基于轉錄組數據的濕地松密碼子使用偏好性分析
2024-11-07 00:00:00胡蓉賴猛張露陳婷萱黃文丁易敏
江蘇農業科學 2024年18期
摘要:研究濕地松轉錄組編碼序列的密碼子使用偏好性,明確密碼子使用頻率,探究濕地松異源表達受體的最佳選擇,為濕地松分子育種奠定基礎。以篩選到的13 932條濕地松轉錄組編碼序列(coding sequence,CDS)為研究對象,利用CodonW 1.4.2、EMBOSS在線分析程序、MEGA 7等軟件進行濕地松密碼子組成分析,計算同義密碼子相對使用度(relative synonymous codon usage,RSCU)、不同位置GC含量和有效密碼子數(effective number of codon,ENc)等指標,同時展開中性繪圖分析和偏倚性分析等,系統地探究影響密碼子偏好性的主導因素,從而篩選出最優密碼子,并選擇合適的遺傳轉化受體。研究結果表明,濕地松轉錄組密碼子中的平均GC含量為43.72%,其中GC1含量最大,為50.78%,其次是GC2和GC3,含量分別為40.68%和39.71%,表明不同位置的堿基組成存在差異,第3位上是A/U堿基的概率更大;ENc值在25.94~61.00之間,平均值為53.58,CAI均值較低,為0.20,表明密碼子整體偏好性較弱。中性繪圖、PR2-plot繪圖和ENc-GC3s關聯分析結果均表明,自然選擇是大多數濕地松基因密碼子存在使用偏好性的主要原因;以高頻率高表達的標準篩選出25個濕地松最優密碼子,均以A/U結尾。通過將濕地松與大腸桿菌、擬南芥、煙草、酵母菌、水稻等5種生物的密碼子使用頻率進行對比分析得出,煙草、擬南芥是濕地松基因進行遺傳轉化功能驗證的理想選擇。綜上所述,濕地松轉錄組密碼子整體偏好性弱,第3位堿基偏好使用A/U,自然選擇是L/glweODKoYek9h8Qj5+dlZ7Sw06tEPlLdpOzyfAu84=其產生密碼子偏好性的關鍵原因,煙草、擬南芥可作為濕地松基因異源表達的理想植物受體。結果有利于濕地松功能基因的異源表達,也為進一步研究濕地松遺傳背景及高產脂分子機制提供科學依據。
關鍵詞:濕地松;轉錄組;最優密碼子;密碼子使用偏好性
中圖分類號:S718.46 文獻標志碼:A
文章編號:1002-1302(2024)18-0050-09
收稿日期:2023-10-12
基金項目:江西省重點研發計劃一般項目(編號:20203BBFL63058);江西省自然科學基金青年基金(編號:20212BAB215013);國家自然科學基金(編號:32160385);江西省林業廳林業科技創新專項(編號:202311)。
作者簡介:胡 蓉(1997—),女,云南大理人,碩士研究生,主要從事林木遺傳改良研究。E-mail:hurong089@163.com。
通信作者:易 敏,博士,副教授,主要從事濕地松遺傳改良研究。E-mail:yimin6104@163.com。
密碼子(codon)又稱遺傳密碼,由mRNA上相鄰的3個核苷酸構成,在中心法則翻譯過程中作為遺傳信息和蛋白質之間的重要紐帶,起決定性作用[1]。生物體內共有64種密碼子,除色氨酸(Trp)和甲硫氨酸(Met)外,存在同一氨基酸由2個及以上密碼子編碼的簡并現象,對應的編碼密碼子被稱為同義密碼子[2-3]。同義密碼子在沒有選擇壓力和突變基因偏倚作用的情況下使用頻率是一致的[4]。但生物體為適應環境變化,會偏好優選適合自身生長發育的最優密碼子,同義密碼子間選擇使用頻率存在差別,有密碼子偏好性(codon usage bias,CUB)[5]。密碼子偏好性的產生受到GC含量、tRNA豐度、選擇壓力、突變壓力、基因表達水平等多種因素的影響[6-7]。因此,針對植物的密碼子偏好性進行分析,探尋密碼子使用偏好模式在不同物種間的異同,有助于進一步了解物種相關基因的功能表達機制、親緣關系和進化歷程,也可為基因工程研究中最優密碼子、最適外源宿主選擇提供一定的理論依據。
