產蛋間隔前期鴿卵泡轉錄組比較分析揭示卵泡發育相關基因
2024-09-30 00:00:00陳靜吳薛蓓苗冬枝張弛郭振玉王瑩
畜牧獸醫學報 2024年8期



摘 要: 旨在對產蛋間隔前期母鴿卵泡顆粒細胞(GC)層進行高通量測序,篩選出與鴿卵泡發育和選擇相關的關鍵基因。本研究選擇了80對12月齡產蛋規律、體重相近的白羽王鴿為研究對象,在產蛋間隔第1天(LI1)和第3天(LI3)分別選擇3只母鴿,采集F1和SF1卵泡GC層,并利用RNA-seq進行轉錄組分析,篩選L1F1/L1SF1、L3F1/L3SF1、L1F1/L3F1和L1SF1/L3SF1組,4組間差異表達基因(DEGs),對DEGs進行GO功能注釋和KEGG通路富集分析,通過STRING數據庫進行蛋白質互作(PPI)網絡分析并使用Cytoscape軟件進行可視化處理篩選關鍵基因,各組隨機選擇5個DEGs進行RT-qPCR驗證轉錄組的可靠性。在4組比對中分別獲得77、2 736、5 698和3 864個DEGs。GO分析顯示,4組DEGs都主要富集的GO條目為細胞過程、細胞解剖實體和連接。KEGG通路富集分析顯示,L1F1/L1SF1組和L3F1/L3SF1組DEGs均顯著富集在類固醇激素生物合成和卵巢類固醇合成等信號通路,L1F1/L3F1組和L1SF1/L3SF1組DEGs均顯著富集在糖胺聚糖降解、突觸囊泡周期和氧化磷酸化等信號通路。進一步地,進行PPI網絡分析并篩選與卵泡發育和選擇密切相關的DEGs。各組隨機選擇5個DEGs進行RT-qPCR驗證,表達趨勢與測序結果一致。本研究利用RNA-seq技術篩選到與鴿卵泡發育和選擇相關的關鍵基因——EEF2、DDX5、PLK2、IGF2R、LHCGR、HSD3B1、CYP19A1和StAR,為進一步探究鴿卵泡發育和選擇的分子調控機制提供了理論依據。
關鍵詞: 鴿;產蛋間隔;卵泡發育;轉錄組
中圖分類號:S836.2
文獻標志碼:A
文章編號:0366-6964(2024)08-3503-13
收稿日期:2024-02-01
基金項目:國家自然科學基金(32372875);江蘇高校優勢學科建設工程四期項目(PAPD)
作者簡介:陳 靜(2001-),女,陜西咸陽人,碩士生,主要從事家禽繁殖研究,E-mail:cj82955@163.com
通信作者:王 瑩,主要從事家禽繁殖研究工作,E-mail:dkwangying@yzu.edu.cn
Comparative Analysis of Transcriptome of Pigeon Follicles at Early Stage of Laying Interval
Reveals Genes Related to Follicular Development
CHEN" Jing, WU" Xuebei, MIAO" Dongzhi, ZHANG" Chi, GUO" Zhenyu, WANG" Ying*
(College of Animal Science and Technology, Yangzhou University, Yangzhou 225009," China)
Abstract:" High throughput sequencing was performed on granulosa cell(GC)layer at early stage of laying interval(LI)to screen key genes involved in pigeon follicle development and selection. Eighty pairs of 12-month-old White King pigeons, with similar laying patterns and body weights were selected in this study. Three female pigeons were selected at first day (LI1) and third day (LI3) of LI, respectively, the GC layer of F1 and SF1 follicles were collected, followed by transcriptome analysis using RNA-seq. Differentially expressed genes (DEGs) among 4 groups: L1F1/L1SF1 group, L3F1/L3SF1 group, L1F1/L3F1 group, and L1SF1/L3SF1 group were analyzed. Gene Ontology (GO) functional annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of DEGs were performed, protein-protein interaction (PPI) network analysis using the STRING database and visualization using Cytoscape software were conducted to selected key genes. Five DEGs from each group were randomly selected for RT-qPCR to verify the reliability of transcriptome. The 77, 2 736, 5 698 and 3 864 DEGs were obtained in the 4 groups, respectively. GO analysis results showed that DEGs in all groups were mainly enriched in cellular processes, cellular anatomical entity and binding. KEGG analysis result showed that DEGs in L1F1/L1SF1 and L3F1/L3SF1 groups were enriched in the steroid hormone biosynthesis and ovarian steroidogenesis signaling pathways. DEGs in L1F1/L3F1 and L1SF1/L3SF1 groups were enriched in the glycosaminoglycan degradation, synaptic vesicle cycle and oxidative phosphorylation signaling pathways. In addition, DEGs closely associated with follicle development and selection were screened by PPI.Five DEGs were randomly selected from each group for RT-qPCR verification, and the expression trends of these DEGs were consistent with the sequencing results. RNA-seq technology was applied to identify key genes associated with pigeon follicle development and selection, which including EEF2, DDX5, PLK2, IGF2R, LHCGR, HSD3B1, CYP19A1 and StAR. These results will provide the theoretical basis for further exploring the molecular mechanisms of follicle development and selection in pigeon.
