豫南小麥品種(系)重要功能基因的KASP檢測與應用
2024-11-07 00:00:00陳真真周國勤申冠宇蔡晨陽陳俊
江蘇農業科學 2024年18期


摘要:為了解豫南地區小麥品種(品系)重要性狀功能基因組成,利用KASP標記技術對33份小麥品種(品系)光周期、株高、粒重、穗粒數、生物量、谷蛋白亞基、籽粒蛋白相關基因、多酚氧化酶活性、黃色素含量、抗逆等相關功能基因進行檢測。結果表明:在光周期Ppd-D1基因上33份材料均表現出光周期不敏感型;在Vrn-A1、Vrn-5A基因上33份材料均表現出冬性品種,32份材料在Vrn-D1基因上表現出春性品種,16份材料檢測出矮稈基因Rht-D1b。產量性狀相關基因中TaGS5-A1、TaGW2-6B、TaTGW-7A、TaGS-D1、TaSus2-2B、TaTGW6-A1、Sus1-7A、TaCWI-A1和TaGS2-A1高千粒重優異等位變異分布比率分別為3.03%、36.36%、93.94%、18.18%、42.42%、3.03%、100%、54.55%和81.82%。高小穗數基因TaMoc-A1優異等位變異分布比率為3.03%;高單株生物量TaGS2-B1的優異等位變異分布比率為48.48%。籽粒品質相關基因中低分子量麥谷蛋白亞基Glu-A3d、Glu-B3g、Glu-B3e、Glu-A3b的分布比率分別為3.03%、75.76%、100%和96.97%;Glu-A1的弱筋等位變異null分布比率為48.48%;小麥籽粒蛋白相關基因的NAM-A1b,A1d分型和NAM-A1a,A1b分型分布比率分別為93.94%和78.79%。面粉品質相關基因中八氫番茄紅素脫氫酶基因Pds-B1、八氫番茄紅素合成酶基因PsyB1c和PsyA1b的優異等位變異低黃色素含量的的分布比率分別為42.42%、100%和100%;多酚氧化酶基因TaPpoA1優異等位變異TaPpo-A1b酶基因低含量的分布比率為30.3%;面團強度ALPb7A優異等位變異ALPb7Aa高面團強度的分布比率為45.45%。抗穗發芽基因中Vp-B1和TaSdr-B1的優異等位變異抗穗發芽所占的比例為100%和60.61%;抗旱Dreb-B1和1fehw3基因的優異等位變異分布比率分別為69.70%和90.90%;高莖稈木質素含量優異等位變異COMT3Ba分布比率分別為100%。本研究明確了豫南地區小麥品種(品系)的重要功能,為生產布局和品種參試提供了理論依據。
關鍵詞:豫南;小麥;功能基因;KASP檢測;應用
中圖分類號:S512.103;S330 文獻標志碼:A
文章編號:1002-1302(2024)18-0128-10
收稿日期:2024-03-19
基金項目:河南省“揭榜掛帥”技術攻關專項(編號:21110110800);國家小麥產業技術體系建設專項(編號:CARS-03-01A);河南省小麥產業技術體系項目。
作者簡介:陳真真(1990—),女,河南駐馬店人,碩士,助理研究員,主要從事小麥遺傳育種研究,E-mail:396939159@qq.com;周國勤,副研究員,主要從事小麥遺傳育種與栽培工作研究,E-mail:zhouguoqin74@126.com。