目前,針對植物密碼子偏好性的研究報道不少,但在針葉樹種中的研究仍以利用葉綠體基因組進行分析較為常見,如思茅松[Pinus kesiya var. langbianensis (A.Chev ) Gaussen][8]、高山松(P. densata Mast.)[9]、云南油杉(Keteleeria evelyniana Mast.)[10]等。高通量測序技術的發展與大量轉錄組序列的公布為基于轉錄組的密碼子相關研究提供了數據基礎。目前,已在樟樹[Cinnamomum camphora (L.) Presl]、橄欖[Canarium album (Lour.) Rauesch.][11]、杜仲(Eucommia ulmoides Oliver)等多種植物上均有報道,但在松屬(Pinus)中僅在馬尾松[12]上有相關報道,在濕地松上尚未見有相關方面的研究。
濕地松(Pinus elliottii Engelm)原產于美國東南部,是松科(Pinaceae)松屬(Pinus)速生常綠針葉喬木,20世紀30年代引入我國,由于適應性強、生長快、松脂質量好以及松香不易結晶、不易凝固、雜質少等優點,已成為我國人工栽培面積最大的外來產脂樹種[13-14]。松脂是由松樹樹脂道的泌脂細胞產生的一種天然樹脂,由揮發性的松節油(單萜和倍半萜)和非揮發性的松香(雙萜)等萜類化合物組成,被廣泛應用于紙漿材、食品、農藥、生物燃料和醫藥等多個領域,是林業中最重要的產品之一[15]。針對濕地松的高產脂遺傳改良,目前已先后進行了種源試驗、采脂工藝、產脂相關性狀遺傳變異、松脂成分等方面的研究[16-20],并取得了一定的進展。松脂產量及品質等為重要經濟性狀,同時又屬于多基因控制的數量性狀,易受環境影響,導致僅利用常規育種手段往往難以滿足定向培育高產脂濕地松新品種的要求,因此,亟需運用現代林木生物技術來彌補常規育種技術的不足,充分挖掘和利用濕地松基因資源中的有利變異,加速高產、優質濕地松新品種的選育進程。盡管濕地松的分子研究起步較遲,但在濕地松TPS基因的克隆[21]、松脂合成基因的挖掘[13,22]、松脂性狀的關聯分析[23]等方面已取得一定的研究進展。但目前濕地松轉基因技術仍存在瓶頸,基因功能的驗證只能借助其他異源物種,導致其表達效率高低無法預測。因此,了解濕地松和轉化物種在同義密碼子使用頻率上的差異有利于提高濕地松功能基因在異源物種中的表達效率,從而為濕地松基因功能驗證提供重要的理論依據。
本研究以不同產脂量濕地松單株通過轉錄組測序獲得的13 932條序列為研究對象,利用軟件CodonW和在線網站EMBOSS Explore進行密碼子偏好性分析[11],獲得最優密碼子,明確密碼子使用頻率,探究濕地松異源表達受體的最佳選擇,對進一步揭示濕地松的遺傳背景及其高產脂機制具有重要作用,旨在為后續濕地松分子育種研究奠定基礎。
1 材料與方法
1.1 序列獲取與篩選
