需更為安全有效的防控手段。
由于生物防控更安全,在作物病害防控中得到了廣泛應(yīng)用。研究表明,貝萊斯芽孢桿菌(Bacillus velezensis)ZF2 菌株和解淀粉芽孢桿菌(Bacillus amyloliquefaciens)ZF57菌株對(duì)黃瓜棒孢葉斑病具有良好的防控效果[910];枯草芽孢桿菌(Bacillus subtilis)NBF809菌株和死亡谷芽孢桿菌(Bacillus vallismortis)NBIF001菌株對(duì)番茄棒孢葉斑病具有良好的防控效果[1112]。前人關(guān)于棒孢葉斑病的生物防控研究主要集中于芽孢桿菌,而對(duì)鏈霉菌(Streptomycete)防控棒孢葉斑病的報(bào)道較少。鏈霉菌作為重要的生防微生物,可以產(chǎn)生多種抗生素、植物激素、水解酶等抑菌活性物質(zhì),對(duì)防控病害和提高植物抗病性具有重要作用,是創(chuàng)制新型生物農(nóng)藥的重要資源。本研究從番茄根際土壤中分離出一株對(duì)多種植物病原真菌具有良好抑菌效果的產(chǎn)褐黃色鏈霉菌(Streptomycesphaeoluteichromatogenes)HEBRC45958,菌種保藏號(hào)為CCTCC NO:M2015768,進(jìn)一步分析其對(duì)番茄棒孢葉斑病的防控效果和抑菌機(jī)理,以期為生物農(nóng)藥的開(kāi)發(fā)與應(yīng)用奠定基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料
供試菌株為多主棒孢(Corynespora cassiicola)和產(chǎn)褐黃色鏈霉菌(S. phaeoluteichromatogenes)HEBRC45958,均由湖北省生物農(nóng)藥工程研究中心分離和保藏。供試藥劑為50%啶酰菌胺水分散粒劑,由巴斯夫歐洲公司生產(chǎn)。供試番茄品種為‘中雜109’,由中蔬種業(yè)科技(北京)有限公司提供。
1.2 試驗(yàn)方法
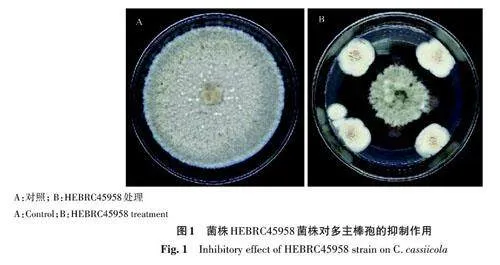
1.2.1 HEBRC45958菌株抑菌譜測(cè)定 HEBRC45958菌株從高氏1號(hào)斜面活化至高氏1號(hào)平板,瓜果腐霉(Pythium aphanidermatum) 、辣椒疫霉(Phytophthora capsici) 、灰葡萄孢(Botrytiscinerea)、多主棒孢(C. cassiicola)、尖孢鐮刀菌(Fusarium oxysporum)、膠孢炭疽菌(Colletotrichumgloeosporioides)和核盤菌(Sclerotinia sclerotiorum)共7 種植物病原菌由湖北省生物農(nóng)藥工程研究中心分離保藏,從馬鈴薯葡萄糖瓊脂(potatodextrose agar,PDA)斜面活化至PDA 平板備用。用直徑4 mm 打孔器打取HEBRC45958 菌餅,置于PDA 平板上、下、左、右4 個(gè)點(diǎn),距離中心約30 mm,48 h后打取病原菌菌餅置于平板中央,以不加HEBRC45958 菌株作為對(duì)照,每個(gè)處理設(shè)3個(gè)重復(fù),待對(duì)照菌落長(zhǎng)滿培養(yǎng)皿時(shí)測(cè)量菌落直徑,并計(jì)算抑菌率,公式如下。
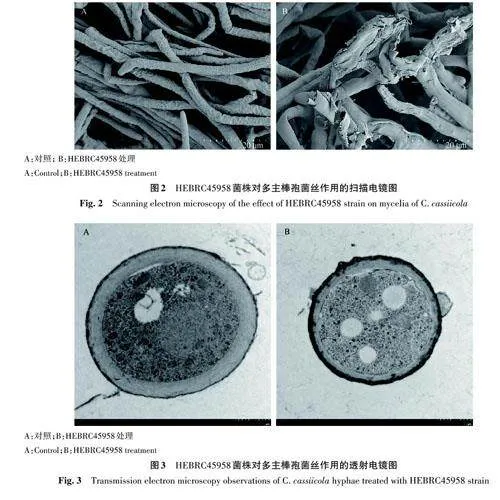
1.2.2 HEBRC45958菌株對(duì)多主棒孢菌絲作用觀察 從對(duì)峙培養(yǎng)的平板抑菌帶邊緣和對(duì)照菌落邊緣挑取多主棒孢菌絲,掃描電鏡和透射電鏡菌絲處理參考Li等[13]的方法,觀察菌絲形態(tài)和細(xì)胞結(jié)構(gòu)的變化。