小麥(Triticum aestivum L.)是三大糧食作物之一,是種植面積最大和分布最廣的糧食作物。根據國家統計局官方發布,我國2022年小麥播種面積約2 351.85萬hm2,占全國糧食播種總面積(11 833.2萬hm2)的1/5左右,小麥播種面積約占全球小麥播種面積的11%。因此,小麥生產安全對國家糧食安全和世界糧食安全都有著重要的影響。豫南地區的土壤、氣候生態條件等非常有利于小麥的生產,為河南省糧食生產核心區。南方的品種在該區域產量潛力有限,銹病重;北方的小麥品種在該區域不耐漬害,赤霉病重,不抗穗發芽,晚熟。現在豫南地區生產上應用的品種主要是以江蘇、四川、信麥系列品種為主。豫南地區主推小麥品種的重要性狀功能基因研究對長江中下游小麥種質資源創新和生產上的推廣應用具有非常重要的作用。
近年來,隨著小麥全基因組測序的完成和高通量測序技術的發展,基于小麥基因組開發的SNP(single nucleotidepolymorphism )和InDels (insertion-deletion)等標記被廣泛用于關聯分析、基因定位和遺傳圖譜的構建等方面的研究,為小麥多基因聚合育種和分子標記輔助選擇奠定了基礎[1-2]。競爭性等位基因特異性PCR(kompetitiveallele-specificPCR,KASP) 技術是一種高通量的SNP分型技術,其是由英國LGC公司開發的,其具有對樣品的基因組DNA進行SNP分型和鑒定多個性狀的功能[3]。KASP技術與SSR、KFLP等傳統分子標記相比更靈活、更經濟、更準確,而且其更加適合大樣本檢測,其在篩選植物優良抗逆基因、優良的適應性基因、品質基因以及篩選優良的種質材料等方面具有重要的價值[4-8]。
近年來,基于KASP標記技術對各種作物種質資源基因分型或作物輔助育種的研究越來越多。閆金龍等對晉東南小麥品種(系)部分抗病基因的KASP標記檢測,明確了晉東南地區小麥品種(系)中抗病基因分布[9];權有娟等利用KASP標記檢測青海和西藏小麥品種中光周期基因分布,明確了青海和西藏小麥品種中光周期基因的分布情況[10];王志偉等對云南小麥品種(系)抗逆性相關基因的KASP標記進行檢測,明確了云南小麥品種中抗旱和抗穗發芽等抗逆性狀相關基因的組成分布;高振賢等對河北省小麥重要農藝性狀的KASP標記檢測,明確了河北小麥重要農藝性狀的優異等位變異[12];王君嬋等對揚麥系列品種(系)重要性狀功能基因的KASP進行檢測,明確了揚麥系列品種(系)部分重要性狀功能基因的組成[5];杜瑩瑩等對江蘇淮北小麥品種(系)重要性狀功能基因的KASP檢測,明確了淮北麥區育成品種(系)的優異等位變異組合[13]。
本研究對豫南地區大面積推廣的小麥品種以及信陽市農業科學院育成的品種和高代穩定品系進行了KASP標記選擇,以期明確光周期、株高、粒重、穗粒數、生物量、谷蛋白亞基、籽粒蛋白相關基因、多酚氧化酶活性、黃色素含量、抗逆等相關功能基因等性狀相關基因的組成,為準確評價推廣品種和育成材料的利用價值提供參考,為生產布局和品種參試提供依據。
1 材料與方法
1.1 試驗材料
試驗材料為33份,其中8份是信陽市農業科學院育成的品種和高代穩定品系,有25份是在豫南地區種植面積較大的品種于2023—2024年度種植于信陽市農業科學院陸廟基地。
1.2 KASP檢測