2016年4、7、10月分別從江西省吉安市白云山林場(27.22°N,115.13°E)1990年造林的濕地松家系測定林中選擇高、中、低產脂量的單株木質部材料,用液氮速凍后,交由北京百邁客生物信息科技有限公司利用Illumina Hiseq 2000平臺開展RNA-seq測序[13]。對原始測序數據進行過濾后,采用Trinity進行轉錄本序列拼接,并將測序結果提交至NCBI數據庫(http://www.ncbi.nlm.nih.gov/),登錄號為PRJNA723669。對所獲取的CDS序列使用perl程序[24]進行篩選,每條CDS序列應滿足堿基數為3的倍數,且長度≥300 bp,最終篩選得到含有13 932條序列的fasta文件。
1.2 密碼子使用偏好性參數指標
利用CodonW 1.4.2軟件 (http://codonw.sourceforge.net)對濕地松進行轉錄組密碼子使用偏好性參數分析。密碼子的偏好程度可由RSCU值體現,當值為0時,密碼子不存在偏好性,當RSCU>1時,表示偏好性強,為高頻密碼子,反之則使用頻率較低,為低頻密碼子。ENc是用于衡量基因中偏好程度的參8735f70668aeb4dc1b2a5d930344006a數,能將偏好程度作數據化呈現。ENc的取值范圍在20~61之間,低于35則認為密碼子使用具有高偏好性。密碼子適應性指數(codon adaptative index,CAI)介于0~1之間,越接近1,表明偏好性越強,反之則偏好性弱。同時利用軟件計算密碼子偏好指數(codon bias index,CBI)和最優密碼子使用頻率(frequency of optimal codons,Fop)等參數。使用EMBOSS Explore在線軟件中的CUSP程序(https://www.bioinformatics.nl/emboss-explorer/)分別計算濕地松轉錄組序列中密碼子第1、2、3位GC含量,同義密碼子第3位的GC含量及平均GC含量(GC1、GC2、GC3、GC3s、GCall)。
1.3 中性繪圖分析
以GC3值為橫坐標,GC12[GC12=(GC1+GC2)/2]為縱坐標繪制散點圖。若回歸系數趨于1,表示以突變為主導因素,反之趨于0,則以自然選擇為主導因素。
1.4 ENc-GC3s關聯分析
以GC3s值、ENc值分別為橫坐標、縱坐標繪制散點圖。設置ENc=2+GC3s+29/[GC3s2+(1-GC3s)2]的ENc理論值標準曲線[25-26]。標準曲線可反映僅受突變壓力影響時,GC3s值及ENc值變化情況。密碼子偏好性主要受堿基突變影響時,每個序列的實際ENc值都將落在標準理論曲線附8735f70668aeb4dc1b2a5d930344006a近或曲線上,反之ENc值遠離標準曲線時,則表示自然選擇或其他因素在密碼子偏好性中起主導作用[27]。同時根據吉德娟等在霸王(Sarcozygium xanthoxylon Bunge)中使用的ENc頻率分布統計方法[ENc比值=(ENc理論值-ENc實際值)/ENc理論值][28]判斷在密碼子偏好性中起主導作用的因素,若ENc比值>0.05或ENc比值<-0.05,自然選擇在密碼子偏好性中起到主要作用,反之,則是堿基突變為主要影響因素。
1.5 偏倚分析(PR2-plot分析)
以G3s/(G3s+C3s)和A3s/(A3s+U3s)分別作橫坐標、縱坐標繪制散點圖,其中A3s、U3s、C3s、G3s分別表示同義密碼子第3位堿基為A、U、C、G堿基的含量。每條序列代表1個散點,序列點與中心點的矢量距離表示偏好性程度和方向。根據偏倚規則,2條互補鏈間不存在偏倚情況即在無突變和自然選擇影響時[29],A=U、C=G,處于散點圖的中心位置。
