芒果可可毛色二孢咪鮮胺抗藥性菌株的轉錄組分析
2024-10-09 00:00:00劉錦霖李鵬聲楊葉
熱帶作物學報 2024年9期


關鍵詞:芒果;可可毛色二孢;咪鮮胺;轉錄組;解毒代謝酶
中圖分類號:S436.67 文獻標志碼:A
芒果(Mangifera indica L.)是我國重要的熱帶經(jīng)濟作物,海南地區(qū)的芒果產(chǎn)量和種植面積均位居我國前列。芒果果肉香味濃郁,除鮮食外還可制作果醬和飲品,深受消費者喜愛。芒果蒂腐病是危害芒果品質(zhì)、導致采后果實嚴重腐爛的重要病害。可可毛色二孢(Lasiodiplodia theobromae)是引發(fā)芒果蒂腐病的主要病原菌之一[1]。該病原菌是一種全球性病原真菌,能夠在芒果等多種熱帶果樹及林木作物上侵染為害,造成采后果實蒂腐病和田間樹木、枝干的枯死等病害[1]。此外,該菌具有潛伏侵染的特性,病菌可在幼果期侵染并在果實中潛伏,果實成熟后病斑迅速擴展,發(fā)生褐變并散發(fā)出酸臭味,在芒果采后貯藏和運輸?shù)倪^程中爆發(fā),嚴重影響芒果的品質(zhì)和經(jīng)濟價值[2]。目前防治芒果病害的方法主要包括物理防治、生物防治與化學防治,其中化學防治是最常用的方式,異菌脲、多菌靈、咪鮮胺等殺菌劑能夠有效防治芒果蒂腐病[3-4]。然而,隨著藥劑施用量持續(xù)增加,使得病原菌的選擇壓力持續(xù)增大,單一靶標位點的殺菌劑已經(jīng)產(chǎn)生了嚴重的抗藥性。據(jù)報道,芒果可可毛色二孢對多菌靈、嘧菌酯抗性已經(jīng)達到十分嚴重的水平[5-6]。
大量研究表明,害蟲、雜草及病原菌對農(nóng)藥的抗性機制主要包括靶標抗性機制和非靶標抗性機制[6]。靶標抗性主要是殺菌劑靶標基因發(fā)生突變所致,致使藥劑與作用位點酶的親和性降低。而非靶標抗性主要是增加藥劑外排作用、解毒作用和對藥劑的通透性下降所致,又稱為代謝抗性,增加藥劑外排與代謝解毒常常協(xié)同發(fā)揮作用[7]。以細胞色素P450 介導的微粒體多功能氧化酶、谷胱甘肽轉移酶等代謝酶系為主體的解毒系統(tǒng)能夠把農(nóng)藥分解成毒性更低、親水性更強的物質(zhì),便于排出體外[8]。代謝抗性的發(fā)生不僅會提升抗藥性水平,也有可能導致交互抗性及多藥抗性的發(fā)生[9]。目前,代謝抗性的研究主要集中在害蟲及雜草抗藥性的產(chǎn)生機制,病原真菌的代謝抗性研究尚處于初步階段。本研究擬在芒果上分離獲得的咪鮮胺抗藥性菌株為供試材料,通過轉錄組測序及差異表達基因的分析,挖掘與代謝抗性相關的基因,對代謝抗性的機制進行初步探析,為后續(xù)代謝抗性機制的深入研究提供依據(jù)。研究芒果可可毛色二孢抗藥性產(chǎn)生機制,是延緩田間抗藥性發(fā)展的關鍵所在,對提升芒果蒂腐病防治效果及提高芒果產(chǎn)量和品質(zhì)具有重要指導價值。