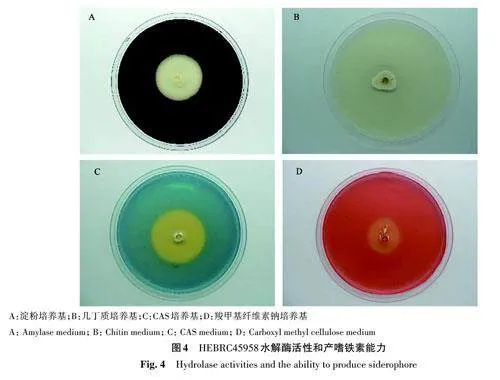
1.2.3 產(chǎn)嗜鐵素和水解酶活性測(cè)定 產(chǎn)嗜鐵素活性測(cè)定:使用打孔器打取HEBRC45958菌餅接種于CAS培養(yǎng)基平板,28 ℃培養(yǎng)箱培養(yǎng)7 d后觀察是否產(chǎn)生黃色暈圈[14]。產(chǎn)水解酶活性測(cè)定:將HEBRC45958菌株分別接種于幾丁質(zhì)、酪蛋白、羧甲基纖維素鈉(carboxyl methyl cellulose,CMC)、淀粉和茯苓粉培養(yǎng)基平板,28 ℃培養(yǎng)箱培養(yǎng)7 d后觀察是否產(chǎn)透明圈[1516]。其中,酪蛋白平板需在培養(yǎng)后加入10%三氯乙酸;CMC平板需加入1 g·L-1剛果紅染1 h后,倒掉染液,再用1 mol·L-1 NaCl溶液浸泡1 h;淀粉平板需加入2.5% 盧氏碘液,浸泡15 min后倒掉,用無(wú)菌水沖洗。
1.2.4 HEBRC45958菌株對(duì)番茄棒孢葉斑病的防效測(cè)定 將番茄種子催芽后播種于32孔穴盤中,溫室常規(guī)管理30 d。按照Miyamoto等[17]方法制備多主棒孢病原菌孢子。將斜面病原菌活化至PDA平板,在25 ℃黑暗培養(yǎng)10 d,用無(wú)菌手術(shù)刀片刮去表面氣生菌絲,隨后在BLB(black lightblue)燈下繼續(xù)培養(yǎng)3 d使其產(chǎn)孢。刷取孢子并用無(wú)菌水調(diào)至104 CFU·mL-1備用。
將HEBRC45958菌株孢子從斜面培養(yǎng)基劃線至高氏1號(hào)平板培養(yǎng),28 ℃培養(yǎng)箱培養(yǎng)7 d后用4 mm打孔器打取菌餅至ISP2液體培養(yǎng)基,500 mL三角瓶裝入100 mL ISP2 液體,搖床180 r·min-1,28 ℃振蕩培養(yǎng)5 d作為種子液,隨后按5%體積接入ISP2 液體培養(yǎng)基,500 mL 三角瓶裝入100 mLISP2液體,搖床180 r·min-1,28 ℃振蕩培養(yǎng)7 d備用。將發(fā)酵液原液及25和50倍稀釋液均勻噴施番茄葉面至飽和,以有效成分333.3 μg·mL-1啶酰菌胺作為對(duì)照藥劑,無(wú)菌水作為空白對(duì)照。每處理3次重復(fù),每重復(fù)16棵番茄。24 h后噴霧接種病原菌孢子懸浮液,28 ℃保濕培養(yǎng)4 d后統(tǒng)計(jì)各處理病情指數(shù)并計(jì)算防效。分級(jí)標(biāo)準(zhǔn)按照Ishii等[18]方法。
1.3 數(shù)據(jù)分析
使用SPSS 13.0軟件進(jìn)行試驗(yàn)數(shù)據(jù)的整理及差異顯著性分析。
2 結(jié)果與分析
2.1 HEBRC45958 菌株對(duì)植物病原菌離體抑菌效果
在供試的幾種病原菌中,HEBRC45958菌株對(duì)灰葡萄孢、多主棒孢和核盤菌的抑菌活性較好,抑菌率分別達(dá)到59.63%、52.65%和53.33%;對(duì)辣椒疫霉及尖孢鐮刀菌無(wú)抑菌活性(表1)。HEBRC45958菌株對(duì)峙的多主棒孢菌落邊緣氣生菌絲稀疏(圖1)。
2.2 HEBRC45958 菌株對(duì)多主棒孢菌絲作用分析
在掃描電鏡下觀察,與對(duì)照相比,經(jīng)HEBRC45958處理后多主棒孢菌絲出現(xiàn)扭曲和破裂(圖2)。透射電鏡下觀察,與對(duì)照相比,經(jīng)HEBRC45958 處理后多主棒孢菌細(xì)胞壁外層加厚,細(xì)胞壁和細(xì)胞膜間距變大(圖3)。