Key words: pigeon; laying interval; follicular development; transcriptome
*Corresponding author:WANG Ying, E-mail:dkwangying@yzu.edu.cn
鴿已成為我國繼雞、鴨、鵝之后的第四大家禽[1],鴿蛋是富含蛋白質、脂質、氨基酸和礦物質的健康動物蛋白[2],據報道,與其他禽蛋相比,鴿蛋的脂肪含量低、鈣含量較高[3],是理想的滋補佳品。隨著人們生活水平的提高,對鴿蛋的需求也越來越大,而配對種鴿在產蛋周期內只產2枚蛋[4],且產蛋間隔期較長,鴿的這些繁殖特性影響了其產蛋性能。家禽產蛋性能與卵泡發育和排卵密切相關,卵泡發育涉及卵泡募集、選擇和排卵,是一個復雜的生物過程[5]。按照直徑大小,家禽卵泡可分為等級卵泡和等級前卵泡,等級卵泡由大到小命名為F1~F6,等級前卵泡又分為小白卵泡、大白卵泡、小黃卵泡、大黃卵泡[6]。本課題組前期研究發現,與其他家禽不同,鴿卵巢僅有兩個排卵前卵泡F1和F2,等級前卵泡——小卵泡(SF)在卵巢上成對存在,按照發育等級劃分為SF1、SF2、SF3、SF4[7]。卵泡主要由膜細胞層、顆粒細胞(granular cell, GC)層和卵母細胞組成[8],GC層分泌雌二醇(estradiol, E2)和孕酮(progesterone, P4)能引發卵母細胞的成熟和排卵,因此,GC的增殖和分化與卵泡發育密切相關[9-10]。
隨著轉錄組測序技術的成熟,對家禽卵巢、卵泡基因表達譜的研究越來越多,張俊珍等[11]對不同生理時期邊雞卵巢進行轉錄組分析,篩選出與邊雞卵泡發育相關的基因,為研究其就巢性提供了依據。Yang等[12]對鴨GC進行轉錄組分析,發現降低類固醇激素合成相關基因的表達可抑制GC合成E2。Sun等[13]對雞白卵泡、小黃卵泡和大黃卵泡進行轉錄組分析,篩選出與其產蛋性能相關的候選基因。本課題組前期初步探究了產蛋間隔期鴿卵泡的發育圖譜及生殖激素的調控機制,并發現產蛋間隔前期(LI1和LI3期)SF1卵泡還未選擇為F2卵泡[14],本試驗通過對白羽王鴿F1和SF1的GC層進行轉錄組測序,篩選獲得與卵泡發育和選擇相關的關鍵基因,為促進等級卵泡選擇、縮短卵泡成熟間隔,進而提高鴿產蛋性能提供理論依據。
1 材料與方法
1.1 試驗動物
選擇80對12月齡產蛋規律、體重相近的白羽王鴿為研究對象,試驗期60 d,試驗期間按照常規飼養管理制度進行,觀察并記錄產蛋間隔。公母配對的種鴿產兩枚蛋后,取出鴿蛋,進入下一輪產蛋,鴿產蛋間隔為8~10 d。在產蛋間隔第1天(LI1)和第3天(LI3)分別選擇3只母鴿,利用RNA-seq進行轉錄組分析。
1.2 樣本采集