1 材料與方法
1.1 材料
1.1.1 供試菌株 供試3 株芒果蒂腐病可可毛色二孢菌株HL02、M108 和DF04 由海南大學植物保護學院殺菌劑生物學實驗室分離鑒定并保存。將菌株接種到馬鈴薯葡萄糖瓊脂(PDA)培養(yǎng)基上,于28 ℃黑暗中培養(yǎng)3 d 后備用。
1.1.2 供試藥劑 97%咪鮮胺購自海南正業(yè)中農(nóng)高科股份有限公司;總RNA 提取試劑盒購自天根生化科技(北京)有限公司;DL2000 DNA Marker、RNA逆轉錄試劑盒、ChamQ Universal qPCR SYBRMaster Mix 購自南京諾唯贊生物科技有限公司;DNA 1000 assay Kit 購自安捷倫科技有限公司。
1.2 方法
1.2.1 咪鮮胺敏感性測定 采用菌絲生長速率法測定3株可可毛色二孢菌株對咪鮮胺的敏感性。咪鮮胺用少量丙酮溶解并配制成1×103 mg/L 的母液備用。在PDA 培養(yǎng)基中加入不同量的殺菌劑母液,配制得到不同濃度的含藥培養(yǎng)基平板,最終試驗濃度為0、0.2、0.8、3.2、12.8、51.2 mg/L。對照培養(yǎng)基中加入等量的溶劑。每處理3個重復,試驗重復3 次。從培養(yǎng)3 d 的菌落邊緣取菌餅(直徑5 mm)轉移到含藥PDA平板上,28 ℃培養(yǎng)36 h 后,測量菌落直徑并計算抑制率。將供試濃度轉換為濃度對數(shù),抑制率轉換為幾率值,使用Probit 分析求回歸方程y=a+bx,計算半數(shù)有效濃度EC 值。
1.2.2 可可毛色二孢的RNA 提取及轉錄組測序從培養(yǎng)3 d 的菌落邊緣取菌餅(直徑5 mm)轉移到100 mL 馬鈴薯葡萄糖液體培養(yǎng)(PDB),每瓶培養(yǎng)液接種5 個菌餅,于145 r/min 震蕩培養(yǎng)24 h;然后加入咪鮮胺使其終濃度為10 mg/L,再培養(yǎng)12 h 后取出每個樣品的菌絲,使用總RNA 提取試劑盒,嚴格按照說明書提取樣品的總RNA。采用1%瓊脂糖凝膠電泳及微量紫外分光光度計檢測其純度和濃度。每個處理3 個重復樣品,將質(zhì)檢合格的樣品混合,然后設3 個生物學重復。用Oligo(dT)磁珠從上述RNA 樣品中富集mRNA 并進行超聲處理,以片段化的mRNA 為模板合成cDNA 第一鏈,隨后用RNaseH 降解RNA 鏈,并在DNA polymerase Ⅰ體系下,在第二鏈合成時加入dNTPs 補齊雙鏈DNA的末端。在純化后的雙鏈DNA 的兩端各加上1 個A 堿基后與末端帶有T 堿基的接頭進行連接,使用AMPure XP beads磁珠篩選200 bp 左右的cDNA,進行PCR 擴增并再次純化后獲得cDNA 文庫,選擇DNA 1000assay Kit 對文庫進行質(zhì)檢合格后,使用IlluminaHiSeq TM 2500 測序儀測序。利用HISAT2 將雙端測序得到的序列比對到參考基因組( NCBI_ASM982982v1)。