2.3 HEBRC45958 菌株的嗜鐵素和水解酶活性分析
對(duì)HEBRC45958菌株分泌的嗜鐵素和5種細(xì)胞水解酶活性進(jìn)行測(cè)定,結(jié)果(圖4)表明,HEBRC45958菌株能夠在淀粉、幾丁質(zhì)、茯苓粉和CMC平板上產(chǎn)生透明圈,說(shuō)明該菌株能產(chǎn)生淀粉酶、幾丁質(zhì)酶、和纖維素酶;且能夠在CAS平板產(chǎn)生黃色暈圈,說(shuō)明菌株能夠產(chǎn)生嗜鐵素。
2.4 HEBRC45958 菌株發(fā)酵液防控番茄棒孢葉斑病
盆栽試驗(yàn)(表2和圖2)表明,HEBRC45958菌株發(fā)酵液對(duì)番茄棒孢葉斑病具有良好的防控效果,經(jīng)HEBRC45958菌株發(fā)酵液原液及25和50倍稀釋液處理后,番茄病情指數(shù)顯著降低,由對(duì)照的96.88分別降為0.00、6.90和11.11。HEBRC45958菌株發(fā)酵液原液及25和50倍稀釋液的防效分別為100.00%、92.88% 和88.53%,均高于對(duì)照藥劑啶酰菌胺。
3 討論
關(guān)于棒孢葉斑病的生物防控研究多集中于芽孢桿菌,而鏈霉菌的報(bào)道較少。金麗穎[19]報(bào)道了Streptomyces inhibens NEAU-D10 發(fā)酵液對(duì)黃瓜棒孢葉斑病具有良好的防控效果,本研究首次報(bào)道了S. phaeoluteichromatogenes HEBRC45958防控棒孢葉斑病。
經(jīng)HEBRC45958處理后,在掃描電鏡下觀察,多主棒孢的菌絲出現(xiàn)扭曲和破裂;透射電鏡下觀察,細(xì)胞膜和細(xì)胞壁間距離變大。由此推斷破壞細(xì)胞壁結(jié)構(gòu)可能是HEBRC45958 的一種抑菌機(jī)理。這與Xiong 等[20] 報(bào)道Streptomyces padanusJAU4234 的代謝產(chǎn)物多烯大環(huán)內(nèi)酯抗生素Antifungalmycin 702對(duì)立枯絲核菌菌絲作用的結(jié)果相似。
HEBRC45958能產(chǎn)生嗜鐵素、淀粉酶、幾丁質(zhì)酶和纖維素酶等抑菌物質(zhì)。其中淀粉酶、幾丁質(zhì)酶和纖維素酶作為細(xì)胞壁水解酶,能夠水解病原菌細(xì)胞壁[2021],該結(jié)果與掃描電鏡觀察結(jié)果一致。Fe3+是微生物生長(zhǎng)的關(guān)鍵因子,嗜鐵素具有與病原菌競(jìng)爭(zhēng)Fe3+ 能力,從而達(dá)到抑制病原菌生長(zhǎng)[2223]。因此HEBRC45958菌株代謝產(chǎn)生嗜鐵素也是防控棒孢葉斑病的一種抑菌機(jī)制。
鏈霉菌能產(chǎn)生豐富的次級(jí)代謝產(chǎn)物,除了水解酶外,還能產(chǎn)生多種類型的抗生素。Ma等[24]報(bào)道,不吸水鏈霉菌梧州亞種(Streptomyces ahygroscopicussubsp. wuzhouensis)11371菌株能產(chǎn)生的一種26環(huán)四烯大環(huán)內(nèi)酯類抗生素四霉素,其對(duì)黃瓜棒孢葉斑病盆栽防效達(dá)到79.70%。HEBRC45958 菌株產(chǎn)生的抗生素種類還需要進(jìn)一步鑒定。本研究通過(guò)盆栽試驗(yàn)評(píng)價(jià)了HEBRC45958菌株對(duì)多主棒孢的防效,后續(xù)需要進(jìn)一步進(jìn)行田間防控效果的評(píng)價(jià)。
參考文獻(xiàn)
[1] 李寶聚,高葦,石延霞,等.多主棒孢和棒孢葉斑病的研究進(jìn)
展[J].植物保護(hù)學(xué)報(bào),2012,39(2):171-176.