1.2.1 DNA提取 首先用DNA 提取試劑盒提取DNA,整個過程在Douglas Scientific Array Tape平臺上完成,先用紫外分光光度計檢測DNA的含量及純度,然后根據小麥基因組大小,利用Tecan液體自動化工作站進行DNA的稀釋。
1.2.2 PCR體系配制 在Nexar工作站上將DNA工作液、PCR mix 及引物加入到384 PCR反應Array Tape中,PCR總反應體系1.622 μL,DNA 0.8 μL,2×KSPMaster Mix 0.8 μL,KASP Assay mix 0.022 μL。
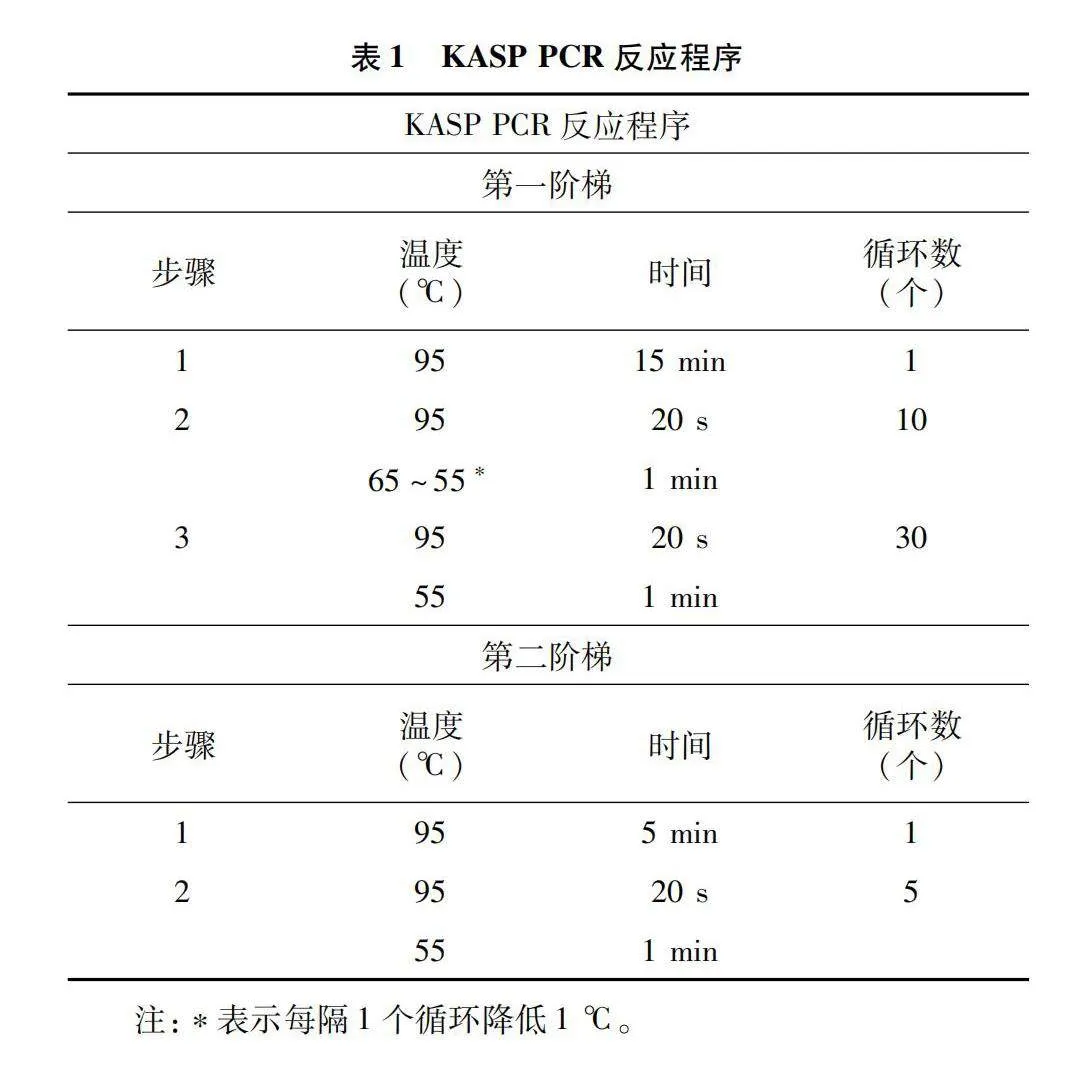
1.2.3 PCR擴增 在Soellex水浴中完成PCR反應,PCR反應程序見表1。
1.2.4 熒光信號讀取 在Araya上進行熒光強度的掃描檢測,在相應軟件中分析和讀取數據。
2 結果與分析
2.1 適應性相關基因的KASP檢測結果
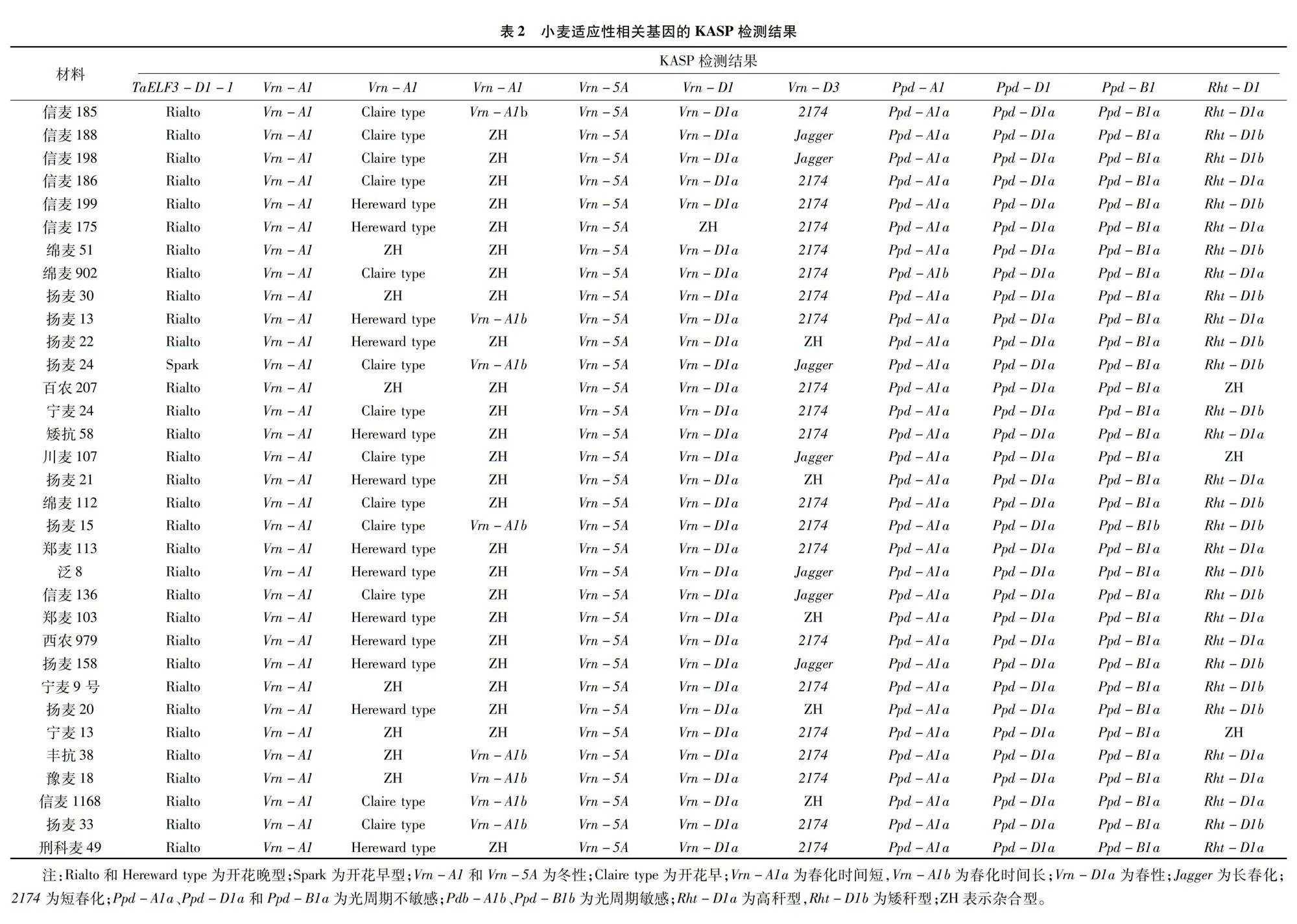
為了了解豫南地區小麥品種的廣泛適應性特點,檢測了TaELF3-D1-1和Vrn-A1等花期相關基因、Ppd-A1、Ppd-D1和Ppd-B1等光周期相關基因、Vrn-A1和Vrn-D3等春化相關基因、Vrn-A1、Vrn-5A和Vrn-D1等冬春性相關基因、Rht-D株高相關基因的的基因型,KASP標記檢測結果(表2)顯示:32份小麥品種的TaELF3-D1-1基因均表現出開花晚型,13份小麥品種的Vrn-A1表現出開花早;光周期Ppd-A1基因有32份材料表現出光周期不敏感,只有綿麥902表現出光周期敏感;在光周期Ppd-D1基因上33份材料均表現出光周期不敏感;在光周期Ppd-B1基因上32份材料均表現出光周期不敏感;Vrn-A1和Vrn-5A基因33份材料均表現出冬性品種,32份材料在Vrn-D1上表現出春性品種,16份材料檢測出矮稈基因Rht-D1b。
2.2 產量相關基因的KASP檢測結果
12個與粒重有關的基因TaGS5-A1、TaCWI-5D、TaGW2-6B、TaTGW-7A、TaGS-D1、TaSus2-2B、TaTGW6-A1、Sus1-7A、Sus2-2A、TaCWI-A1、TaSus、TaGS2-A1,1個與小穗數TaMoc有關的基因,1個與單株生物量TaGS2-B1有關的基因KASP標記檢測結果(表3)表明:1份小麥品種(系)含有TaGS5-A1的高千粒重優異等位變異TaGS5-A1a;33份小麥品種(系)均未含有TaCWI-5D的粒重有利等位基因單倍型;含有TaGW2-6B的高粒重單倍型Hap-3優異等位變異的小麥品種(系)有12份,優異等位基因所占的比例為36.36%;含有TaTGW-7A的高千粒重優異等位變異TaTGW-7Aa小麥品種(系)31份,優異等位基因所占的比例為93.94%;含有TaGS-D1高千粒重優異等位變異TaGS-D1a小麥品種(系)6份,優異等位基因所占的比例為18.18%;含有TaSus2-2B高粒重單倍型Hap-H小麥品種(系)14份,優異等位基因所占的比例為 