1.6 最優密碼子的分析
利用高頻率高表達的方法篩選出最優密碼子,即ENc值較高的為低表達基因,反之為高表達基因。將ENc值從小到大排序,頭尾各篩選序列數的10%,建立高低表達基因組,計算其相對使用度的差值ΔRSCU,將ΔRSCU≥0.08定義為高表達密碼子;以RSCU值判定高低頻密碼子,即值大于1,被定義為高頻密碼子。若同時兼具RSCU值>1且其高低表達基因組RSCU差值(ΔRSCU)≥0.08,則為最優密碼子。
1.7 密碼子頻率分析
密碼子使用頻率比值能在一定程度上反映物種間密碼子偏好的差異大小,在基因的異源表達分析中起到重要的參考作用。2種生物可以通過比較密碼子使用頻率,分析偏好性差異的大小,從而得出是否適合進行物種間異源表達的結論。濕地松密碼子使用頻率利用在線程序CUSP計算,其余物種的數據源自Codon Usage Database在線數據庫(http://www.kazusa.or.jp/codon/)。
2 結果與分析
2.1 濕地松密碼子組成分析
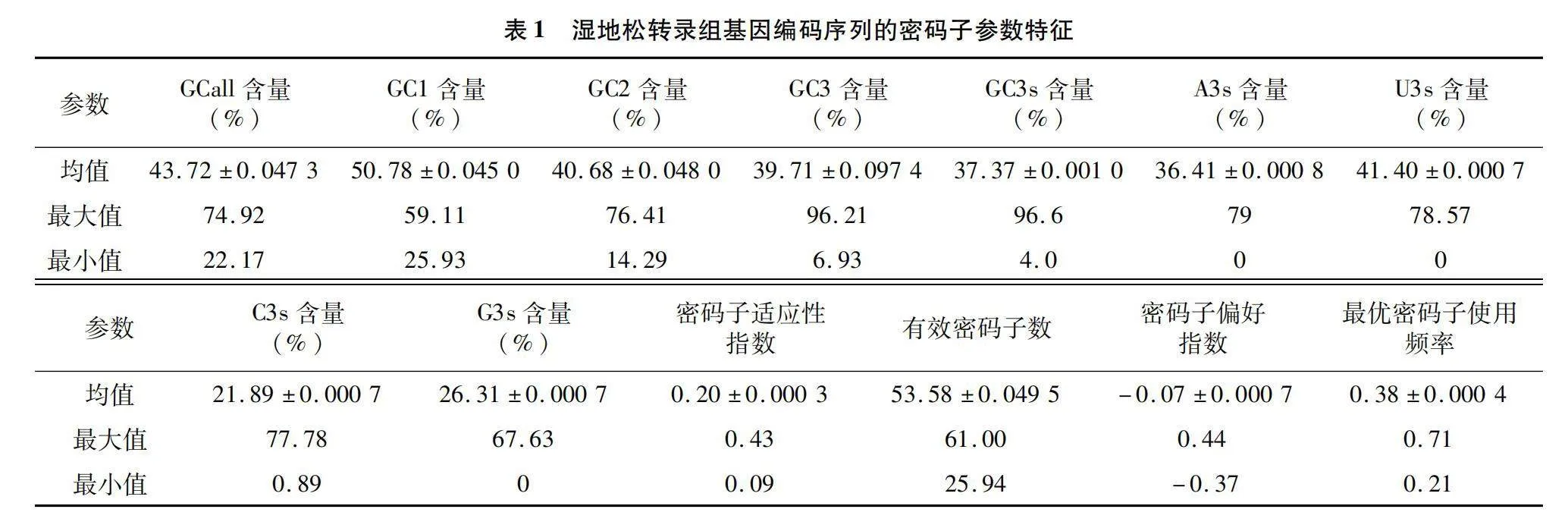
篩選整合序列后用軟件進行參數計算。結果(表1)表明,全部CDS序列的GC1為50.78%、GC2為40.68%、GC3為39.71%,密碼子3個位置的GC含量為GC1>GC2>GC3。平均GC值為43.72%,GC含量在40%~50%之間的CDS序列有10 384條,占總序列的74.53%,GC3含量小于50%的序列有12 205條,占序列總數的87.60%,濕地松轉錄組的U3s(41.40%)>A3s(36.41%)>G3s(26.31%)>C3s(21.89%)。表明濕地松密碼子不同位置的堿基組成存在差異,第3位上是A/U堿基的概率更大。
理論ENc值范圍為20~61,數值越小表示同義密碼子選擇范圍越小、偏好性越強,反之,當ENc=61時,均勻使用密碼子,偏好性較弱。偏好性強弱界定標準為ENc=35[25,30],濕地松密碼子序列的ENc值在25.94~61.00之間,平均值為53.58(表1),ENc≤35的基因有296個,大于35的基因有13 637個,各占比2.12%和97.88%,其中248個ENc值為61個,說明濕地松轉錄組CDS的密碼子偏好性整體水平偏低,僅有少數序列具有較強密碼子偏好性。CAI值較低,介于0.09~0.43之間,均值為0.20,更趨向0,進一步驗證了偏好性弱、基因表達水平較低的觀點。CBI值介于-0.37~0.44之間,Fop值介于0.21~0.71之間。
2.2 同義密碼子相對使用度分析
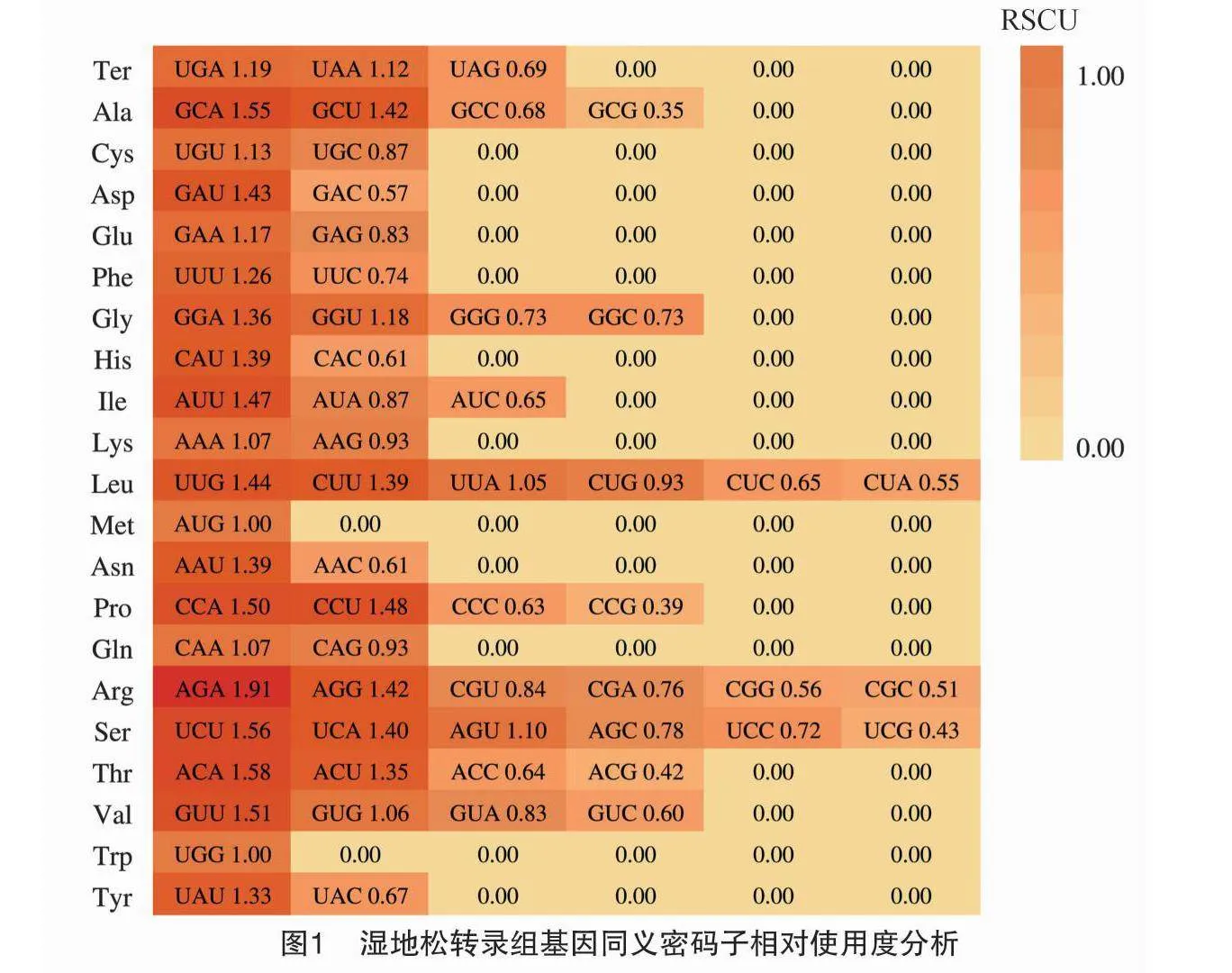
采用CodonW分析濕地松的RSCU值,結果(圖1)顯示,RSCU≥1的密碼子有32個(UGA、UAA、UUA、UCU、UCA、UUG、UUU、UGG、UAU、GCA、GCU、UGU、GAU、GAA、GGA、GGU、CAU、CCA、CCU、AGA、AGU、AUU、ACA、AAA、AGG、CUU、AUG、AAU、CAA、ACU、GUU、GUG),其中結尾為U和A的密碼子分別為15個和12個,以G結尾的密碼子僅有5個,無以C結尾的密碼子,說明濕地松密碼子第3位出現A/U堿基的頻率更高,是濕地松偏好的密碼子。
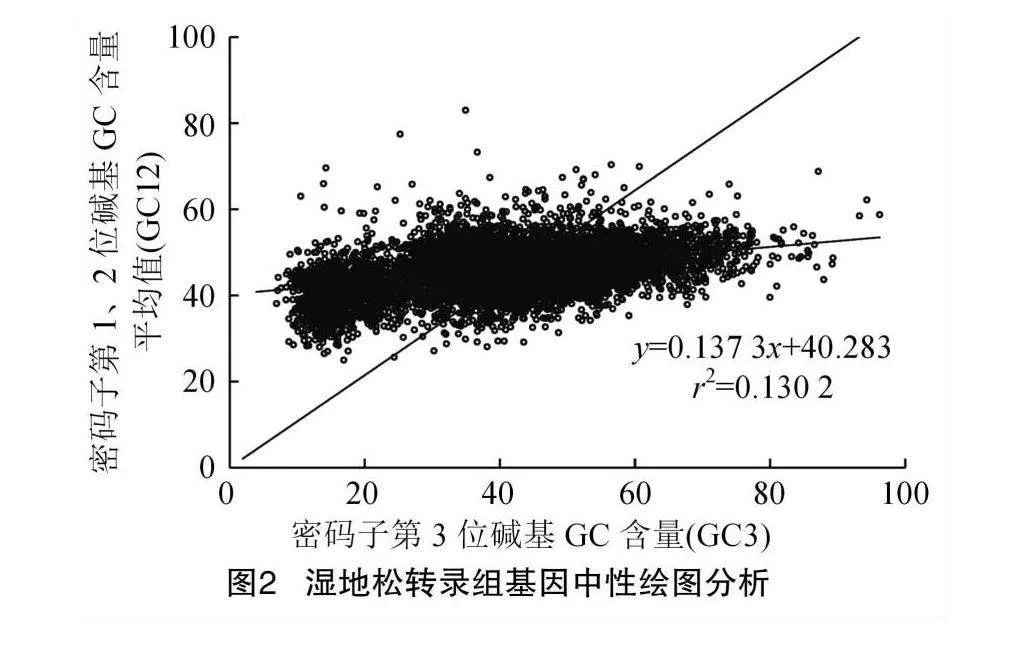
2.3 中性繪圖分析(Neutrality Plot)