LI B J, GAO W, SHI Y X, et al .. Progress in researches on
Corynespora leaf spot [J]. Acta Phytophylacica Sin., 2012, 39(2):
171-176.
[2] 田守波,朱為民,朱龍英,等.番茄棒孢葉斑病病原鑒定及生
物學(xué)特性研究[J].植物保護(hù),2020, 46(3):157-162.
TIAN S B, ZHU W M, ZHU L Y, et al .. Identification of the
pathogen from Corynespora leaf spot of tomato and research on
its biological characteristics [J]. Plant Prot., 2020, 46(3):
157-162.
[3] 李明遠(yuǎn).李明遠(yuǎn)斷病手跡(六十一)北京發(fā)現(xiàn)番茄棒孢葉斑
病[J].農(nóng)業(yè)工程技術(shù),2015(10):84-85.
[4] DUAN Y B, XIN W J, LU F, et al.. Benzimidazole- and QoIresistance
in Corynespora cassiicola populations from greenhousecultivated
cucumber: an emerging problem in China [J].
Pesticide Biochem. Physiol., 2019, 153: 95-105.
[5] ZHU F D, SHI Y X, XIE X W, et al .. Occurrence distribution
and characteristics of boscalid-resistant Corynespora cassiicola
in China [J]. Plant Dis., 2019, 103(1) :69-76.
[6] SHI Y, ZHU F, SUN B, et al .. Two adjacent mutations in the
conserved domain of SdhB confer various resistance
phenotypes to fluopyram in Corynespora cassiicola [J]. Pest
Manage. Sci., 2021, 77(9):3980-3989.
[7] SHI Y, SUN B, XIE X, et al .. Site-directed mutagenesis of the
succinate dehydrogenase subunits B and D from Corynespora
cassiicola reveals different fitness costs and sensitivities to
succinate dehydrogenase inhibitors [J]. Environ. Microbiol., 2021,
23(10):5769-5783.
[8] SUN B X, ZHU G X, XIE X W, et al.. Double mutations on
succinate dehydrogenase are involved in SDHI resistance in
Corynespora cassiicola [J/OL]. Microorganisms, 2022, 10(1): 132
[2023-04-30]. https://doi.org/10.3390/microorganisms10010132.
[9] 趙昱榕,李磊,謝學(xué)文,等.貝萊斯芽胞桿菌ZF2對(duì)多主棒孢
病菌防治效果[J].中國(guó)生物防治學(xué)報(bào),2019,35(2):217-225.
ZHAO Y R, LI L, XIE X W, et al .. Biocontrol effect of Bacillus
velezensis strain ZF2 against Corynespora cassiicola [J]. J. Chin.
Biol. Control., 2019, 35(2):217-225.
[10] 李新宇,李磊,石延霞,等.黃瓜棒孢葉斑病拮抗細(xì)菌的篩選,
鑒定及防治效果[J].植物保護(hù)學(xué)報(bào),2020,47(3):620-627.
LI X Y, LI L, SHI Y X, et al .. Screening, identification and
control effects of antagonistic bacteria against cucumber
Corynespora leaf spot [J]. J. Plant Prot., 2020, 47(3):620-627.
[11] 黃大野,賁海燕,曹春霞,等.死亡谷芽胞桿菌NBIF-001防治
番茄棒孢葉斑病研究[J].植物保護(hù),2017,43(6):192-195.
HUANG D Y, BEN H Y, CAO C X, et al .. Control effect of
Bacillus vallismortis NBIF001 on Corynespora leaf spot on
tomato [J]. Plant Prot., 2017, 43(6):192-195.