42.42%;含有TaTGW6-A1的高粒重優異等位變異TaTGW6-A1a小麥品種(系)1份,優異等位基因所占的比例為3.03%;含有Sus1-7A的高粒重單倍型Hap-4/Hap-2小麥品種(系)33份;優異等位基因所占的比例為100%;33份小麥品種(系)均未含有高千粒重單倍型;含有TaCWI-A1的高千粒重優異等位變異TaCwi-A1a的小麥品種(系)18份,優異等位基因所占的比例為54.55%;33份小麥品種(系)均未含有TaSus1-7B的高千粒重單倍型;含有TaGS2-A1的高千粒重優異等位變異TaGS2-A1a小麥品種(系)27份,優異等位基因所占的比例為81.82%;含有TaMoc-A1高小穗數優異單倍型Hap-H小麥品種(系)1份,優異等位基因所占的比例為3.03%;含有TaGS2-B1的高單株生物量的優異等位變異TaGS2B1a小麥品種(系)16份,優異等位基因所占的比例為48.48%;試驗結果表明這33份小麥品種(系) 基本上都含有3個以上的優異基因,進一步分析發現信麥185、信麥188、信麥198、信麥186和豫麥18這5份小麥品種(系)中都含有6個與產量相關的優異基因。
2.3 籽粒品質相關基因的KASP檢測結果
供試材料中Glu-A3d低分子量麥谷蛋白亞基位點優質亞基的分布比例為3.03%(表4);Glu-A3g低分子量麥谷蛋白亞基在33份材料中都沒有檢測到,含有Glu-A1的弱筋等位變異null小麥品種(系)16份,優異等位基因所占的比例為48.48%;含有Glu-B3g低分子量麥谷蛋白亞基小麥品種(系)25份,分布比例為75.76%;33份小麥品種(系)含有Glu-B3e低分子量麥谷蛋白亞基,分布比例為100%;Glu-B3fg低分子量麥谷蛋白亞基在33份材料中都沒有檢測到;GCP--高蛋白質含量在33份材料中都沒有檢測到;32份小麥品種(系)含有Glu-A3b低分子量麥谷蛋白亞基低分子量麥谷蛋白亞基,分布比例為96.97%;31份小麥品種(系)含有NAM-A1的小麥籽粒蛋白相關基因的NAM-A1b,A1d分型,分布比例為93.94%;26份小麥品種(系)含有NAM-A1的小麥籽粒蛋白相關基因的NAM-A1a,A1b分型,分布比例為78.79%;R_B1b為紅粒基因在33份小麥品種(系)都有檢測到,分布比例為100%;進一步分析發現:信麥188、信麥198、綿麥51、綿麥902、揚麥13、揚麥22、綿麥112、泛8、信麥136、揚麥158、寧麥9號、揚麥20、信麥1168均含有Glu-A1的弱筋等位變異null,Glu-B3g低分子量麥谷蛋白亞基,Glu-B3e低分子量麥谷蛋白亞基,Glu-A3b低分子量麥谷蛋白亞基。
2.4 面粉品質相關基因的KASP檢測結果
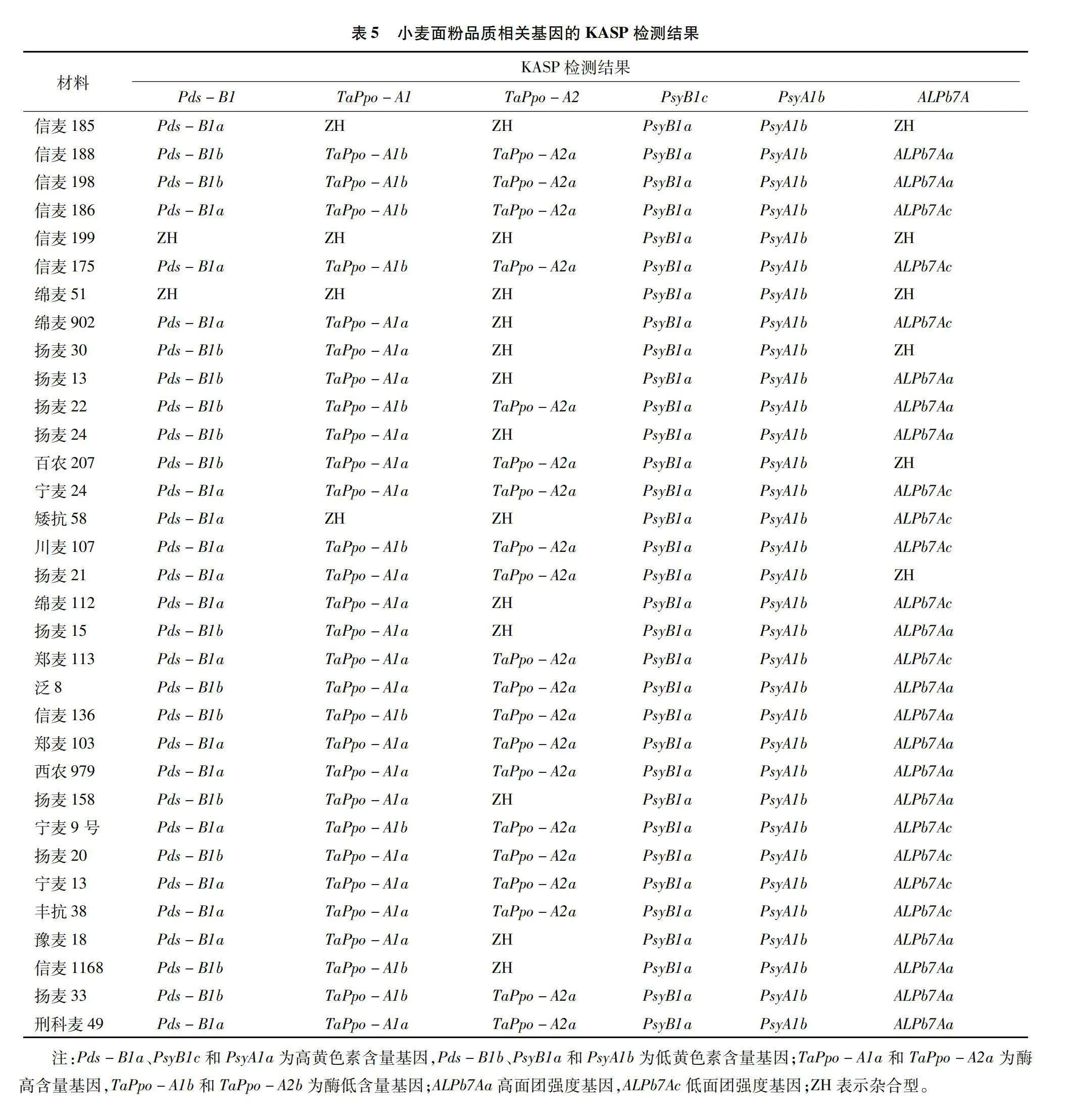
從表5可以看出,八氫番茄紅素脫氫酶Pds-B1優異等位變異Pds-B1b低黃色素含量的分布比例為42.42%;八氫番茄紅素合成酶PsyB1c優異等位變異PsyB1a低黃色素含量的分布比例為100%;八氫番茄紅素合成酶PsyA1b優異等位變異PsyA1b低黃色素含量的分布比例為100%;多酚氧化酶TaPpoA1優異等位變異TaPpo-A1b酶低含量的分布比例為30.3%;多酚氧化酶TaPpo-A2優異等位變異TaPpo-A2b酶低含量的分布比例為0;面團強度ALPb7A的優異等位變異ALPb7Aa高面團強度的分布比例為45.45%;面粉品質優異等位變異組合Pds-B1b+PsyB1a+PsyA1b+TaPpo-A1b+ALPb7Aa,品種(品系)有信麥188、信麥198、信麥1168、揚麥33等,占比12.12%。