1.2.3 轉錄組數(shù)據(jù)分析和差異表達基因篩選 測序原始數(shù)據(jù)raw reads 利用fastp 過濾和質(zhì)控得到cleanreads[10]。使用短reads 比對工具Bowtie 2將cleanreads 比對到可可毛色二孢的核糖體數(shù)據(jù)庫[11],在不允許錯配情況下去除比對上核糖體的reads,將保留下來的unmapped reads 用于后續(xù)轉錄組分析。利用DESeq2軟件比較不同處理的樣品與對照樣品,從而鑒定差異基因[12]。以FDR<0.05 且|log2(FC)|>1 為標準篩選識別顯著變化的差異表達基因( DEG ), 并對差異基因進行GO( http://www.geneontology.org/ ) 功能分析和KEGG(http://www.genome.jp/kegg/)通路富集分析。另外,使用HMMER3.0 軟件和BLASTP 程序比對含有細胞色素P450 蛋白結構域、谷胱甘肽轉移酶蛋白結構域和ABC 轉運蛋白結構域的序列,篩選后比對到轉錄組差異基因篩選結果中,結合差異基因功能和通路富集分析結果篩選與抗藥性相關的解毒代謝基因。
1.2.4 差異表達基因的定量驗證3個菌株分別設10 mg/L 咪鮮胺處理組和未處理組(對照),每個樣品設置3個獨立的生物學重復。提取芒果可可毛色二孢菌株總RNA 并使用RNA 逆轉錄試劑盒合成cDNA,使用ChamQ Universal qPCR SYBRMaster Mix 配制熒光定量檢測體系,以actin 基因作為內(nèi)參基因測定差異基因表達量。使用PrimerPremier 5 軟件設計特異性引物(表1),所有引物均由楠山科技有限公司(海南)合成。qPCR 反應體系:2×ChamQ Universal qPCR SYBR Master Mix10 μL、10 μmol/L 上/下游引物各0.4 μL、cDNA 模板1 μL、ddHO 8.2 μL。反應程序:95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火30 s,循環(huán)42 次。每個處理3個生物學重復和3個技術性重復。
1.3 數(shù)據(jù)處理
使用2?ΔΔCT 方法計算基因相對表達量[13]。使用GraphPad Prism 軟件進行數(shù)據(jù)分析,采用LSD法進行差異顯著性檢驗。