在LI1和LI3期采集鴿F1和SF1卵泡,放置于預冷PBS中,沖洗干凈,分離卵泡GC層,分別標記為L1F1-1、L1F1-2和L1F1-3;L3F1-1、L3F1-2和L3F1-3;L1SF1-1、L1SF1-2和L1SF1-3;L3SF1-1、L3SF1-2和L3SF1-3。由于SF1的GC層樣本量少,將2個SF1的GC層合并,迅速放于液氮,然后置于-80 ℃冰箱中保存備用。
1.3 轉錄組測序分析
使用Trizol試劑提取GC層總RNA,RNA的質量和濃度采用2100 Bioanalyzer檢測,選擇符合標準的RNA(A260 nm/A280 nm≥1.8,A260 nm/A230 nm≥2.0,RNA完整性數gt;7.0)用于合成cDNA并構建文庫。利用Illumina Hiseq 2000測序平臺進行測序,對原始下機數據進行過濾。利用HISAT軟件將樣本Clean Reads與鴿的參考基因組(GCA_000337935.1)進行比對,利用FPKM算法計算基因表達量,利用DESeq2軟件對各個樣本基因的counts數目進行標準化處理。以log2FCgt;2和q-valuelt;0.05篩選出L1F1/D1SF1、L3F1/L3SF1、L1F1/L3F1和L1SF1/L3SF1組的DEGs,并繪制火山圖。
1.4 GO、KEGG和PPI網絡分析
利用GO數據庫和KEGG數據庫對L1F1/L1SF1、L3F1/L3SF1、L1F1/L3F1和L1SF1/L3SF1組DEGs進行功能注釋,利用STRING數據庫預測蛋白質互作關系,使用Cytoscape軟件構建可視化PPI網絡。
1.5 實時熒光定量PCR檢測
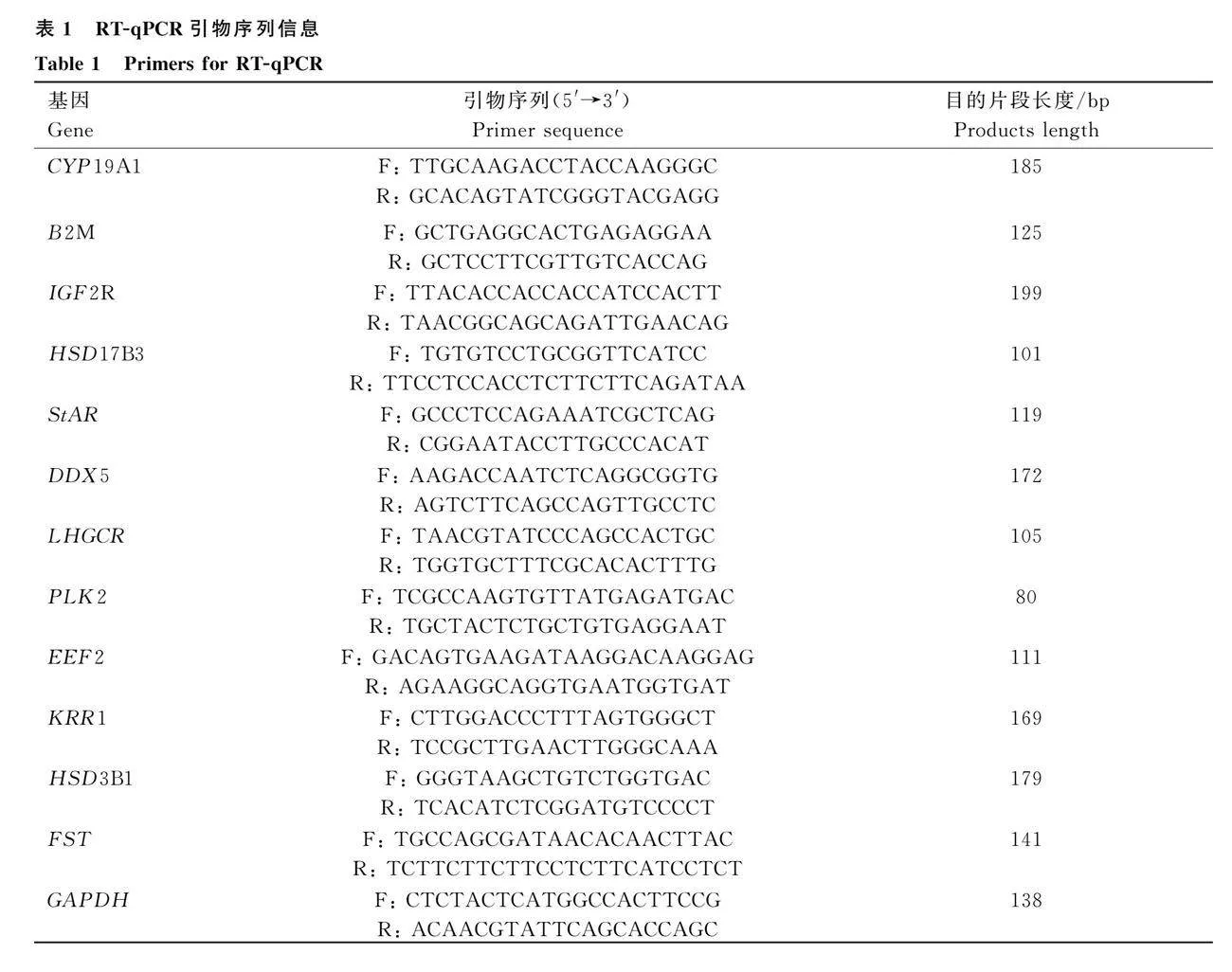
各組隨機選擇5個DEGs進行RT-qPCR驗證,根據NCBI中鴿(Columba livia)參考序列,利用Primer 6.0軟件設計引物,引物信息見表1。使用SuperReal PreMix Plus(SYBR Green)試劑盒進行熒光定量驗證,反應體系20 μL:2×Super Real Premix 10.0 μL,50×ROX Reference Dye 0.4 μL,上、下游引物(10 μmol·L-1)各0.6 μL,cDNA(200 ng·μL-1)1.0 μL和ddH2 O 7.4 μL。反應程序:95 ℃預變性15 min;95 ℃變性10 s,60 ℃退火/延伸32 s,共40個循環。以鴿GAPDH為內參基因,每個樣品重復3次。
1.6 統計分析
采用SPSS18.0軟件對試驗數據進行ANOVA分析,采用2-ΔΔCt法計算基因相對表達量 并使用GraphPadPrism8軟件進行繪圖。
2 結 果
2.1 轉錄組測序數據分析