開展中性繪圖分析,是為了深入探究導致濕地松出現密碼子使用偏好性的主要原因,結果(圖2)發現,轉錄組GC3含量的分布范圍較廣,在6.93%~96.21%之間,而GC12含量的分布范圍相對較窄,在25.00%~83.02%之間。GC12和GC3含量的決定系數r2=0.130 2,相關性較弱,這表明密碼子受突變壓力的影響程度有限。中性圖的回歸系數(斜率)表示突變的壓力[31]。通過對濕地松進行中性繪圖分析得線性回歸方程y=0.137 3x+40.283,斜率為0.137 3,即突變壓力對濕地松轉錄組密碼子偏好的影響占比僅為13.73%,不足50%,其余占比為自然選擇和其他因素產生的影響。由此可知,濕地松的密碼子使用偏好性是自然選擇壓力主導的,突變因素是次要影響因素。
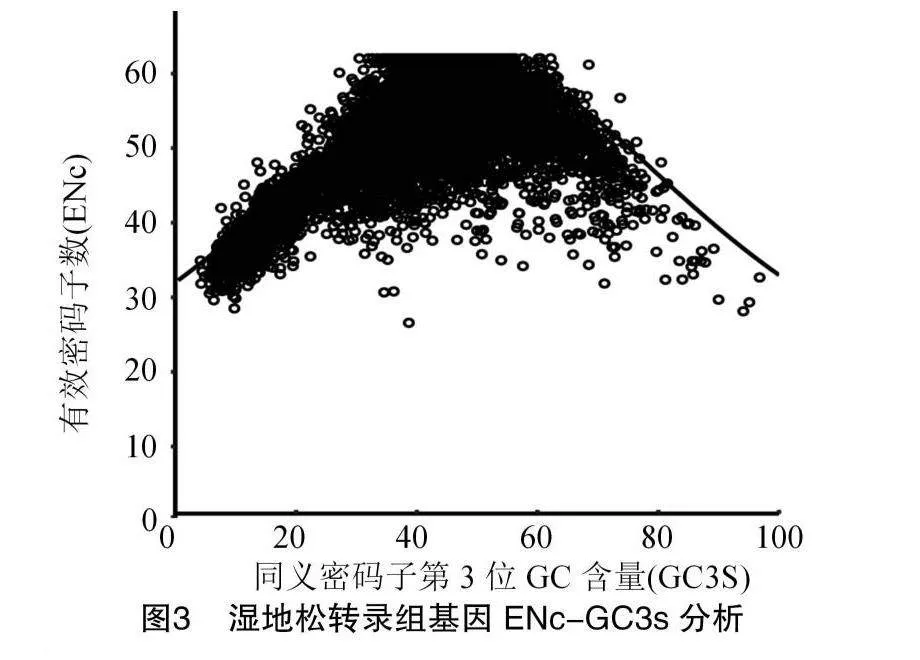
2.4 ENc-GC3s關聯分析
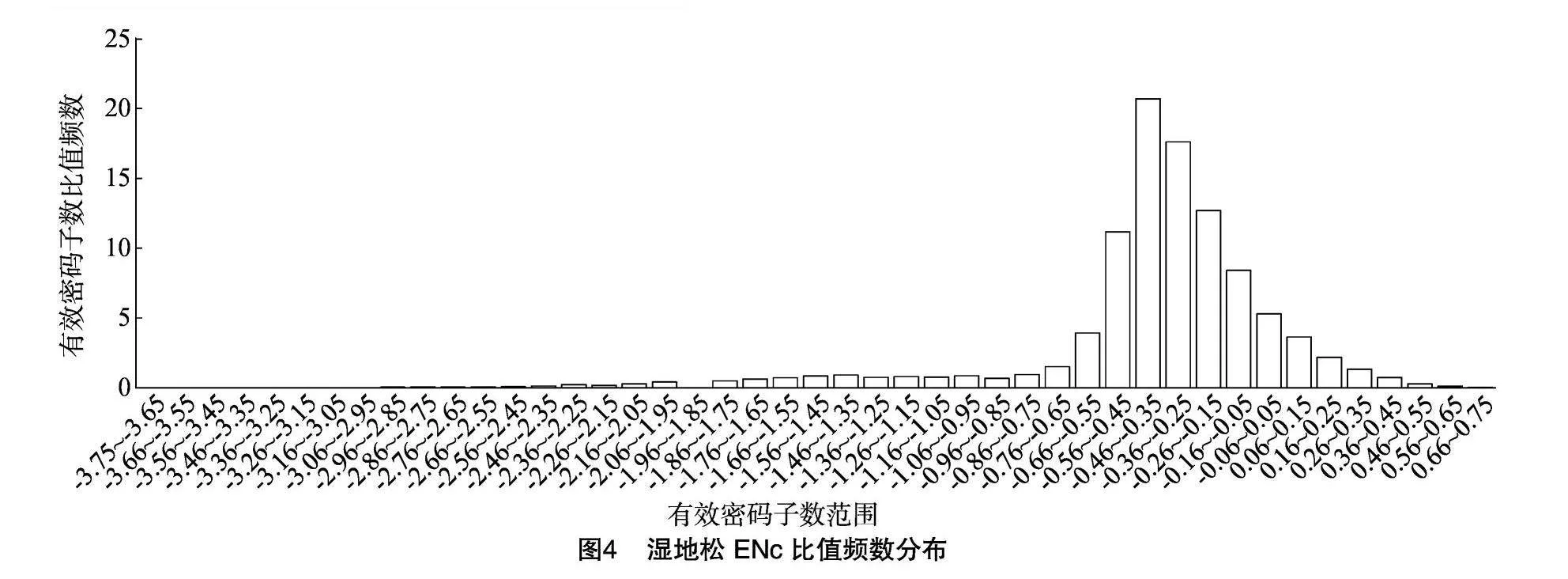
fmwU7P7xQ8yoXd6Ec346fw==ENc-GC3s分析結果(圖3)顯示,基因點大多偏離理論值,也有少數點分布在曲線上或附近。為精準評估實際值與理論值間的差異,進行ENc比值計算,并繪制頻數分布圖,結果(圖4)顯示,ENc比值分布在-3.73~0.72之間,分布在-0.05~0.05之間的基因僅占總數的5.30%。ENc比值在-0.45~-0.35之間的分布占比達到峰值,大多數基因的比值集中在-0.55~-0.15之間,ENc實際值和理論值之間差異較大,大多數理論值高于實際值,表明自然選擇主要影響密碼子偏好性,堿基突變的影響不是關鍵性的,與Neutrality Plot分析結果相同。
2.5 偏倚分析(PR2-plot分析)
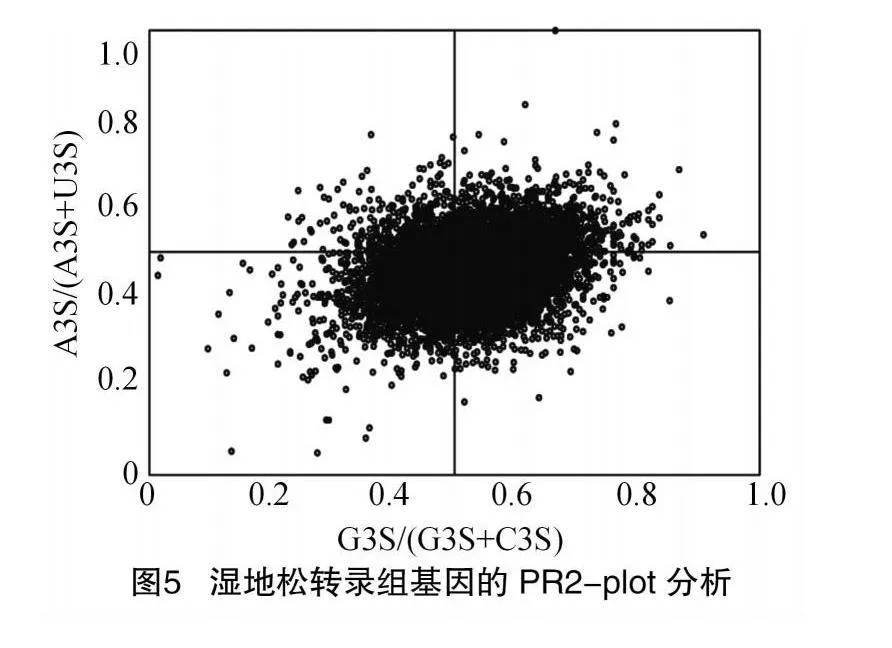
從偏倚分析結果(圖5)可以看出,4個區域內序列點分布不均勻,在垂直方向上,中線下方的基因數量要多于上方;在水平方向上,大部分基因位于中線右側。由此表明,濕地松密碼子中第3位堿基組成頻率U>A、G>C。若基因點均勻分布且堿基使用頻率相等,則突變為主導因素,反之則為圖中情況,非均勻分布,表明自然選擇壓力可能在濕地松密碼子使用偏好中起主導作用,突變壓力是次要因素。
2.6 轉錄組最優密碼子分析
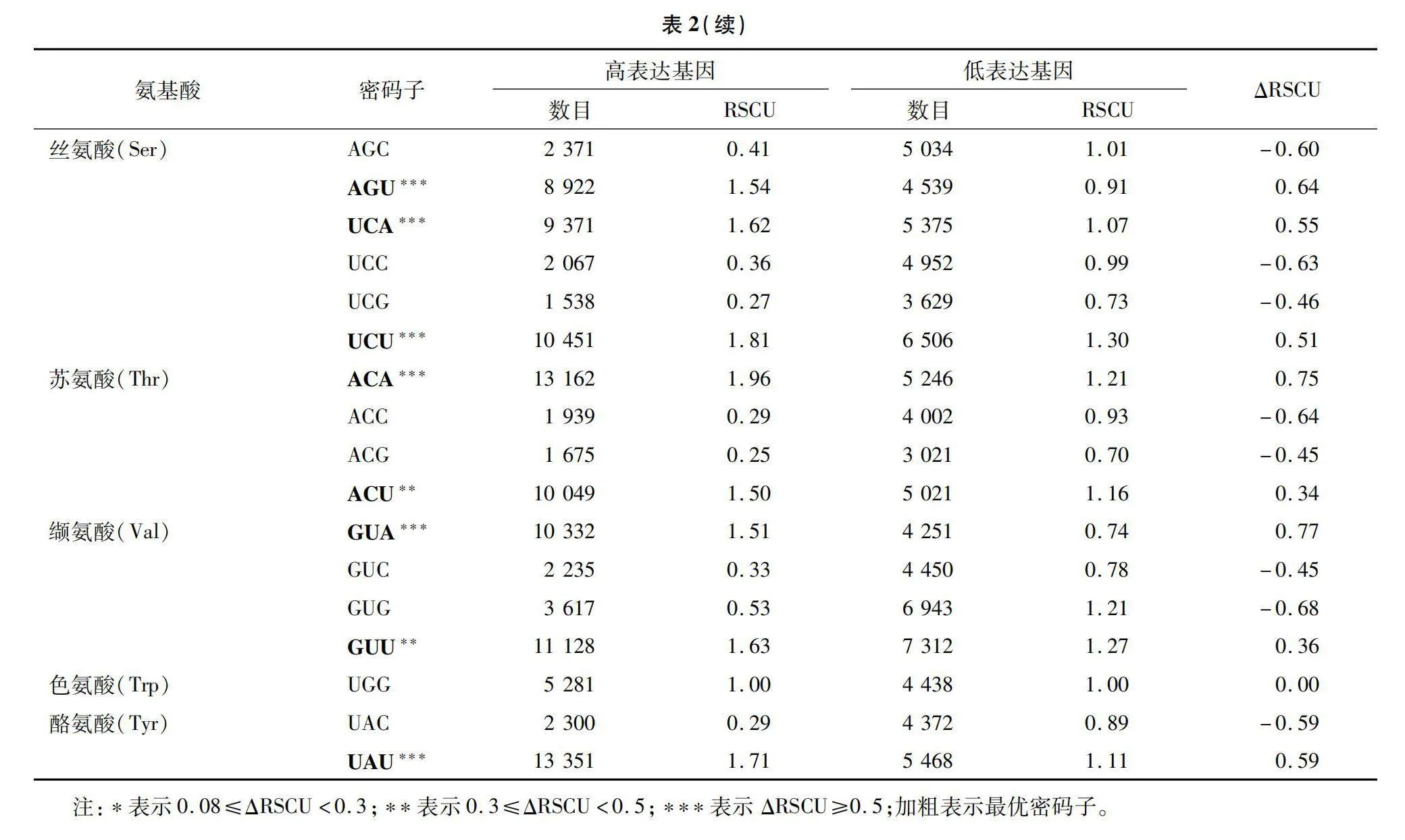
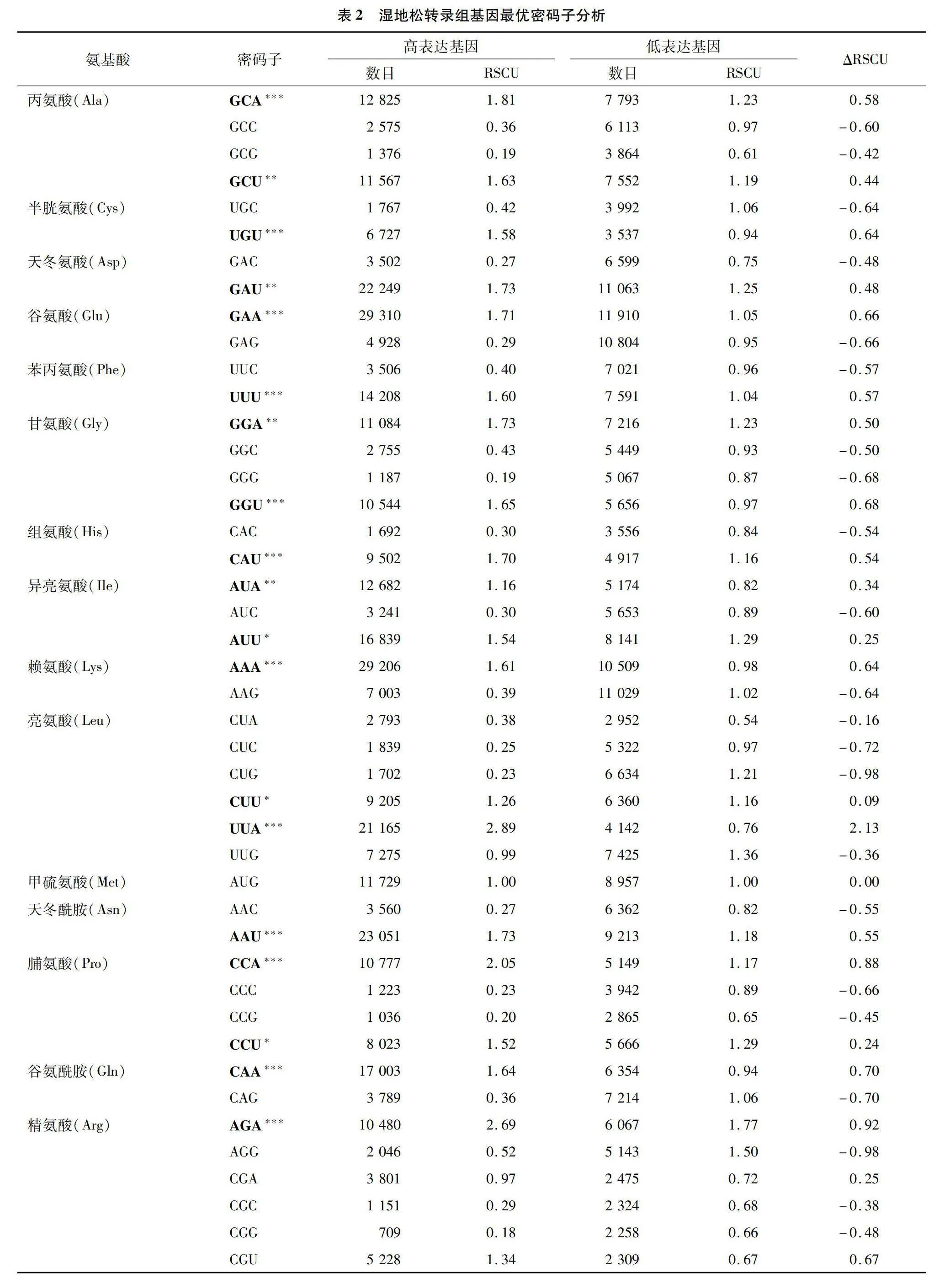
以濕地松轉錄組各基因的ENc值作衡量依據,將數值從小到大排列,首尾各選10%的基因構建高表達和低表達基因組樣本,進行RSCU值計算分析(表2),密碼子的ΔRSCU≥0.08時,被認定為高表達密碼子,共有29個。若某密碼子RSCU=1,表示該密碼子無使用偏性,反之則存在偏好性。有30個密碼子RSCU>1(圖1),為高頻密碼子。同時若密碼子兼具高頻(RSCU值>1)和高表達(ΔRSCU≥0.08)的特點,則被認定為最優密碼子[32],分別有GCA、GCU、UGU、GAU、GAA等25個。其中第3位堿基為A的密碼子有10個、為U的有15個,沒有以C/G結尾的最優密碼子。這與濕地松密碼子組成分析所得的密碼子第3位偏好使用A/U的結論相一致。
2.7 密碼子使用頻率分析
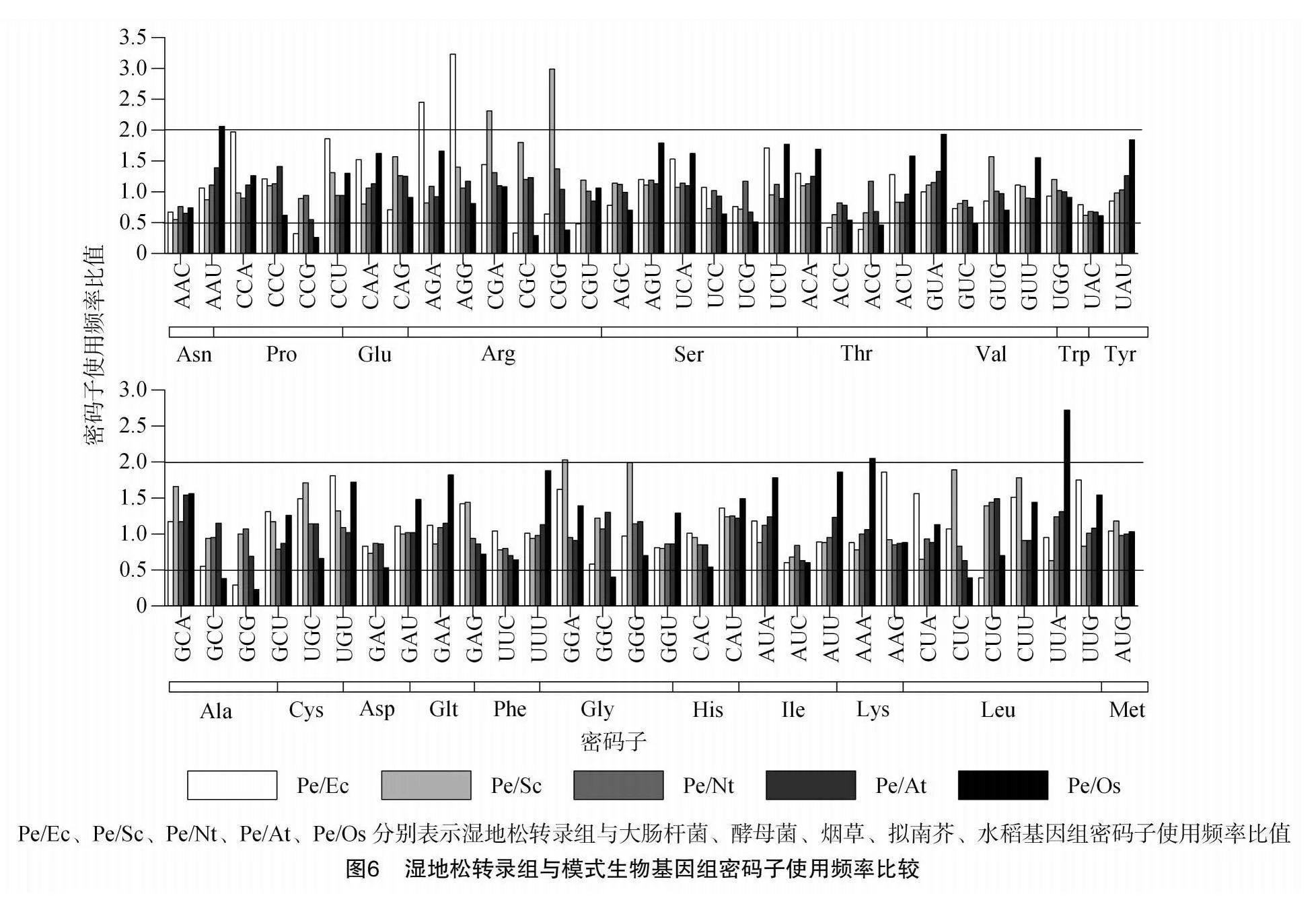