2 結果與分析
2.1 咪鮮胺處理對菌株的作用
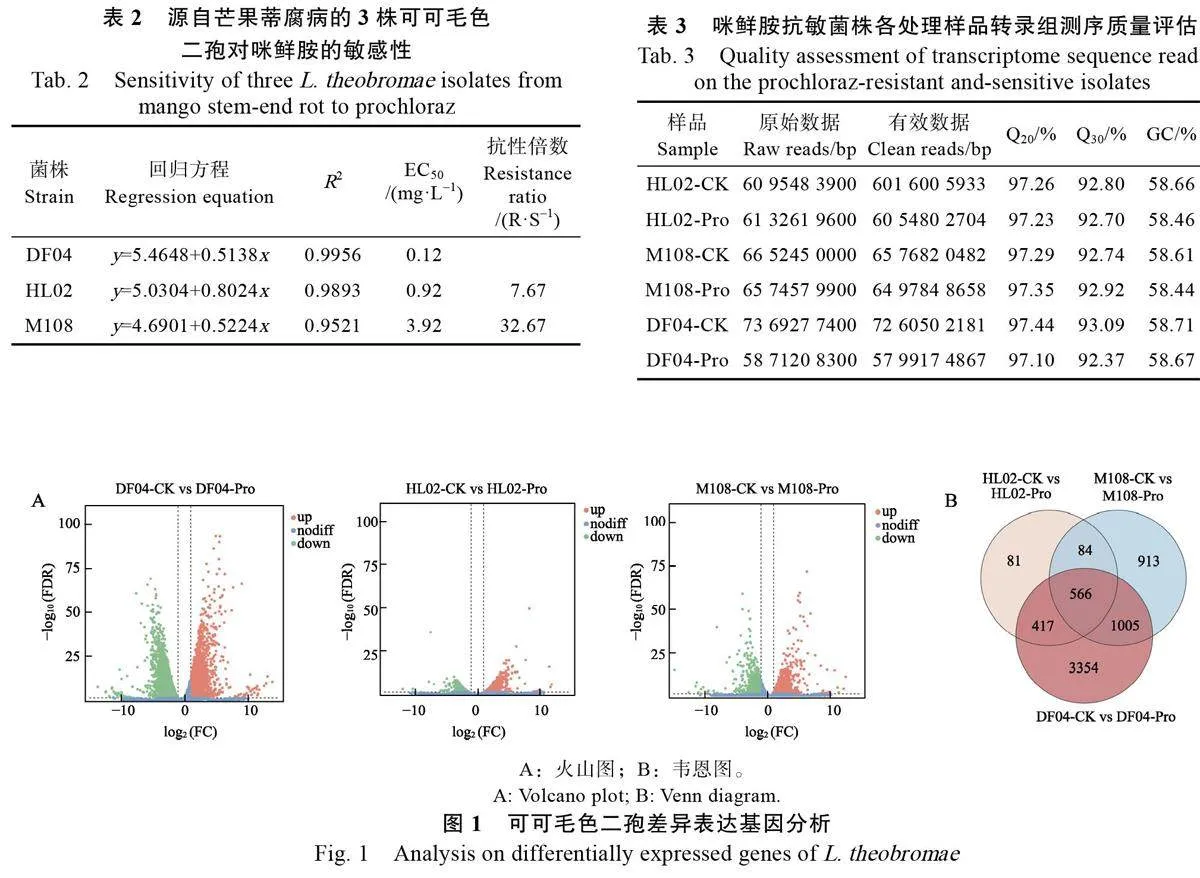
供試3個菌株對咪鮮胺的EC50 值在0.12~3.92 mg/L 之間,與敏感菌株DF04 相比,抗性菌株HL02和M108的抗性倍數(shù)分別為7.67倍和32.67倍(表2)。咪鮮胺敏感菌株的菌絲生長受藥劑的抑制,除了菌落大小的差異,菌絲也較抗性菌株稀薄。
2.2 轉錄組測序數(shù)據(jù)質(zhì)量分析
3個菌株分別設10mg/L 咪鮮胺處理組和未處理組,共6個樣品,每個樣品設置3 個獨立的生物學重復。過濾低質(zhì)量數(shù)據(jù)后,從8 個文庫中生成了超過50 億個clean reads,3 個重復的平均Q介于97.10%~97.443%之間,Q介于92.375%~93.09%之間,GC 含量介于58.46~58.71 之間(表3)。質(zhì)控后的clean reads 數(shù)超過3500 萬條,其中超過90.81%(unique map)比對到參考基因組唯一位置上,全部的可以定位到基因組上的reads數(shù)量及占有效reads 比例(total mapped)超過91.40%。以上結果表明轉錄組測序數(shù)據(jù)質(zhì)量高,可用于后續(xù)生物信息學分析。
2.3 差異表達基因表達分析
以FDR<0.05 且|log2(FC)|>1 為條件進行差異表達基因的篩選。差異基因火山圖數(shù)據(jù)顯示(圖1A),10 mg/L 咪鮮胺處理(Pro)與未處理(CK)樣品間的基因表達水平及差異基因數(shù)量存在差異:經(jīng)10 mg/L 咪鮮胺處理后的抗性菌株HL02、M108 分別有609、1570 個顯著上調(diào)表達基因,539、998 個顯著下調(diào)表達基因;而敏感菌株DF04有2026 個顯著上調(diào)表達基因,3316 個顯著下調(diào)表達基因,敏感菌株的差異基因數(shù)量顯著高于抗藥菌株。韋恩圖顯示(圖1B),3 個菌株添加咪鮮胺處理前、后共有566 個差異表達基因。
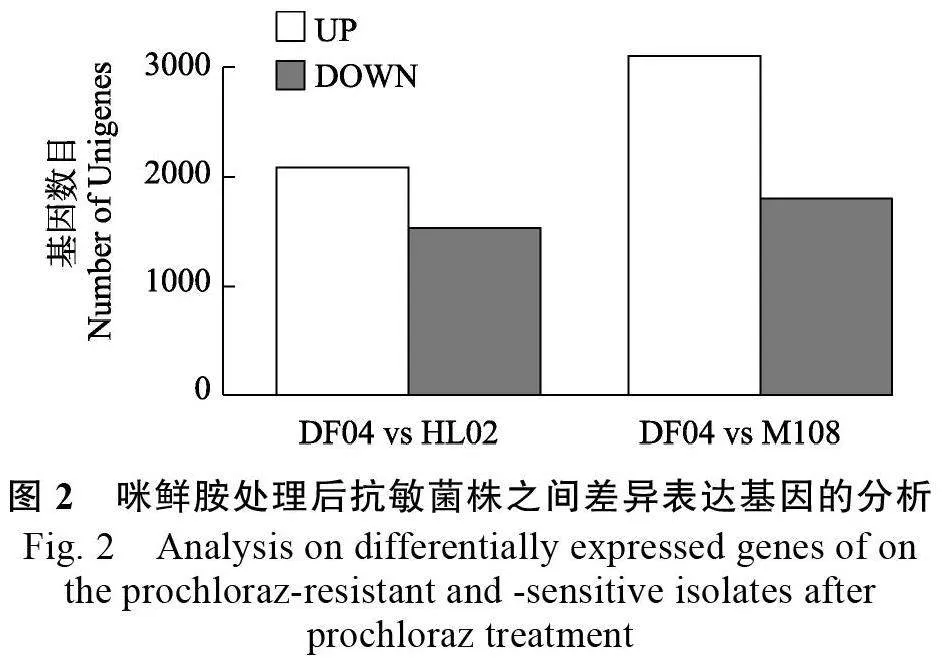
針對咪鮮胺處理后抗性菌株與敏感菌株之間的差異表達基因進行分析,結果表明,抗性菌株HL02、M108 分別有2078、3096 個上調(diào)表達基因,1527、1793 個下調(diào)表達基因(圖2),上調(diào)表達基因數(shù)量遠大于下調(diào)表達基因。
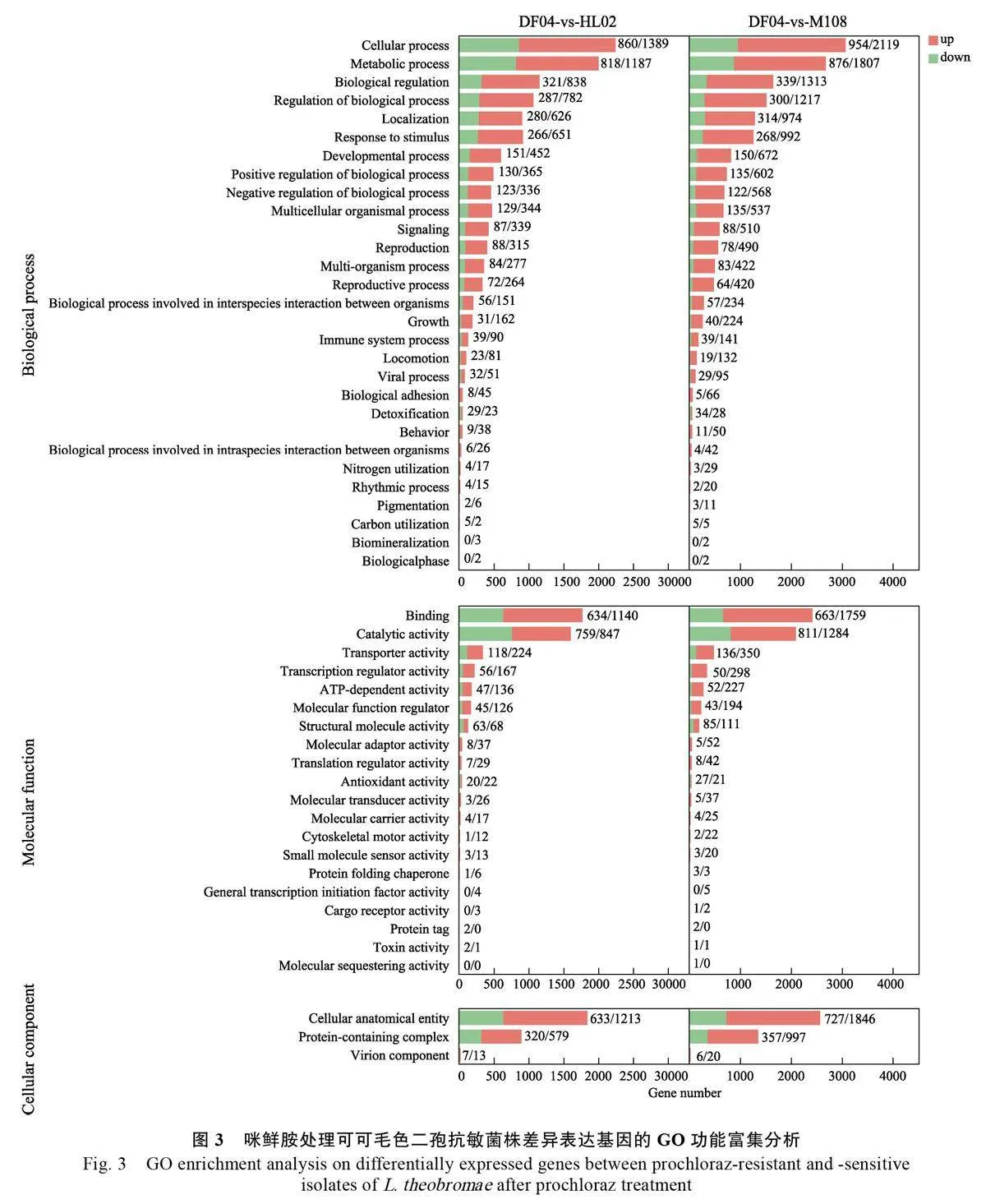