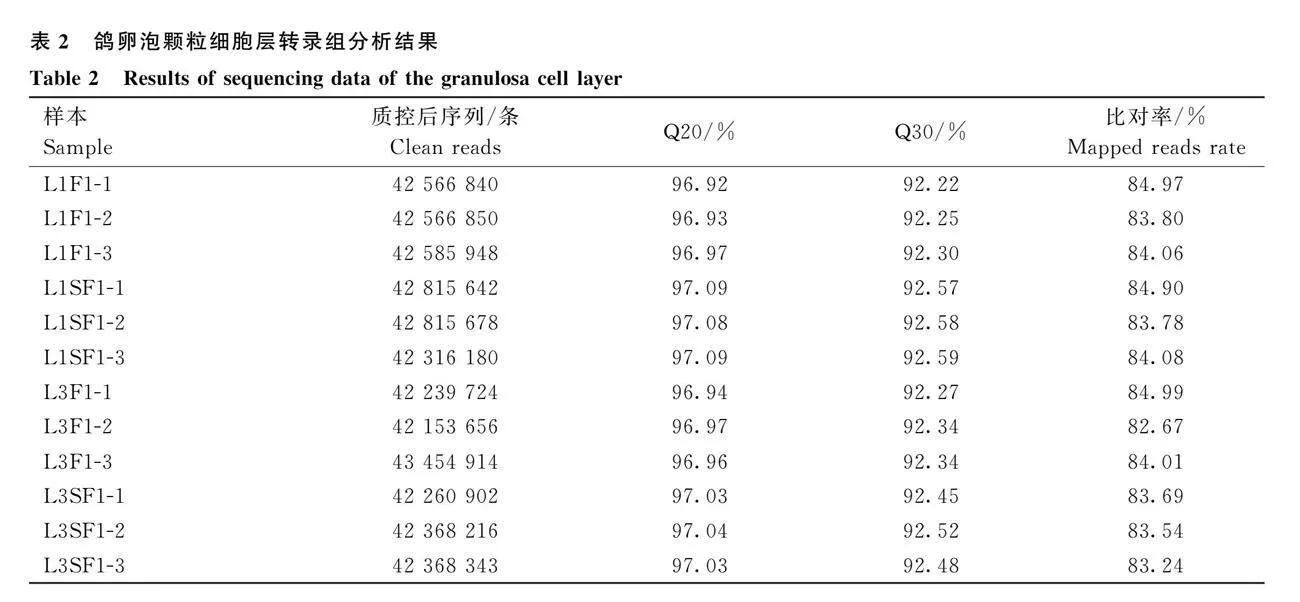
LI1和LI3期鴿F1和SF1卵泡GC層cDNA共構建了12個文庫,質控后共獲得42 153 656~43 454 914條Clean reads,所有樣本的Q20大于96.92%,Q30大于92.22%。將過濾得到Clean reads與鴿參考基因組進行對比,比對率為82.67%~84.99%,說明測序數據可信度高,可用于生物信息學分析(表2)。
2.2 DEGs表達分析
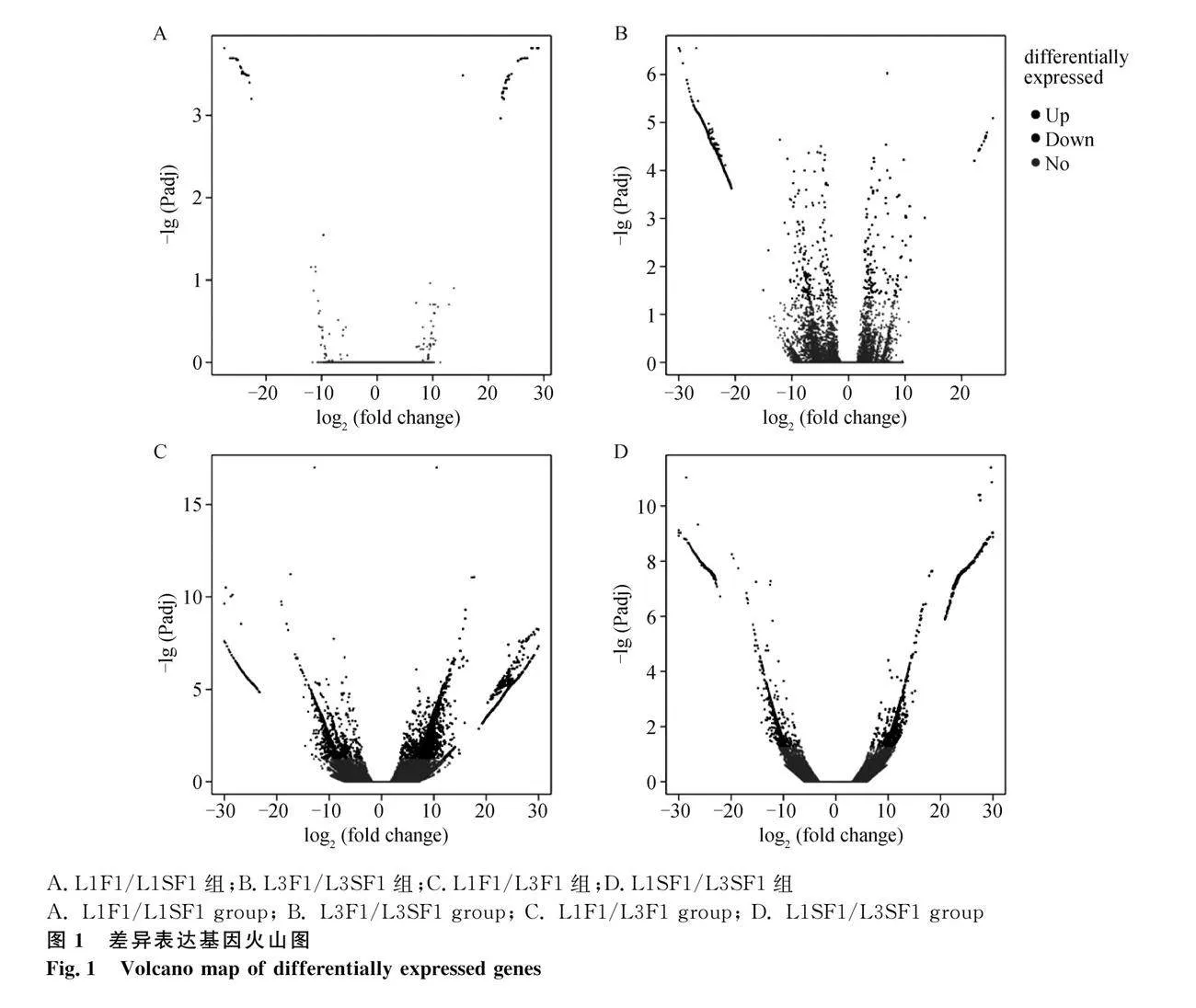
在L1F1/L1SF1組,共篩選出了77個DEGs,其中34個基因上調和43個基因下調(圖1A)。在L3F1/L3SF1組共篩選出了2 736個DEGs,其中131個基因上調,2 605個基因下調(圖1B)。在L1F1/L3F1組共篩選出了5 698個DEGs,其中4 161個基因上調,1 537個基因下調(圖1C)。在L1SF1/L3SF1組共篩選出了3 864個DEGs,其中1 987個基因上調,1 877個基因下調(圖1D)。
2.3 GO和KEGG富集分析
按照生物過程(biological process, BP)、分子功能(molecular function, MF)和細胞組分(Cellular component, CC)對L1F1/L1SF1組、L3F1/L3SF1組、L1F1/L3F1組和L1SF1/L3SF1組的DEGs進行GO注釋,并對其進行和KEGG富集分析。
2.3.1 L1F1/L1SF1組
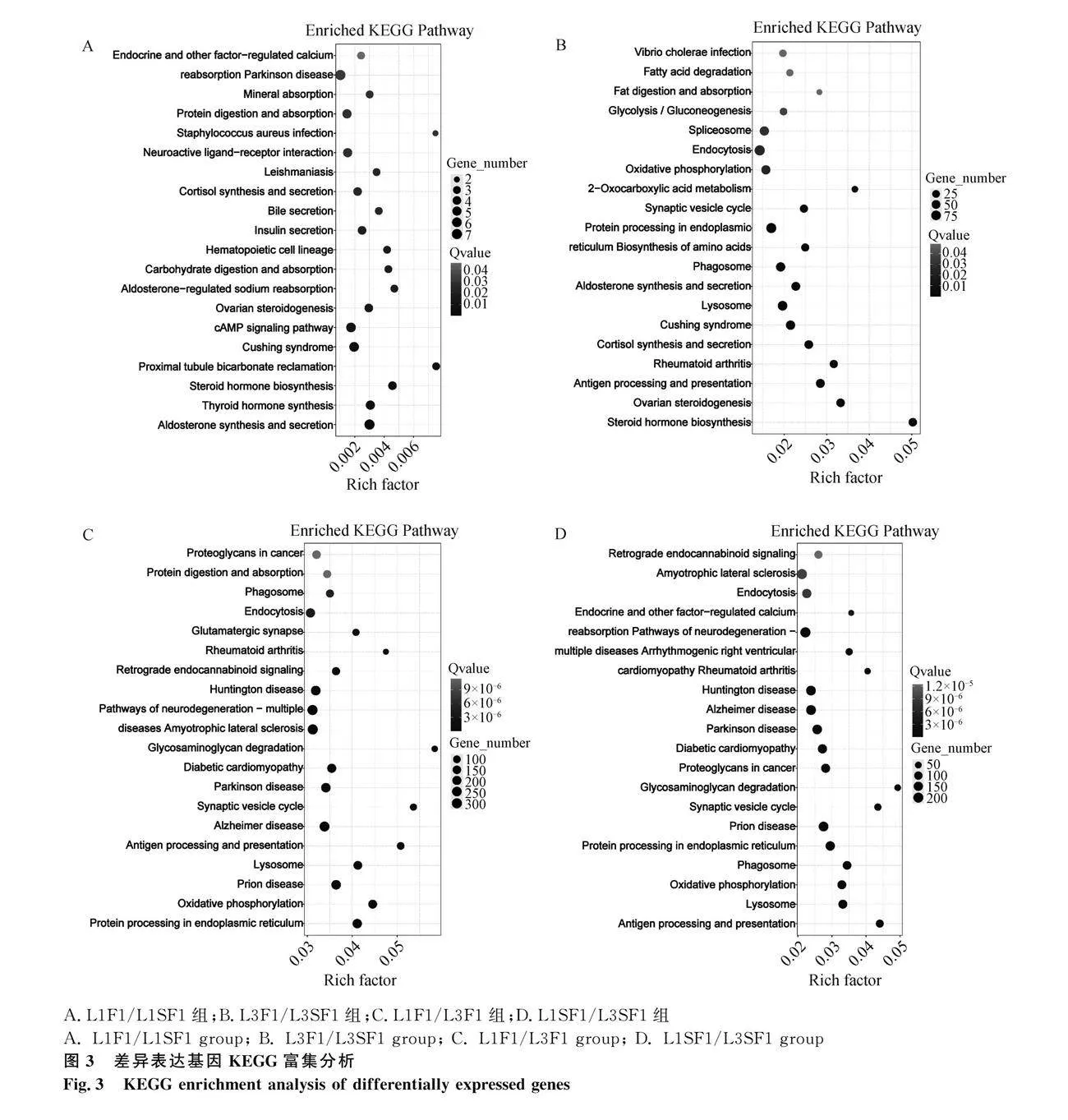
L1F1/L1SF1組DEGs進行GO功能富集和KEGG信號通路分析,獲得25個顯著GO條目(圖2A):14個生物過程、2個細胞組分、9個分子功能和顯著富集的前20個KEGG通路(圖3A)。顯著GO條目主要有生物過程,如細胞過程、代謝過程;細胞組分,如細胞解剖實體、含蛋白質復合物;分子功能,如連接、催化活性等。與卵泡發育相關信號通路有類固醇激素生物合成、甲狀腺激素合成、卵巢類固醇合成、醛固酮-調節鈉的重吸收等。
2.3.2 L3F1/L3SF1組
L3F1/L3SF1組的DEGs進行GO功能富集和KEGG信號通路分析,獲得41個顯著GO條目(圖2B):23個生物過程、2個細胞組分、16個分子功能和顯著富集的前20個KEGG通路(圖3B)。顯著GO條目主要有生物過程,如細胞過程、代謝過程、生物調節;細胞組分,如細胞解剖實體、含蛋白質復合物;分子功能,如連接、催化活性、轉運體活性等。與卵泡發育相關信號通路有類固醇激素生物合成、卵巢類固醇合成、醛固酮的合成和分泌和突觸囊泡周期。
2.3.3 L1F1/L3F1組