2.5 抗逆性相關基因的KASP檢測結果
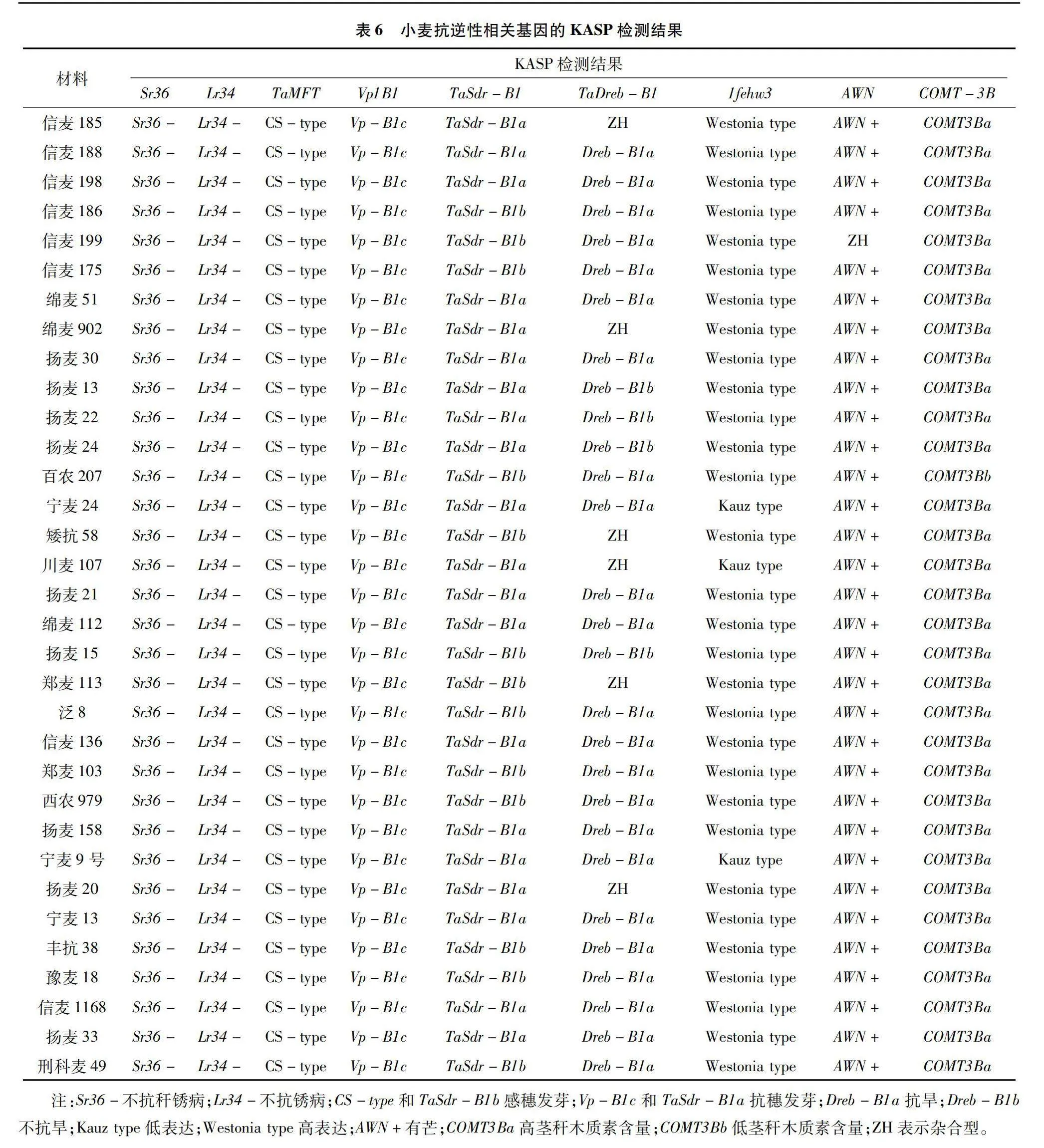
從表6可以看出,在供試材料中沒有檢測到抗稈銹病基因Sr36的優異等位基因變異Sr36+;沒有檢測到抗葉銹病Lr34的優異等位基因變異Lr34+;抗穗發芽的基因中,TaMFT-A1的優異等位變異分布比例為0;Vp-B1的優異等位變異抗穗發芽Vp-B1c的分布比例為100%;含有TaSdr-B1的優異等位變異抗穗發芽TaSdr-B1a小麥品種(品系)20份,優異等位基因所占的比例為60.61%;所以有信麥188、信麥198、信麥1168、綿麥51、綿麥902、揚麥30、揚麥22、揚麥13、信麥136等20份小麥品種(品系)同時含有抗穗發芽的優異等位變異Vp-B1c和TaSdr-B1a;23份小麥品種(品系)含有抗旱的優異等位變異Dreb-B1a,分布比例為69.70%;30份小麥品種(品系)含有抗旱1fehw3優異等位變異Westonia type,分布比例為90.9%;信麥188、信麥198、信麥186、信麥199、信麥175、綿麥112、鄭麥103、西農979、豫麥18、信麥1168等21份小麥同時含有抗旱的優異等位變異組合Dreb-B1a+Westonia type;占比63.64%;32份小麥品種(品系)均表現出有芒,1份小麥品種(品系)表現出雜合型;33份材料均含有高莖稈木質素含量優異等位變異COMT3Ba,分布比例100%。
3 結論與討論
3.1 適應性相關基因的分布與應用
對33份試驗材料的適應性相關基因的KASP檢測結果表明,Vrn-A1、Vrn-D1 和 Vrn-D3等春ab8cbe918c9a0e2636e0aa82fb6effbd09cde67f1a66871b232c7c419f113f35化基因在33份小麥品種(品系)中有著較高的利用率,Vrn-A1和Vrn-5A等春性早花基因在這些材料中有著較低的利用率;信麥185、信麥136、信麥1168等檢測出同時含有 Vrn-A1早開花基因和Ppd-D1a光周期不敏感基因,因其生態適應性較好可以在豫南地區種植。豫南地區屬于長江中下游冬麥區,作物種植制度一般為“水稻—小麥”一年兩熟制,一方面豫南地區由于優質粳稻、直播稻、機插秧稻種植面積擴大,水稻騰茬收獲較晚導致小麥播種較晚,另一方面水稻收獲小麥播種期間由于遭遇連陰雨天氣導致小麥播期推遲,這2個因素導致該地區小麥增產緩慢,在生產上產量高、成熟較早、后期灌漿速度快、播期彈性大、抗逆性好的小麥品種在該區域種植面積較大。揚麥系列品種、寧麥系列品種、綿麥系列品種和信麥系列品種含有較多的早開花基因和春化基因,種植或遺傳改良這些早熟高產品種,比如信麥1168、信麥136、揚麥15、揚麥13等品種,能更好地適應豫南地區小麥播種晚成熟早的需求。
3.2 產量相關基因的分布與應用