2.4 咪鮮胺caWio5UGbH3r9GPI2F1ULw==處理后的差異表達基因GO富集分析
GO功能富集分析發(fā)現(xiàn),咪鮮胺處理后的抗性和敏感菌株的差異表達基因分別集中在生物學進程(biological process)、分子功能(molecularfunction)及細胞組分(cellular component)方面,其中,基因功能主要集中在27 個term 中,它們的差異表達基因數(shù)均大于100。生物學進程中富集的差異基因數(shù)量最多,主要富集在細胞過程(cellular process)和代謝過程(metabolic process),其中,DF04 vs HL02 中分別有2249(18%)、2005(16%)個DEGs,DF04 vs M108 中分別有3073(17%)、2683(15%)個DEGs。分子功能進程中差異表達基因主要富集在結合(binding)、催化活性(catalytic activity),DF04 vs HL02 中分別有1774(38%)、1606(35%)個DEGs,DF04 vsM108 中分別有2422(38%)、2095(33%)個DEGs。細胞組分進程中的差異表達基因顯著富集在細胞結構體(cellular anatomical entity)和含蛋白質(zhì)復合物(protein-containing complex)中,DF04 vs HL02中分別有1846(67%)、899(32%)個DEGs,DF04vs M108 中分別有2573(65%)、1354(34%)個DEGs(圖3)。2 個抗性菌株HL02、M108 與敏感菌株DF04 之間的差異表達基因分類趨勢表現(xiàn)一致,但DF04 vs M108 比對的差異表達基因高于DF04 vs HL02。
2.5 咪鮮胺處理后差異表達基因KEGG信號通路富集
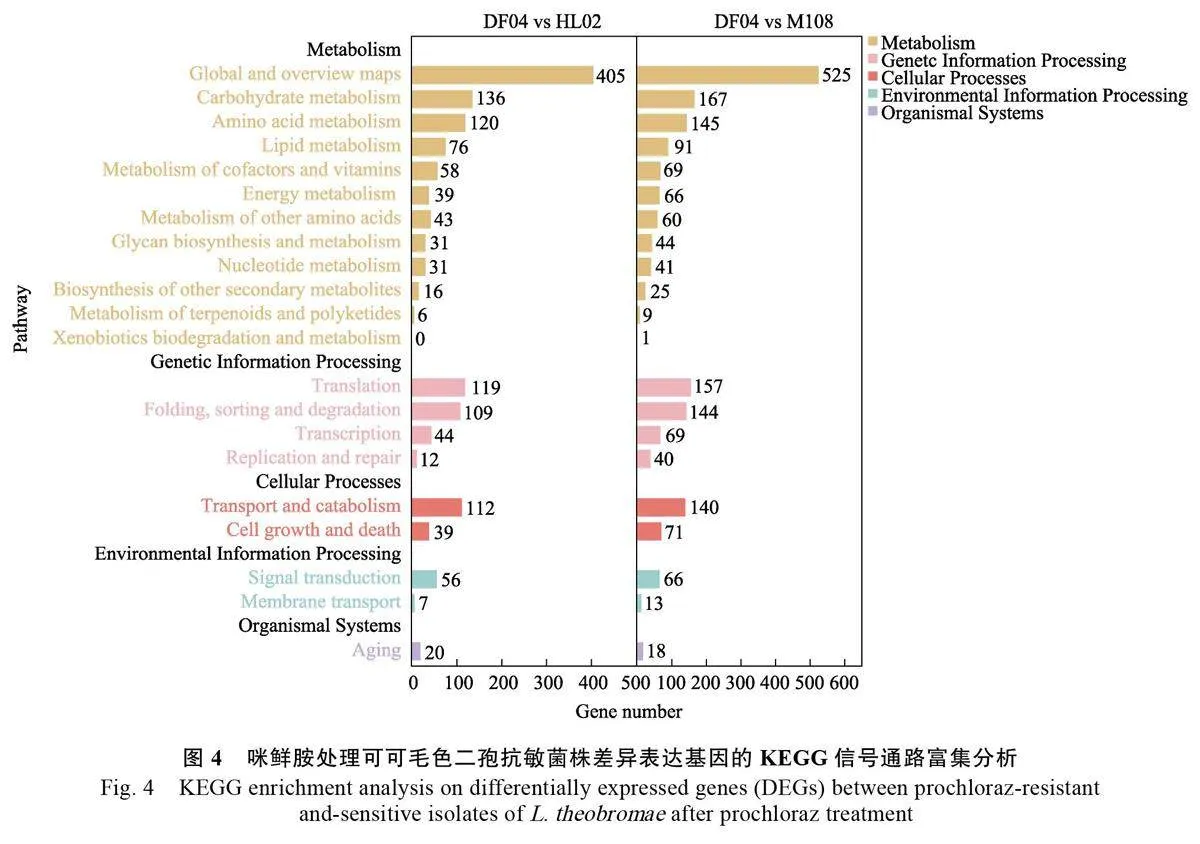
KEGG 信號通路富集分析結果顯示,咪鮮胺處理后的差異表達基因集中在代謝(metabolism)、遺傳信息處理(genetic information processing)、細胞過程(cellular processes)、環(huán)境信息處理(environmental information processing)及有機系統(tǒng)(organismal systems)通路,DF04 vsHL02 和DF04 vs M108 中DEGs 分別為1479、1961個DEGs。抗性菌株與敏感菌株之間差異表達基因顯著富集在代謝通路中,代謝通路差異基因在DF04 vs HL02 和DF04 vs M108 比對中分別占64.98%和63.39%;其中,位于前五的分別是全局和總覽圖(global and overview maps)、碳代謝(carbon metabolism)、氨基酸代謝(amino acidsmetabolism)、脂質(zhì)代謝(lipid metabolism)和輔助因子和維生素的代謝(metabolism of cofactorsand vitamins)(圖4)。