L1F1/L3F1組的DEGs進行GO和KEGG富集分析,獲得41個顯著GO條目(圖2C):23個生物過程、2個細胞組分、16個分子功能和顯著富集的前20個KEGG通路(圖3C)。顯著GO條目主要有生物過程,如細胞工程、代謝過程、生物調節、生物過程調節、定位、刺激反應;細胞組分,如細胞解剖實體、含蛋白質復合物;分子功能,如連接、催化活性、轉運體活性、分子功能調節器、ATP-依賴活性等。與卵泡發育相關信號通路有糖胺聚糖降解、突觸囊泡周期和氧化磷酸化。
2.3.4 L1SF1/L3SF1組
L1SF1/L3SF1組的DEGs進行GO和KEGG富集分析,獲得42個顯著GO條目(圖2D):24個生物過程、2個細胞組分、16個分子功能和顯著富集的前20個KEGG通路(圖3D)。DEGs與生物過程,如細胞工程、代謝過程、生物調節、多細胞生物過程、生物過程的負調控;細胞組分,如細胞解剖實體、含蛋白質復合物;分子功能,如連接、催化活性等GO條目。與卵泡發育相關信號通路有糖胺聚糖降解、突觸囊泡周期和氧化磷酸化等信號通路。
2.4 PPI網絡構建
2.4.1 L1F1/L1SF1組
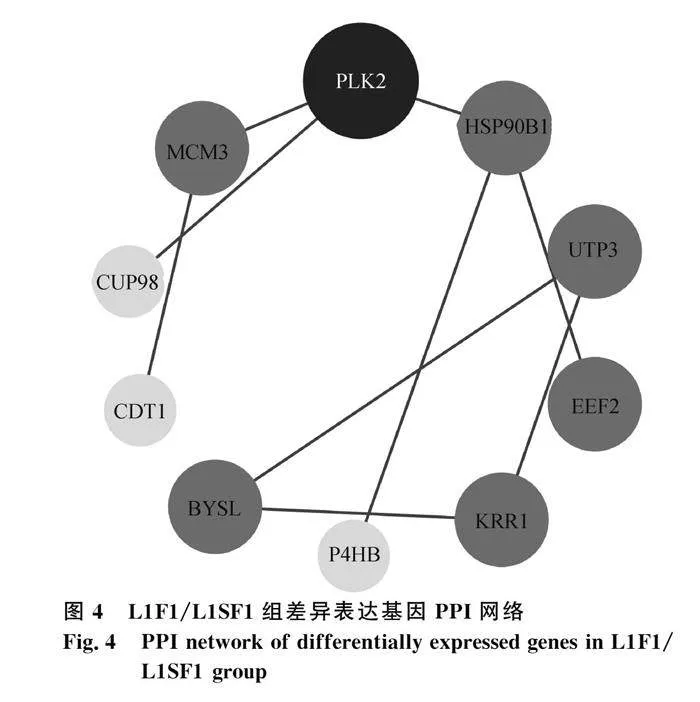
將L1F1/L1SF1組DEGs導入STRING數據庫進行PPI網絡分析,去除孤立蛋白后將數據導入Cytoscape軟件進行可視化處理(圖4)。網絡中的核心基因有PLK2、MCM3、HSP90B1、UTP3、EEF2和KRR1,結合KEGG通路和參考文獻,發現PLK2和EEF2基因在卵泡發育中起關鍵作用。
2.4.2 L3F1/L3SF1組
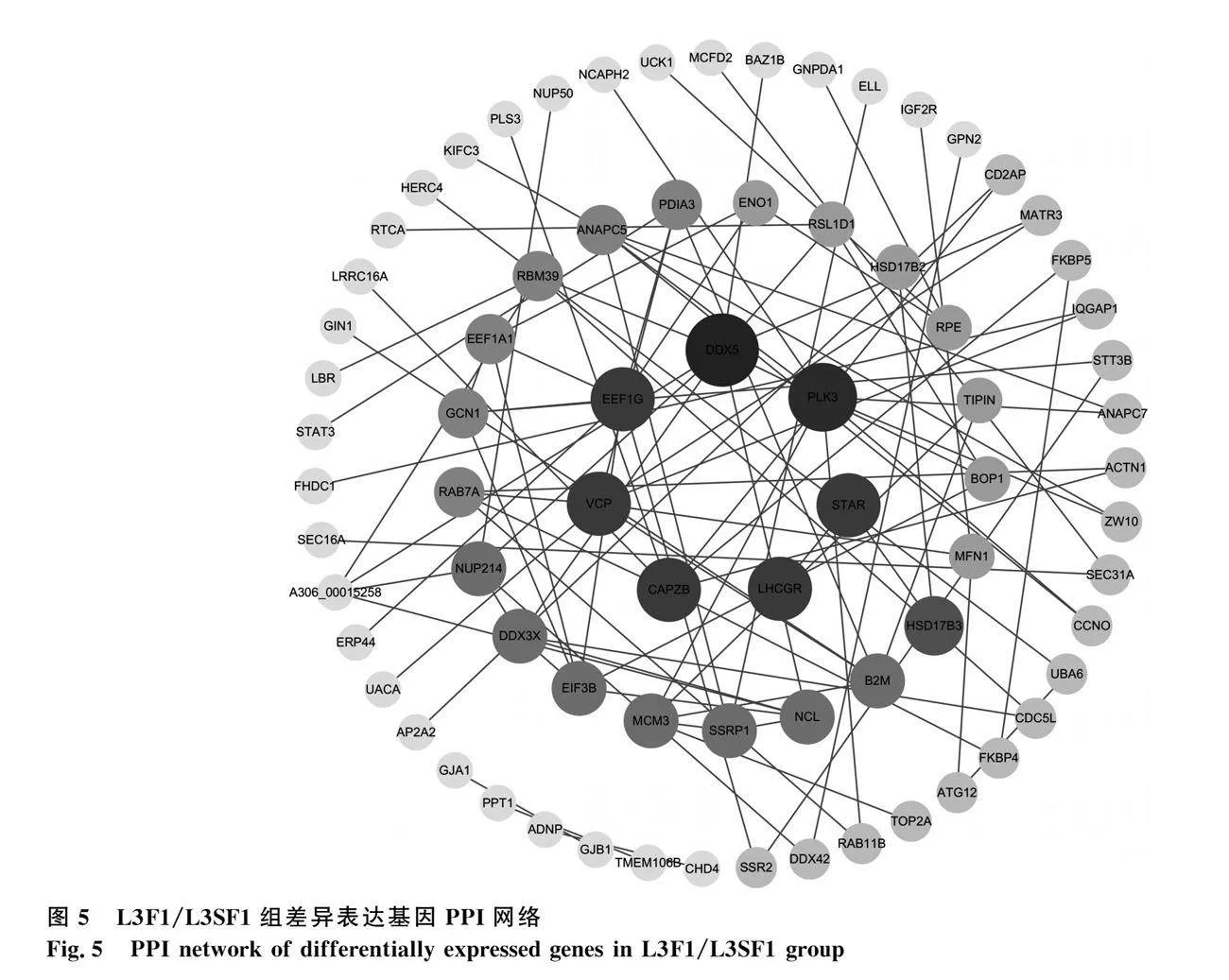
將L3F1/L3SF1組DEGs導入STRING數據庫進行PPI網絡分析,去除孤立蛋白后將數據導入Cytoscape軟件進行可視化處理,包括75個基因137條互作關系(圖5)。網絡中節點度較高的基因有DDX5、PLK3、StAR、LHCGR、CAPZB、VCP、EEF1G和HSD17B3,結合KEGG通路和參考文獻,發現DDX5、StAR、LHCGR基因在卵泡發育中起關鍵作用。
2.4.3 L1F1/L3F1組
將L1F1/L3F1組DEGs導入STRING數據庫進行PPI網絡分析,去除孤立蛋白后將數據導入Cytoscape軟件進行可視化處理,包括123個基因212條互作關系(圖6)。網絡中的核心基因有HSP90B1、HSD17B3、ATF2、IGF2R、CDC5L、DDX5、VCP、RAD23B和CLTCL1,結合KEGG通路和參考文獻,發現IGF2R和DDX5與卵泡發育相關。
2.4.4 L1SF1/L3SF1組
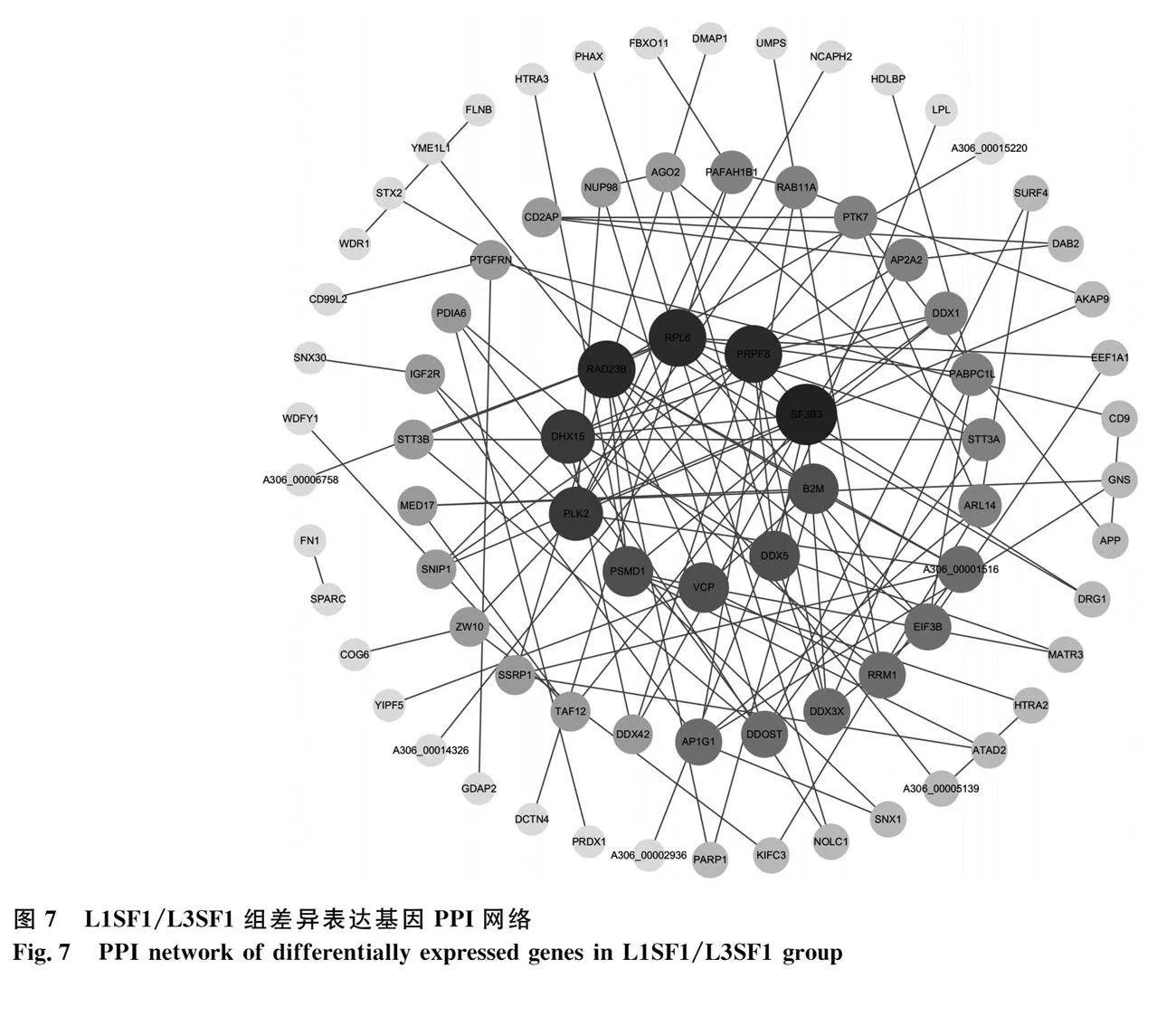