小麥產量的三要素是穗數、穗粒數、千粒重。千粒重對小麥產量的高低起著至關重要的作用,小麥育種中高產目標需要通過提高粒重來實現。千粒重在小麥產量三要素中是遺傳比較穩定的數量性狀,遺傳因素、環境條件和遺傳的加性都對其有影響[14]。粒重相關的基因主要有TaCWI-4A、TaCWI-5D、TaSus1、TaSus2、TaCwi-A1、TaGS3-D1、TaGS5-A1、TaGS3-D1、TaGW2、TGW6-A1等[15-23]。穗粒數相關的基因有TaMoc1-7A等[24]。這些基因的優異等位變異在小麥產量增加上具有重要作用。本研究利用KASP標記對12個與粒重有關的基因TaGS5-A1、TaCWI-5D、TaGW2-6B、TaTGW-7A、TaGS-D1、TaSus2-2B、TaTGW6-A1、Sus1-7A、Sus2-2A、TaCWI-A1、TaSus1-7B、TaGS2-A1,1個與小穗數TaMoc-A1有關的基因,1個與單株生物量TaGS2-B1相關的基因進行檢測,試驗結果表明33份小麥品種品種(系) 都聚合了3個以上(21.4%)的優異等位基因,其中7份小麥品種(系)中聚合6個(42.9%)產量相關優異位點。這可能是育種家在育種的時候對產量三要素進行了有目的的選擇,高千粒重和高小穗數的優異等位變異在品種選育過程中都被保留了下來。在以后的小麥品種選育工作中應該注意產量三要素的協同發展。
3.3 籽粒品質相關基因的分布與應用
小麥籽粒品質受多基因控制,與高分子量谷蛋白亞基、籽粒蛋白相關基因、低分子量谷蛋白亞基和蛋白質含量等有關系。高低分子量谷蛋白亞基對小麥面團彈性和延展性具有一定的影響,而其又受多基因控制,這些優異等位變異基因可以提高面粉加工品質。試驗結果表明,33份材料中Glu-A3d低分子量麥谷蛋白亞基位點優質亞基所占的比例為3.03%;Glu-A1的弱筋等位變異null分布比率為48.48%;Glu-B3g低分子量麥谷蛋白亞基分布比率為75.76%;Glu-B3e低分子量麥谷蛋白亞基分布比率為100%;由此表明,豫南麥區主要以弱筋低分子量小麥為主,豫南麥區的育種目標也是中筋偏弱,育種目標和生產實踐具有一定的切合度。含有Glu-A1的弱筋等位變異null的小麥品種比如信麥136、信麥1168、揚麥13和揚麥33等可作為該麥區中筋偏弱小麥育種的重要育種材料。
3.4 面粉品質相關基因的分布與應用
面粉品質與多酚氧化酶(PPO)活性、黃色素含量、面團強度、面筋質量等高度相關。研究表明多酚氧化酶活性與面制品制作過程中褐變密切相關,多酚氧化酶活性低,面制品質量好,與其相關的基因主要有TaPpo-1b、TaPpo-D1a、TaPpo-A2b、TaPpo-D2b[25]。小麥中黃色素主要由類胡蘿卜素組成,黃色素含量影響面粉的顏色,八氫番茄紅素脫氫酶Pds-B1,八氫番茄紅素合成酶PsyB1c、PsyA1b,胡蘿卜素脫氫酶等基因(Zds)控制類胡蘿卜素合成[26-27]。面團強度參數與面包烘焙體積呈正相關,也是量化面粉品質的重要指標[28]。本研究發現信麥188、信麥198、信麥1168、揚麥33等4個品種(品系)含有小麥面粉品質優異等位變異組合Pds-B1b+PsyB1a+PsyA1b+TaPpo-A1b+ALPb7Aa。
3.5 抗逆性相關基因的分布與應用