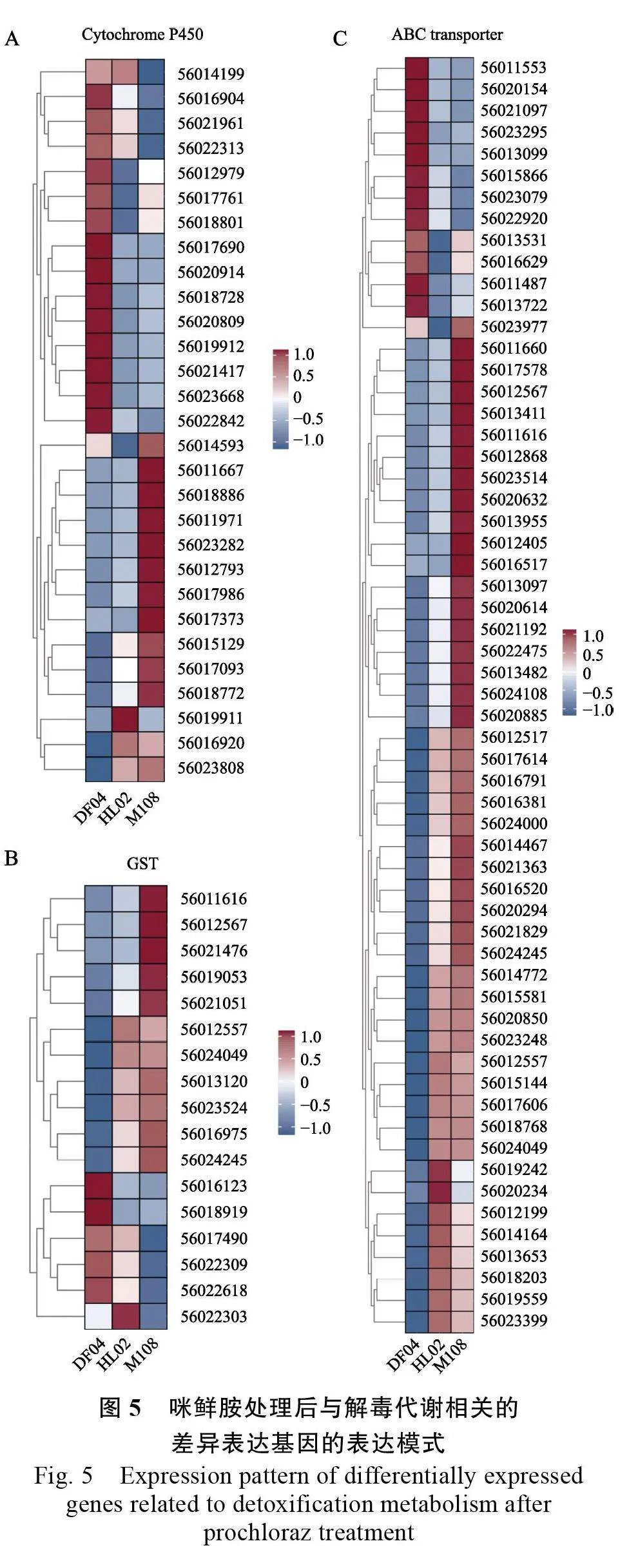
2.6 與代謝抗性相關基因的篩選及表達分析
使用細胞色素P450蛋白結構域、谷胱甘肽轉移酶蛋白結構域和ABC 轉運蛋白結構域比對到參考基因組后,結合轉錄組數(shù)據(jù)分別篩選出46 個細胞色素P450(Cytochrome P450)差異表達基因、19個谷胱甘肽轉移酶(GST)差異表達基因和84個ABC 轉運蛋白(ABC transporter)差異表達基因。其中,抗敏菌株間基因表達水平存在明顯差異,被注釋為顯著差異表達基因的有:細胞色素P450差異表達基因27 個、谷胱甘肽轉移酶差異表達基因17 個和ABC 轉運蛋白差異表達基因59 個。聚類熱圖分析表明,與敏感菌株相比,2 個抗性菌株的差異表達基因基本一致。DF04 vs HL02 和DF04 vs M108 比對中,呈現(xiàn)上調(diào)表達P450s 基因分別有14 和15 個(圖5A),呈現(xiàn)上調(diào)表達GSTs基因分別有12 和11 個(圖5B),呈現(xiàn)上調(diào)表達ABC 轉運蛋白基因分別有46 和47 個(圖5C)。
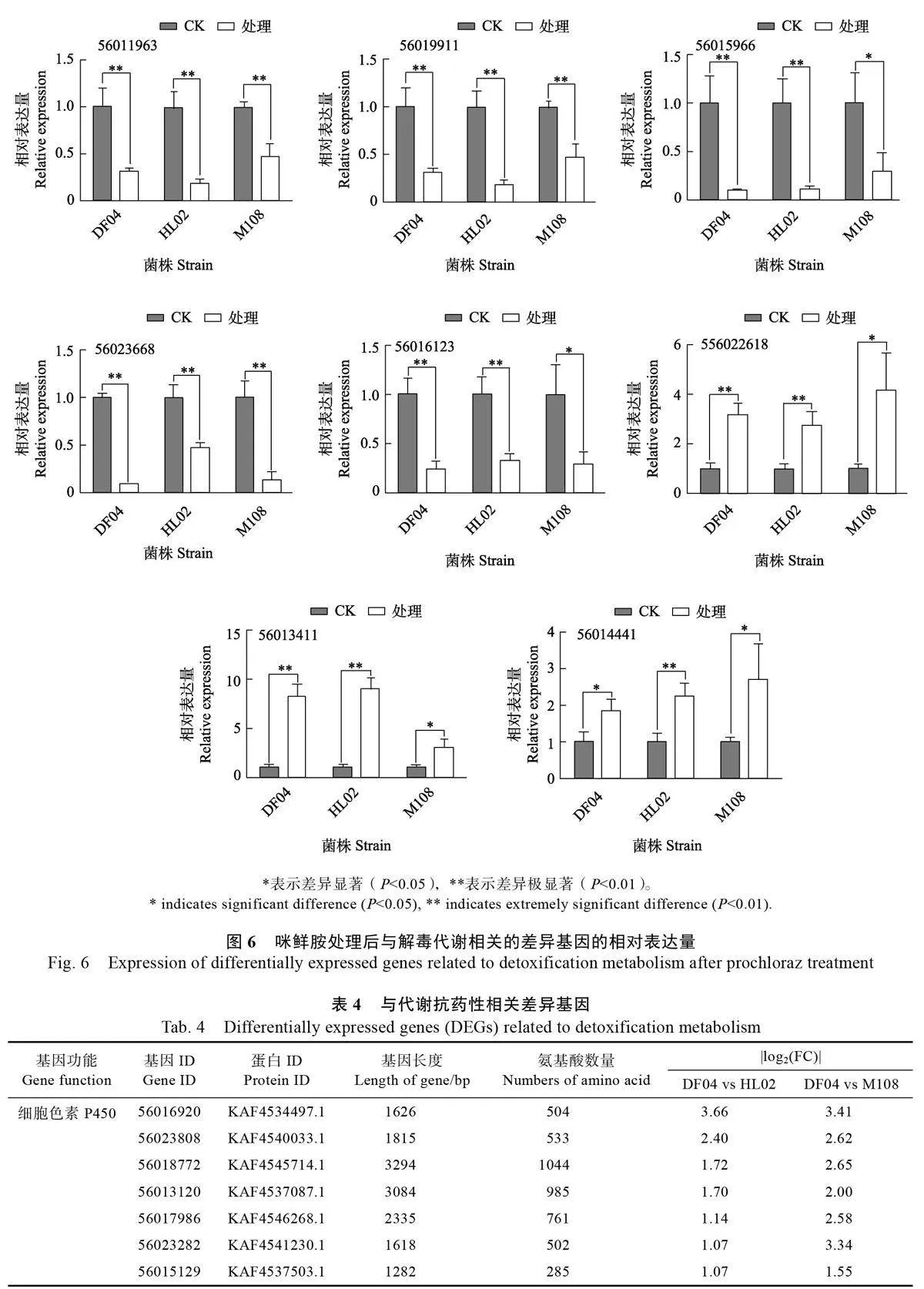
基于轉錄組數(shù)據(jù),隨機選取4 個細胞色素P450 基因(56011963、56011911、56015966、56023668)、2 個谷胱甘肽轉移酶基因(56016123、56022618)和2 個ABC 轉運蛋白基因(56013411、56014441)進行qPCR 驗證(圖6),結果表明差異表達基因變化趨勢與轉錄組數(shù)據(jù)一致,說明該轉錄組數(shù)據(jù)可靠。
代謝抗性相關的基因中,與敏感菌株相比,在2 個抗性菌株中均呈現(xiàn)顯著上調(diào)表達的基因包括:7 個P450s 基因、8 個GSTs 基因和11 個ABC轉運蛋白基因(表4)。上述基因在芒果可可毛色二孢對咪鮮胺的解毒代謝中可能發(fā)揮重要作用,抗性菌株表達水平的變化很可能與可可毛色二孢菌株抗藥性相關。
3討論