將L1SF1/L3SF1組DEGs導入STRING數據庫進行PPI網絡分析,去除孤立蛋白后將數據導入Cytoscape軟件進行可視化處理,包括79個基因115條互作關系(圖7)。網絡中的核心基因有RPL6、PRPF8、SF3B3、B2M、DDX5、VCP、PSMD1、PLK2、DHX15和RAD23B,結合KEGG通路和參考文獻,發現DDX5和PLK2與卵泡發育相關。
2.5 RT-qPCR驗證
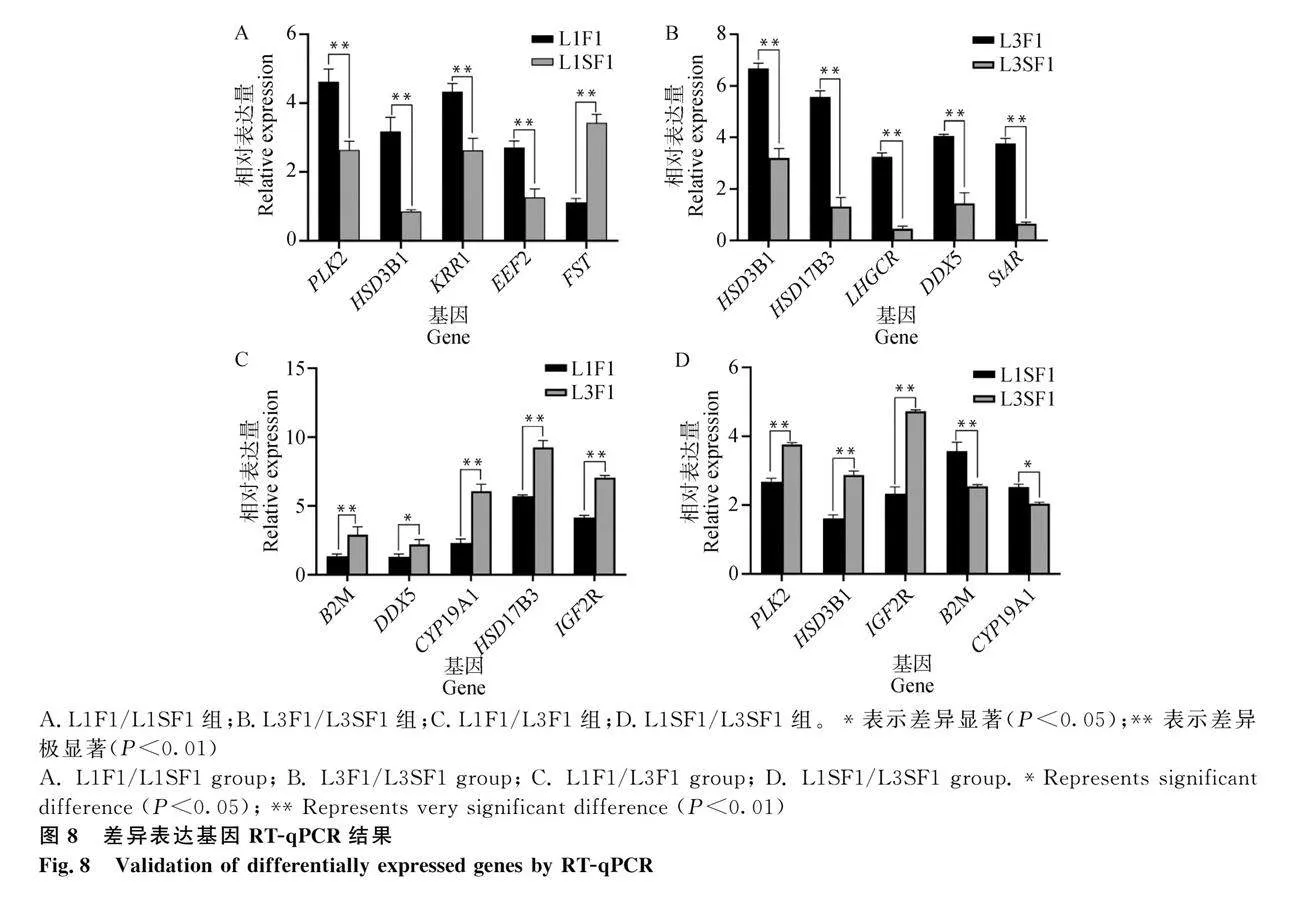
各組分別選擇5個DEGs進行RT-qPCR驗證(圖8),L1F1/L1SF1組(PLK2、HSD3B1、KRR1、EEF2和FST),L3F1/L3SF1組(HSD3B1、HSD17B3、LHGCR、DDX5和StAR),L1F1/L3F1組(B2M、DDX5、CYP19A1、HSD17B3和IGF2R),L1SF1/L3SF1組(PLK2、HSD3B1、IGF2R、B2M和CYP19A1),這些DEGs的表達趨勢與轉錄組測序結果一致,證明了本次測序結果的可靠性(圖9)。
3 討 論
產蛋性能是家禽生產的重要經濟性狀之一,卵泡發育在很大程度上決定了家禽的產蛋性能[15]。本研究利用轉錄組篩選了產蛋間隔前期(LI1和LI3期)白羽王鴿F1和SF1卵泡GC層中與卵泡發育和選擇相關的關鍵基因,為探究鴿卵泡發育分子機制提供理論基礎。為探索在同一時期不等級的卵泡以及同一等級的卵泡在不同時期的發育變化情況,本試驗進行了L1F1/L1SF1、L3F1/L3SF1、L1F1/L3F1和L1SF1/L3SF1組4組分析。對4組的DEGs進行GO和KEGG富集分析,進一步將DEGs進行PPI網絡分析,篩選獲得真核翻譯延伸因子2(EEF2)、DEAD-box RNA解旋酶(DDX5)、Polo樣激酶2(PLK2)、胰島素樣生長因子受體2(IGF2R)和黃體生成素/絨毛膜促性腺激素受體(LHCGR)和類固醇生成急性調節蛋白(StAR)基因在卵泡發育過程中起關鍵作用,此外,3β-羥基類固醇脫氫酶(HSD3B1)和P450芳香化酶基因(CYP19A1)在各組中也差異表達,并參與類固醇激素合成調控,在卵泡選擇中發揮重要作用。
PLK2是polo樣激酶家族成員之一[16],研究發現PLK2具有抗增殖作用,高水平PLK2引起GC細胞周期停滯[17-18]。本試驗發現,與SF1卵泡GC層相比,F1的GC層中PLK2基因僅在LI1期差異表達,隨著F1卵泡的生長發育PLK2的表達量逐漸降低,抗GC增殖作用減弱,這有利于F1卵泡的發育。而在SF1的GC層隨著卵泡發育PLK2基因表達量不斷升高,抑制了SF1卵泡發育。eEF2是真核細胞肽鏈延伸的重要因子[19]。