同一密碼子在2個物種間的使用頻率比介于0.5~2.0內,則認為物種間對于該密碼子的使用存在不顯著的偏好差異。對濕地松轉錄組中密碼子使用頻率與Condon Usage Datebases數據庫中公布的大腸桿菌(Escherichia coli,Ec)、擬南芥(Arabidopsis thaliana,At)、煙草(Nicotiana sylvestris L.,Nt)、酵母菌(Saccharomyces cerevisiae,Sc)、水稻(Oryza sativa L.,Os)密碼子使用頻率進行比較,得出的結果(圖6)顯示,與2種微生物相比,濕地松密碼子偏好性與大腸桿菌中存在較大差異的密碼子有9個,與酵母菌僅有3個密碼子存差,可見酵母菌真核表達系統更適合濕地松異源表達,但仍可能需要對部分密碼子如GGA、CGA、CGG進行改造和優化。在與模式植物的比較中,濕地松與雙子葉植物煙草和擬南芥的使用特性差異小,比值范圍分別為0.68~1.44和0.55~1.54;與單子葉植物水稻相比密碼子使用偏好性有12個密碼子存在差異。可見,煙草、擬南芥是濕地松基因進行遺傳轉化功能驗證的理想選擇。
3 討論與結論
植物基因和基因組中密碼子偏好性現象常見,生物體在長期進化和自然適應過程中不是隨機使用同義密碼子,而是對特定密碼子的使用具有傾向性,形成密碼子偏好性,分析密碼子偏好性對于揭示物種或基因的起源、進化關系及突變方式有重要意義[31]。密碼子偏好性分析在精細調控基因表達、轉錄、蛋白折疊等方面有所助力[33],除此之外,對研究基因重組、預測基因功能、發現新基因、設計簡并引物、探究合適的異源表達系統等也有參考價值[34-35]。借助遺傳轉化體系成熟的模式生物進行基因的異源表達是基因功能驗證的常用方法之一。在異源表達和遺傳轉化研究過程中,需要對目標基因及受體的密碼子偏好特點進行分析,根據結果進行目標基因的優化和定向改造,否則可能會由于密碼子使用偏好性的差異導致出現基因甲基化、低水平表達或基因沉默等現象[36]。
植物間存在密碼子偏好性差異,在僅受突變壓力和自然選擇影響的情況下,堿基組成的差異被認定為偏好性模式存在差異的重要原因。GC含量是反映堿基組成的重要指標,且可反映方向性突變的強弱。第3位堿基擺動引起的同義突變不會導致氨基酸種類發生變化,其堿基突變受到的選擇壓力亦小于第1、2位堿基,因此,GC3s常用作密碼子偏好性分析的指標之一[37]。本研究篩選出濕地松轉錄組的13 932條CDS序列,利用軟件進行參數分析得出,GC3s值為37.38%,表明濕地松總體上偏好使用第3位堿基為A/U的密碼子,與高山松[9]、云南油杉[10]、朝倉花椒(Zanthoxylum piperitum DC. var. inerme Makino)[38]的分析結果相一致,也符合大多雙子葉植物的密碼子組成規律[39]。若選擇密碼子第3位堿基偏好為G/C的物種作濕地松基因的異源表達受體則需要適當進行密碼子定向優化。
在密碼子偏好性分析中,ENc值被認為是最具有參考價值的指標之一[30]。本研究以ENc值為基礎,按高頻高表達標準篩選最優密碼子,在濕地松基因組中確定了25個最優密碼子,其中以A結尾的密碼子有10個,U結尾的密碼子有15個,無以G/C結尾的最優密碼子,充分說明濕地松偏好選用A/U結尾的密碼子。這與在大黃(Rheum officinale Baill.)[40]和香樟[41]等植物上的研究結果相類似。研究結果也進一步表明,異源基因同義密碼子第3位堿基偏好為A/U時,在濕地松中可能會獲得高表達效率,zttDf/P5YeP8eaGo9loD4w==但還需要進一步開展試驗驗證。前人研究認為,在排除自然環境壓力的前提下,突變壓力的方向會影響密碼子堿基及末尾堿基的組成[11,36],由此推斷,濕地松在進化過程中密碼子可能從GC到AU方向發生突變。