小麥抗逆相關基因主要包括抗病、抗蟲、抗旱、抗穗發芽、抗倒伏等。抗稈銹病基因主要有Sr2和Sr36、抗條銹病基因主要有Yr15和Yr36、抗旱基因主要有Dreb-B1和1fehw3等、小麥抗穗發芽相關基因主要有TaVP1-B1、TaSdr-B1、TaPHS1、TaMFT-A1等[29]。其中TaVp1的TaVp-1Bb和TaVp-1Bc基因型、TaSdr-B1a基因型也具有較高的穗發芽抗性[30-32]。本研究發現20份小麥品種(品系)同時含有抗穗發芽的優異等位變異Vp-B1c和TaSdr-B1a;同時含有抗旱的優異等位變異組合Dreb-B1a+Westonia type的小麥材料占比63.64%;33份材料均含有高莖稈木質素含量優異等位變異COMT3Ba;信麥188、信麥198、揚麥13、信麥1168、綿麥112等可以作為抗穗發芽、抗旱以及高莖稈木質素含量等的優質材料,在生產上應用。
參考文獻:
[1]CavanaghC R,Chao S M,Wang S C,et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars[J]. Proc Natl Acad Sci USA,2013,110(20):8057-8062
[2]Wang S.C,Wong D,Forrest K,et al,Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array[J]. Plant Biotechnology Journal,2014,12(6):787-796.
[3]Semagn K,Babu R,Hearne S,et al. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP):overview of the technology and its application in crop improvement[J]. Molecular Breeding,2014,33(1):1-14.
[4]趙永濤,張 鋒,張中州,等. 豫麥158及其硬質變異系重要性狀基因的KASP標記檢測[J]. 河南農業科學,2021,50(10):37-43.
[5]王君嬋,吳旭江,胡文靜,等. 揚麥系列品種(系)重要性狀功能基因的KASP檢測[J]. 江蘇農業學報,2019,35(6):1271-1283.
[6]楊子博,顧正中,周羊梅,等. 江蘇淮北地區小麥品種資源籽粒硬度基因等位變異的KASP檢測[J]. 麥類作物學報,2017,37(2):153-161.
[7]王 偉,王 斌,于 亮,等. 小麥疊氮化鈉誘變群體重要功能基因的KASP標記檢測[J]. 種子,2020,39(1):42-48.
[8]TrickM,Adamski N M,Mugford S G,et al. Combining SNP discovery from next-generation sequencing data with bulked segregant analysis(BSA)to fine-map genes in polyploid wheat[J]. BMC Plant Biology,2012,12(14):1-17.