[12] 黃大野,曹春霞,張亞妮,等.枯草芽孢桿菌NBF809防治番茄
棒孢葉斑病研究[J].中國(guó)蔬菜, 2018(12):40-44.
HUANG D Y, CAO C X, ZHANG Y N, et al .. Studies on
controlling tomato Corynespora leaf spot by Bacillus subtilis
NBF809 [J]. China Veget., 2018(12):40-44.
[13] LI Q, NING P, ZHENG L, et al .. Effects of volatile substances
of Streptomyces globisporus JK-1 on control of Botrytis cinerea
on tomato fruit [J]. Biol. Cont., 2012, 61(2):113-120.
[14] 王超,申成美,鄭麗,等.煙草青枯病生防細(xì)菌的篩選與生防
效果研究[J].植物保護(hù),2014,40(2):43-47.
[15] 武志江,王亞軍,楊柳,等.百合枯萎病拮抗細(xì)菌的篩選,鑒定
及其抑菌物質(zhì)研究[J].微生物學(xué)通報(bào),2015,42(7):1307-1320.
WU Z J, WANG Y J, YANG L, et al .. Isolation and
identification of an antagonistic bacterium against Fusarium on
lily and its antifungal substances [J]. Microbiol. China, 2015,
42(7):1307-1320.
[16] MUDYTWA R M, CHAIBVA P, TAKAWIRA M, et al ..
Evaluation of Trichoderma harzianum in controlling dampingoff
(Pythium spp.) on tomato (Solanum lycopersicum) seedling
varieties [J]. Ann. Biolo. Res., 2016, 7(6):6-11.
[17] MIYAMOTO T, ISHII H, SEKOT, et al.. Occurrence of
Corynespora cassiicola isolates resistant to boscalid on cucumber in
Ibaraki Prefecture, Japan [J]. Plant Pathol., 2009, 58(6):1144-
1151.
[18] ISHII H, YANO K, DATE H, et al .. Molecular characterization
and diagnosis of QoI resistance in cucumber and eggplant
fungal pathogens [J]. Phytopathology, 2007, 97(11):1458-1466.
[19] 金麗穎. 抗黃瓜棒孢葉斑病放線菌的篩選及Streptomyces
inhibens 多相分類研究[D].哈爾濱:東北農(nóng)業(yè)大學(xué),2019.
JIN L Y. Screening of actinomycetes against cucumber target leaf
spot disease and polyphasic taxonomy study of Streptomyces
inhibens [D]. Harbin: Northeast Agricultural University, 2019.
[20] XIONG Z Q, TU X R, WEI S J, et al .. The mechanism of
antifungal action of a new polyene macrolide antibiotic
antifungalmycin 702 from Streptomyces padanus JAU4234 on
the rice sheath blight pathogen Rhizoctonia solani [J/OL]. PLoS
One, 2013, 8(8):e73884 [2023-04-30]. https:doi.org/10.1371/
journal.pone.0073884.
[21] 董章勇,王振中.植物病原真菌細(xì)胞壁降解酶的研究進(jìn)展[J].
湖北農(nóng)業(yè)科學(xué), 2012,51(21): 4697-4700.
DONG Z Y, WANG Z Z. Research progress of fungal cell walldegrading
enzyme [J]. Hubei Agric. Sci., 2012, 51(21):4697-
4700.
[22] 臧超群,白元俊,張海東,等.暗黑鏈霉菌PY-1活性產(chǎn)物分析
及其對(duì)葡萄霜霉病田間防效評(píng)價(jià)[J].植物保護(hù)學(xué)報(bào), 2018,
45(4): 864-870.
ZANG C Q, BAI Y J, ZHANG H D, et al .. Study on bioactive
metabolite of Streptomyces atratus PY-1 and the field control
efficiency against grapevine downy mildew [J]. J. Plant Prot.,
2018, 45(4): 864-870.
[23] 榮良燕,姚拓,趙桂琴,等.產(chǎn)鐵載體PGPR菌篩選及其對(duì)病原
菌的拮抗作用[J].植物保護(hù), 2011, 37(1):59-64.
RONG L Y, YAO T, ZHAO G Q, et al .. Screening of
siderophore-producing PGPR bacteria and their antagonism
against the pathogens [J]. Plant Prot., 2011, 37(1):59-64.
[24] MA D C, ZHU J M, JIANG J G, et al .. Evaluation of bioactivity
and control efficacy of tetramycin against Corynespora
cassiicola [J]. Pest. Biochem. Physiol., 2018, 152(12):106-113.