突變和自然選擇是影響密碼子偏好性的主要因素,本研究進行濕地松轉錄組數據的中性繪圖、PR2-plot繪圖和ENc-GC3s關聯分析,結果均表明,大多數濕地松基因密碼子存在使用偏好性的關鍵原因是自然選擇,少數基因受突變壓力的影響。這與在香樟和石榴上進行的轉錄組基因密碼子的分析結果[24]相一致。在異源表達研究中,密碼子偏好性分析能為外源基因和受體匹配提供參考,選擇密碼子偏好模式差異小的物種作異源受體,對于外源基因轉錄、翻譯效率的提高及獲取高表達量有益[42]。根據本試驗結果,濕地松與擬南芥、煙草密碼子之間的偏好性差異較小,密碼子使用頻率比值處于0.5~2.0范圍內,雙子葉植物擬南芥、煙草均可作濕地松基因進行遺傳轉化的受體,進一步表明濕地松基因可能更適合在擬南芥、煙草等雙子葉模式植物中進行功能基因的驗證。如朱靈芝和李榮分別將馬尾松萜類化合物合成途徑關鍵酶基因PmDXR和PmDXS轉入擬南芥中,結果發現,超表達擬南芥的1-脫氧-D-木酮糖-5-磷酸還原異構酶(1-deoxy-D-xylulose-5-phosphate reductoisomerase,DXR)活性、1-脫氧-D-木酮糖-5-磷酸合酶(1-deoxy-D-xylulose-5-phosphate synthase,DXS)活性以及葉綠素a、葉綠素b、類胡蘿卜素含量較野生型均有提高[43-44]。
在后續研究中,可進一步與其他針葉樹種密碼子進行比較,精細分析密碼子的區域分布,且可針對萜類合成途徑中的某一關鍵酶基因做密碼子偏好性分析,不僅能了解物種間的遺傳進化規律,還能為基因的外源表達選擇最佳宿主。此外,現代高通量測序技術在不斷的發展,將其與濕地松生長、松脂組分和產脂量等相關轉錄本間密碼子偏好性結合進行分析,能夠為濕地松高產脂性狀形成的遺傳機制研究提供重要的依據[11]。
本研究初步分析濕地松轉錄組的密碼子偏好性及其異源表達模式、表達載體的選擇,為之后濕地松基因功能驗證尋找合適的轉化載體,逐步完善濕地松的遺傳轉化體系提供了參考,也對后續利用基因工程技術進行濕地松高產脂遺傳改良具有重要的指導意義。
本研究表明,濕地松基因組平均GC含量、GC3值、GC3s的值分別為43.72%、39.71%、37.37%。整體密碼子偏好性較弱,偏好以A/U堿基結尾的密碼子。利用高頻率高表達的篩選標準選出25個最優密碼子,均以A/U結尾。中性繪圖、PR2-plot繪圖分析和ENc-GC3s關聯分析結果均表明,濕地松基因組偏好性受多種因素共同影響,其中自然選擇是主要原因。酵母菌真核表達系統較大腸桿菌而言,更適合濕地松基因的異源表達,遺傳轉化體系較為完善的模式植物煙草是濕地松基因進行遺傳轉化功能驗證的理想選擇。
參考文獻:
[1]秦斗文,徐庭亮,閆京艷,等. 柔毛郁金香葉綠體基因組密碼子偏好性分析[J]. 江蘇農業科學,2023,51(22):41-47.
[2]覃興化,張麒功,汪其雙,等. 梔子葉綠體基因組密碼子偏好性分析[J]. 福建農林大學學報(自然科學版),2023,52(5):632-639.
[3]West S E,Iglewski B H. Codon usage in Pseudomonas aeruginosa[J]. Nucleic Acids Research,1988,16(19):9323-9335.