據報道,eEF2是細胞增殖的啟動子[20],增加細胞的增殖和遷移,減少細胞凋亡[21]。eEF2參與調控雞卵泡的生長發育,并促進卵泡GC中E2的合成[22-23]。DDX5是具有多種生物學功能的核蛋白,對多種轉錄因子有激活作用,同時能夠促進細胞增殖[24-25]。DDX5可作為E2受體α的轉錄共激活因子[26],促進卵泡發育。本試驗發現,eEF2和DDX5的表達趨勢相同,在F1的GC層表達量高于SF1,兩者隨著卵泡的發育而不斷升高,促進卵泡GC增殖以及E2的合成,進而促進卵泡的發育和選擇。
IGF2R是胰島素樣生長因子2(IGF2)的清除受體[27],通過清除游離的IGF2參與器官和組織的發育[28]。IGF2是局部調節卵泡發育的卵巢內生長因子[29],Tkachenko等[30]研究發現IGF2響應類固醇激素調節并促進GC增殖和卵泡發育。本試驗中,LI3期IGF2R在卵泡中表達水平均高于LI1期,表明IGF2促進卵泡在產蛋間隔期不斷發育。類固醇激素參與卵泡的發育和成熟[31],StAR將膽固醇從線粒體外膜轉運至內膜,然后膽固醇被類固醇生成酶——HSD3B1和CYP19A1等轉化為類固醇激素[32]。其中StAR和HSD3B1在P4合成中起關鍵作用,CYP19A1是雄激素轉化為E2的關鍵酶,可促進GCs增殖[33-34]。本試驗中StAR、HSD3B1和CYP19A1基因的表達量均隨著各級卵泡發育而升高,其通過促進卵泡GC合成E2和P4進而促進卵泡的發育,在產蛋間隔前期卵泡發育過程中起重要作用,這與Sechman等[35]在雞上的研究結果一致。LHCGR是一種G蛋白偶聯受體,與促黃體生成素和人絨毛膜促性腺激素基因結合,激活下游信號通路,刺激P4的合成和促進排卵[36]。LI3期F1卵泡中LHCGR表達水平顯著高于SF1卵泡,LI3期F1卵泡不斷發育到排卵前卵泡,LHCGR高表達促進其發育,保證等級卵泡的成熟和排卵。
通過轉錄組測序篩選獲得了與細胞增殖和類固醇激素合成相關的基因,其在鴿卵泡發育和選擇中發揮重要作用,為研究鴿不同等級卵泡之間的基因表達譜差異及鴿卵泡發育和選擇的分子調控機制提供了理論參考。
4 結 論
本試驗通過構建產蛋間隔前期(LI1和LI3)鴿F1和SF1卵泡GC層基因表達譜,分別在4個組中獲得了77、2 736、5 698和3 864個DEGs。進一步通過GO、KEGG和PPI網絡分析進行各組間比較,篩選獲得EEF2、DDX5、PLK2、IGF2R、LHCGR、HSD3B1、CYP19A1和StAR等在鴿卵泡發育和選擇中起關鍵作用的基因。
參考文獻(References):
[1] 王明禮,王 猛,李延森,等.不同粗蛋白水平人工鴿乳對乳鴿生長性能、血清抗氧化水平及腸道發育的影響[J].畜牧獸醫學報,2023,54(4):1545-1554.