苯并咪唑類及甲氧丙烯酸酯類均為高抗性風險的殺菌劑,在國內(nèi)的海南及國外的阿曼均有發(fā)現(xiàn)。芒果蒂腐病菌可可毛色二孢對苯并咪唑類多菌靈、甲基硫菌靈和甲氧丙烯酸酯類吡唑醚菌酯、嘧菌酯等已經(jīng)產(chǎn)生嚴重抗藥性,在田間形成了具有較高抗性水平的抗藥性亞種群[14-17];此外,在木瓜上有報道蒂腐病菌可可毛色二孢對甲基硫菌靈和嘧菌酯產(chǎn)生抗藥性[18]。被劃分為中等抗性風險的甾醇脫甲基抑制劑類(DMIs)如咪鮮胺、苯醚甲環(huán)唑等,單用或者作為復配制劑廣泛應用于芒果病害防治,但目前生產(chǎn)上已經(jīng)出現(xiàn)敏感性下降或藥效降低的現(xiàn)象。咪鮮胺主要通過抑制植物病原真菌細胞膜上麥角甾醇的生物合成從而抑制菌絲生長,作為內(nèi)吸性殺菌劑同樣存在作用位點單一的缺點,長期使用同樣具有產(chǎn)生抗藥性的潛在危險。WANG 等[19-20]發(fā)現(xiàn)我國海南芒果可可毛色二孢對DMI 類殺菌劑苯醚甲環(huán)唑和咪鮮胺存在敏感性下降,并發(fā)現(xiàn)抗藥性菌株與戊唑醇存在正交互抗性。
據(jù)報道,已有許多植物病原真菌對DMI 類殺菌劑產(chǎn)生抗藥性,如桃褐腐病菌(Moniliniafructicola)[21-22],蘋果黑星病菌(Venturia inaequalis)[23]。DMI 類殺菌劑的抗藥性機制較為復雜,抗藥性菌株除了存在靶標抗性,即殺菌劑靶標14α-脫甲基酶CYP51 基因突變和過表達導致病原菌產(chǎn)生抗藥性[19, 24-26];還包括外排轉運和解毒代謝酶的變化導致代謝抗性。研究指出,DMI類藥劑的靶標基因CYP51 的點突變、ABC 轉運蛋白基因AtrD 和MFS 轉運蛋白基因Mfs1 的誘導表達,均有可能介導灰葡萄孢(Botrytis cinerea)高抗突變體對啶菌噁唑的抗藥性[ 2 7 ]。研究發(fā)現(xiàn)ABC-G 轉運蛋白基因的過量表達是導致草坪草幣斑病病菌(Sclerotinia homoeocarpa)對丙環(huán)唑產(chǎn)生抗藥性的原因[28]。ABC 轉運蛋白能夠?qū)⒂H脂性藥物和代謝產(chǎn)物排出體外,ABC 轉運蛋白基因AG1IA_06082 的過表達能夠增強立枯絲核菌(Rhizoctonia solani)對殺菌劑SYP-14288 的外排作用從而導致病原菌對殺菌劑的抗藥性[29]。WANG 等[20]的研究發(fā)現(xiàn),芒果可可毛色二孢咪鮮胺抗性菌株CYP51 基因及ABCG 基因的過表達與抗藥性的產(chǎn)生有關。這些研究說明抗藥性產(chǎn)生的機制不是單一的,可能存在多種機制。有研究表明解毒代謝相關基因的特異性表達與病原菌抗藥性有關,如細胞色素P450,這是一類由含鐵血紅素蛋白組成的超家族單加氧酶,具有廣泛的底物特異性,能夠催化多種氧化和還原反應[30]。此外,谷胱甘肽轉移酶(GST)通過催化谷胱甘肽與各種疏水性和親電性基團的結合參與解毒代謝[31]。