[4]蘇 惠,李永光,譚文雍,等. 大腸桿菌gdhA基因的密碼子偏好性分析及優化[J]. 基因組學與應用生物學,2015,34(3):521-529.
[5]雷佳欣,張麗娟,高丹丹,等. 文冠果基因組密碼子偏好性分析[J]. 西北林學院學報,2023,38(4):104-110.
[6]Zhao F Z,Zhou Z P,Dang Y K,et al. Genome-wide role of codon usage on transcription and identification of potential regulators[J]. Proceedings of the National Academy of Sciences of the United States of America,2021,118(6):e2022590118.
[7]Parvathy S,Udayasuriyan V,Bhadana V. Codon usage bias[J]. Molecular Biology Reports,2021,49:539-565.
[8]原曉龍,陳 偉,王 毅,等. 思茅松葉綠體基因組密碼子偏好性分析[J]. 基因組學與應用生物學,2021,40(3):1278-1283.
[9]李江飛,李亞麒,唐軍榮,等. 高山松葉綠體基因組密碼子偏好性模式[J]. 生物學雜志,2023,40(1):52-59.
[10]李江飛,王 瑜,顏廷雨,等. 云南油杉葉綠體基因組密碼子偏好性分析[J]. 中南林業科技大學學報,2022,42(4):30-39.
[11]賴瑞聯,馮 新,陳 瑾,等. 橄欖轉錄組密碼子使用偏好性及其影響因素[J]. 核農學報,2019,33(1):31-38.
[12]朱沛煌,陳 妤,朱靈芝,等. 馬尾松轉錄組密碼子使用偏好性及其影響因素[J]. 林業科學,2020,56(4):74-81.
[13]Yi M,Zhang L,Cheng Z S,et al. Identification of key genes for oleoresin biosynthesis in high and low oleoresin-yielding slash pine based on transcriptome analysis[J]. Forests,2022,13(8):1337.
[14]莊偉瑛,歐陽道明,范國榮,等. 濕地松松脂中單萜類組分研究續報[J]. 江西農業大學學報,2009,31(3):388-392.
[15]Neis F A,de Costa F,de Almeida M R,et al. Resin exudation profile,chemical composition,and secretory canal characterization in contrasting yield phenotypes of Pinus elliottii Engelm[J]. Industrial Crops and Products,2019,132:76-83.
[16]莊偉瑛,張玉英,鄒元熹. 高產脂濕地松選擇和相關因子的分析[J]. 江西農業大學學報,2007,29(1):55-60,65.
[17]李彥杰,欒啟福,沈丹玉,等. 濕地松自由授粉家系松脂組分遺傳變異研究[J]. 林業科學研究,2012,25(6):773-779.
[18]Lai M,Zhang L,Lei L,et al. Inheritance of resin yield and main resin components in Pinus elliottii Engelm.at three locations in southern China[J]. Industrial Crops and Products,2020,144:112065.
[19]Yi M,Jia T,Dong L M,et al. Resin yield in Pinus elliottii Engelm. is related to the resin flow rate,resin components and resin duct characteristics at three locations in southern China[J]. Industrial Crops and Products,2021,160:113141.
[20]冷春暉,張 露,易 敏,等. 濕地松產脂量與生長及樹冠性狀多點遺傳相關及通徑分析[J]. 核農學報,2020,34(7):1598-1605.
[21]雷 蕾,潘顯強,張 露,等. 濕地松左旋β-蒎烯合成酶基因PeTPS-(-)BPin的同源克隆及生物信息學分析[J]. 江西農業大學學報,2015,37(2):205-211.
[22]張文娟,谷振軍,胡 珊,等. 濕地松木質部和針葉松脂合成基因分析[J]. 熱帶亞熱帶植物學報,2023,31(4):531-540.
[23]雷 蕾. 濕地松TPS基因同源克隆及其與產脂性狀的關聯分析[D]. 南昌:江西農業大學,2014.
[24]張太奎,起國海,葉紅蓮,等. 石榴轉錄組密碼子使用偏向性[J]. 園藝學報,2017,44(4):675-690.
[25]盧珍紅,原曉龍,李紳崇,等. 基于轉錄組序列的非洲菊密碼子偏好性分析[J]. 西南林業大學學報(自然科學),2023,43(5):77-86.
[26]Sharp P M,Tuohy T M F,Mosurski K R. Codon usage in yeast:cluster analysis clearly differentiates highly and lowly expressed genes[J]. Nucleic Acids Research,1986,14(13):5125-5143.
[27]郝丙青,夏瑩瑩,葉 航,等. 香花油茶葉綠體基因組密碼子偏好性分析[J]. 中南林業科技大學學報,2022,42(9):178-186.
[28]吉德娟,王占林.霸王密碼子偏好性分析[J]. 分子植物育種,2023,21(20):6705-6713.
[29]Chakraborty S,Yengkhom S,Uddin A. Analysis of codon usage bias of chloroplast genes in Oryza species:codon usage of chloroplast genes in Oryza species[J]. Planta,2020,252(4):67.
[30]Wright F. The ‘effective number of codons’ used in a gene[J]. Gene,1990,87(1):23-29.
[31]趙春麗,彭麗云,王 曉,等. 莧菜AtGAI基因密碼子偏好性與進化分析[J]. 中國農業大學學報,2019,24(12):10-22.
[32]梁皓輝,符虹宇,黎釗坪,等. 小球藻葉綠體基因組密碼子偏好性分析[J]. 分子植物育種,2020,18(17):5665-5673.
[33]任桂萍,董瓔瑩,黨云琨.密碼子中的密碼:密碼子偏好性與基因表達的精細調控[J]. 中國科學(生命科學),2019,49(7):839-847.
[34]Liu S S,Qiao Z Q,Wang X M,et al. Analysis of codon usage patterns in “Lonicerae flos” (Lonicera macranthoides Hand.-Mazz.) based on transcriptome data[J]. Gene,2019,705:127-132.
[35]Deb B,Uddin A,Chakraborty S. Codon usage pattern and its influencing factors in different genomes of hepadnaviruses[J]. Archives of Virology,2020,165(3):557-570.
[36]陳義挺,賴瑞聯,馮 新,等. 獼猴桃查爾酮合成酶基因CHS密碼子偏好性分析[J]. 福建農業學報,2017,32(11):1213-1217.
[37]陳 悅,仇學文,李 鈺,等. 辣椒和番茄PIF基因家族密碼子偏好性分析[J]. 分子植物育種,2023,21(17):5602-5610.
[38]王 超,趙懿琛,趙德剛.朝倉花椒轉錄組密碼子偏好性分析[J]. 分子植物育種,2020,18(21):6989-6996.
[39]Murray E E,Lotzer J,Eberle M. Codon usage in plant genes[J]. Nucleic Acids Research,1989,17(2):477-498.
[40]高夢琦,鄒建珍,霍小位,等. 基于轉錄組數據分析藥用大黃的密碼子使用偏好性[J]. 中草藥,2021,52(20):6344-6349.
[41]梁曉靜,朱昌叁,李開祥,等. 香樟轉錄組基因密碼子偏好性分析[J]. 廣西植物,2021,41(12):2077-2083.
[42]朱靈芝,朱沛煌,李 榮,等. 馬尾松PmDXR基因密碼子偏好性分析[J]. 林業科學研究,2021,34(2):102-113.
[43]朱靈芝.馬尾松PmDXR基因的克隆與功能分析[D]. 南京:南京林業大學,2021.
[44]李 榮.馬尾松PmDXS基因克隆及功能鑒定[D]. 南京:南京林業大學,2021.