[9]閆金龍,張東旭,馮麗云,等. 晉東南小麥品種(系)部分抗病基因的KASP標記檢測[J]. 作物雜志,2023(5):49-58.
[10]權有娟,袁飛敏,劉德梅,等. 利用KASP標記檢測青海和西藏小麥品種中光周期基因分布[J]. 麥類作物學報,2019,39 (10):1165-1172.
[11]王志偉,王志龍,喬祥梅,等. 云南小麥品種(系)銹病和赤霉病抗性功能基因的KASP標記檢測[J]. 作物雜志,2020(1):187-193.
[12]高振賢,趙彥坤,班進福,等. 河北省小麥重要農藝性狀的KASP標記檢測[J]. 分子植物育種,2021,19(2):518-528.
[13]杜瑩瑩,顧正中,周羊梅,等. 江蘇淮北小麥品種(系)重要性狀功能基因的KASP檢測[J]. 麥類作物學報,2023,43(3):279-287.
[14]張福彥,范家霖,陳曉杰,等. 小麥粒重相關基因的遺傳定位和分子標記輔助育種進展[J]. 植物遺傳資源學報,2020,21 (3):507-516.
[15]Jiang Y M,Jiang Q Y,Hao C Y,et al. A yield-associated gene TaCWI,in wheat:its function,selection and evolution in global breeding revealed by haplotype analysis[J]. Theoretical and Applied Genetics,2015,128(1):131-143.
[16]Jiang Q Y,Hou J,Hao C Y,et al. The wheat(T.aestivum)sucroses synthease 2 gene(TaSus2)active in endosperm development is associated with yield traits[J]. Functional Intetive Genomics,2011,11(1):49-61.
[17]Ma D Y,Yan J,He Z H,et42OfYdWTf9qYF9+MKyt4Gg== al. Characterization of a cell wallin vertasegene TaCwi-A1 on common wheat chromosome 2A and development of functional markers[J]. Molecular Breeding,2012,29(1):43-52.
[18]Zhang Y J,Liu J D,Xia X C,et al. TaGS-D1,an ortholog of rice OsGS3,isassociated with grain weight and grain length incommon wheat[J]. Molecular Breeding,2014,34(3):1097-1107.
[19]Wang S S,Zhang X F,Chen F,et al. A single-nucleotide polymorphism of TaGS5 gene revealed its association with kernel weight in Chinese bread wheat[J]. Frontiers in Plant Science,2015,6:1166-1166.
[20]Yang Z B,Bai Z Y,Li X L,et al. SNP identification and allelic-specific PCR markers development for TaGW2,a gene linked to wheat kernel weight[J]. Theoretical and Applied Genetics,2012,125(5):1057-1068.
[21]Geng J,Li L Q,Lv Q,et al. TaGW2-6A allelic variation con-tributes to grain size possibly by regulating the expression of cytokinins and starch-related genes in wheat[J]. Planta,2017,246(6):1153-1163.
[22]Qin L,Hao C Y,Hou J,et al. Homologous haplotypes,expression,genetic effects and geographic distribution of the wheat yield gene TaGW2[J]. BMC Plant Biology,2014,14(1):107.
[23]Hanif M,Gao F M,Liu J D,et al. TaTGW6-A1,an ortholog of rice TGW6,is associated with grain weight and yield in bread wheat[J]. Molecular Breeding,2015,36(1):2016.
[24]Zhang B,Liu X,Xu W N,et al. Novel function of a putative MOC1 ortholog associated with spikelet number per spike in common wheat[J]. Scientific Reports,2015,5(1):12211-12211.
[25]He X Y,He Z H,Zhang L P,et al. Allelic variation of polyphenol oxidase(PPO) genes located on chromosomes 2A and 2D and development of functional markers for the PPO genes in common wheat [J]. Theoretical and Applied Genetics,2007,115(1):47-58.
[26]He X Y,He Z H,Ma W,et al. Allelic variants of phytoene synthase 1(Psyl)genes in Chinese and CIMMYT wheat cultivars and development of functional markers for flour colour [J]. Molecular Breeding,2009,23(4):553-563.
[27]Dong H C,Ma Z Y,Xia X C,et al. Allelic variation at theTaZds-A1locus on wheat chromosome 2A and develop-ment of a functional marker in common wheat [J]. Journal of Integrative Agriculture,2012,11(7):1067-1074.
[28]Delcour J A,Joye I J,Pareyt B,et al. Wheat gluten functionality as a quality determinant in cereal-based food products [J]. Annual Review of Food Science and Technology,2012,3(1):469-492.
[29]Aravindh R,Sivasamy M,Ganesamurthy K,et al. Marker assisted stacking/pyramiding of stem rust,leaf rust and powdery mildew disease resistance genes(Sr2/Lr27 /Yr30,Sr24/Lr24 and Sr36 /Pm6) for durable resistance in wheat(Triticum aestivum L.) [J]. Electronic Journal of Plant Breeding,2020,11(3):907-915.
[30]黃義文,代旭冉,劉宏偉,等. 小麥抗穗發芽基因挖掘及分子育種進展[J]. 麥類作物學報,2021,41(2):147-156.
[31]Yang Y,Zhang C L,Liu S X,et al. Characterization of the rich haplotypes of Viviparous-1A in Chinese wheats and development of a novel sequence tagged site marker for preharvest sprouting resistance [J]. Molecular Breeding,2014,33 (1):75-88.
[32]Zhang Y J,Miao X L,Xia X C,et al. Cloning of seed dormancy genes(TaSdr) associated with tolerance to preharvest sprouting in common wheat and development of a functional marker [J]. Theoretical and Applied Genetics,2014,127(4):855-866.