細胞色素P450 基因AG1IA_05136 和谷胱甘肽轉移酶基因AG1IA_07383 過表達增強立枯絲核菌(Rhizoctonia solani)的解毒代謝,與病原菌對解偶聯(lián)劑SYP-14288、百菌清和苯醚甲環(huán)唑的多藥抗性有關[32]。上述研究表明,ABC 轉運蛋白基因、細胞色素P450 基因和谷胱甘肽轉移酶基因過表達均與病菌抗藥性密切相關。
大量文獻報道轉錄組測序(RNA-seq)可以篩選及挖掘抗藥性相關基因,如HELLIN等[33]通過轉錄組測序發(fā)現(xiàn)Fusarium culmorum 在經(jīng)過戊唑醇處理后,抗性菌株ABC1基因的表達量明顯高于敏感菌株的表達量。ZHANG 等[34] 通過RNA-seq 發(fā)現(xiàn)ABC 轉運蛋白基因和MFS 轉運蛋白基因參與了Penicillium italicum 對咪鮮胺的抗藥性。目前尚未有代謝酶參與可可毛色二孢抗藥性的研究報道。因此,本研究采用RNA-seq 技術分析對芒果可可毛色二孢咪鮮胺的抗敏菌株進行轉錄組測序,對咪鮮胺處理后抗敏菌株之間差異表達基因的分析發(fā)現(xiàn),2個抗性菌株的差異表達基因主要集中在代謝通路中。根據(jù)GO 分析和KEGG 信號通路富集分析,將篩選出的與解毒代謝相關的細胞色素P450 基因、谷胱甘肽轉移酶基因和ABC 轉運蛋白基因進行進一步比較,發(fā)現(xiàn)2個抗性菌株的表達模式基本一致。本研究從轉錄組數(shù)據(jù)中篩選出的7 個細胞色素P450 基因、8個谷胱甘肽轉移酶基因和11個ABC轉運蛋白基因在咪鮮胺抗敏菌株間顯著上調(diào)表達。其中,ABC轉運蛋白基因56017578 和56024245 在GO功能注釋上被認為是編碼多藥耐藥轉運蛋白的基因,在灰葡萄孢上發(fā)現(xiàn)ABC轉運蛋白基因BcatrB 敲除突變體對植物抗毒素白藜蘆醇和殺菌劑拌種咯的敏感性均增加[35],煙曲霉中ABC轉運蛋白基因atrF 的過表達被證明與伊曲康唑耐藥性相關[36]。已有研究報道煙曲霉和白色念珠菌ABC轉運蛋白家族的多藥耐藥轉運蛋白基因過表達可能導致真菌的多藥抗性[37]。其它差異表達基因并沒有相關文獻報道與殺菌劑抗藥性相關,推測這些基因很可能與芒果可可毛色二孢菌株對咪鮮胺的抗藥性有關,但這些基因是否真正參與病原菌抗藥性還需要采用基因過表達、基因敲除和RNAi 等方法進一步驗證。本研究首次對可可毛色二孢進行代謝抗藥性的研究,并發(fā)現(xiàn)部分細胞色素P450、谷胱甘肽轉移酶和ABC 轉運蛋白基因在咪鮮胺處理芒果可可毛色二孢后顯著上調(diào)表達,表明芒果可可毛色二孢對咪鮮胺的抗藥性可能與多種代謝途徑相關。