WANG M L,WANG M,LI Y S,et al.Effect of different crude protein levels of artificial crop milk on growth performance,serum antioxidant level and intestinal development of squabs[J].Acta Veterinaria et Zootechnica Sinica,2023,54(4):1545-1554.(in Chinese)
[2] SUN C J,LIU J N,YANG N,et al.Egg quality and egg albumen property of domestic chicken,duck,goose,turkey,quail,and pigeon[J].Poult Sci,2019,98(10):4516-4521.
[3] 王云浩,鄭玉才,李志雄,等.不同種類禽蛋的蛋品質和蛋內營養成分的比較分析[J].畜牧與獸醫,2022,54(5):40-44.
WANG Y H,ZHENG Y C,LI Z X,et al.Comparative analysis of external quality and nutritional components of poultry eggs[J].Animal Husbandry amp; Veterinary Medicine,2022,54(5):40-44.(in Chinese)
[4] MOGHTADERI M,NABAVIZADEH S H,HOSSEINITESHNIZI S.The frequency of cross-reactivity with various avian eggs among children with hen′s egg allergy using skin prick test results:fewer sensitizations with pigeon and goose egg[J].Allergol Immunopathol (Madr),2020,48(3):265-269.
[5] WANG Y,YANG H M,ZI C,et al.The mediation of pigeon egg production by regulating the steroid hormone biosynthesis of pigeon ovarian granulosa cells[J].Poult Sci,2020,99(11):6075-6083.
[6] TYASI T L,SUN X,SHAN X S,et al.Effects of RAC1 on proliferation of hen ovarian prehierarchical follicle granulosa cells[J].Animals (Basel),2020,10(9):1589.
[7] CUI Z F,SHEN X X,ZHANG X X,et al.A functional polymorphism of inhibin alpha subunit at miR-181b-1-3p-binding site regulates proliferation and apoptosis of chicken ovarian granular cells[J].Cell Tissue Res,2021,384(2):545-560.
[8] 郭振玉.產蛋間隔期鴿卵泡發育、生殖激素濃度及相關基因表達的研究[D].揚州:揚州大學,2023.
GUO Z Y.Study on follicle development,reproductive hormone synthesis and related genes expression patterns in white king pigeons during laying interval[D].Yangzhou:Yangzhou University,2023.(in Chinese)
[9] 茹 盟,曾文惠,彭劍玲,等.蛋雞卵泡發育及其表觀遺傳調控機制研究進展[J].畜牧獸醫學報,2023,54(9):3613-3622.
RU M,ZENG W H,PENG J L,et al.Research progress on follicles development of hens and its epigenetic regulatory mechanism[J]. Acta Veterinaria et Zootechnica Sinica,2023,54(9):3613-3622.(in Chinese)
[10] WEI Q Y,LI J,HE H R,et al.miR-23b-3p inhibits chicken granulosa cell proliferation and steroid hormone synthesis via targeting GDF9[J].Theriogenology,2022,177:84-93.
[11] 張俊珍,李彩娥,劉 博,等.不同生理階段邊雞卵巢轉錄譜的構建及卵泡發育相關基因的分析[J].畜牧獸醫學報,2022, 53(2):423-435.
ZHANG J Z,LI C E,LIU B,et al.Construction of ovary transcription profile of bian chicken at different physiological stages and analysis of genes related to follicular development[J].Acta Veterinaria et Zootechnica Sinica,2022,53(2):423-435.(in Chinese)
[12] YANG C,HUANG X B,CHEN S J,et al.The effect of heat stress on proliferation,synthesis of steroids,and gene expression of duck granulosa cells[J].Anim Sci J,2021,92(1):e13617.
[13] SUN X,CHEN X X,ZHAO J H,et al.Transcriptome comparative analysis of ovarian follicles reveals the key genes and signaling pathways implicated in hen egg production[J].BMC Genomics,2021,22(1):899.
[14] WANG Y,GUO Z Y,ZHANG C,et al.Characterization of ovarian follicles,serum steroid hormone concentration,and steroidogenic gene expression profiles in the developing ovarian follicles in White King pigeons[J].Poult Sci,2023, 102(7):102673.
[15] 任鈺為,陳 星,林燕寧,等.基于全基因組重測序研究文昌雞產蛋性能的影響因素[J].畜牧獸醫學報,2024,55(2):502-514.
REN Y W,CHEN X,LIN Y N,et al.Investigating the influencing factors of egg laying performance in Wenchang chickens based on whole genome resequencing[J].Acta Veterinaria et Zootechnica Sinica,2024,55(2):502-514.(in Chinese)
[16] VAN DE WEERDT B C M,MEDEMA R H.Polo-like kinases:a team in control of the division[J].Cell Cycle,2006,5(8):853-864.
[17] MA S,CHARRON J,ERIKSON R L.Role of PLK2 (Snk) in mouse development and cell proliferation[J].Mol Cell Biol,2003, 23(19):6936-6943.
[18] LI F X,JO M,CURRY JR T E,et al.Hormonal induction of polo-like kinases (PLKS) and impact of PLK2 on cell cycle progression in the rat ovary[J].PLoS One,2012,7(8):e41844.
[19] Belle R,PLUCHON P F,CORMIER P,et al.Identification of a new isoform of eEF2 whose phosphorylation is required for completion of cell division in sea urchin embryos[J].Dev Biol,2011,350(2):476-483.
[20] KALECI B,KOYUTURK M.Efficacy of resveratrol in the wound healing process by reducing oxidative stress and promoting fibroblast cell proliferation and migration[J].Dermatol Ther,2020,33(6):e14357.
[21] CHEN J Y,WANG J,WU X J,et al.eEF2 improves dense connective tissue repair and healing outcome by regulating cellular death,autophagy,apoptosis,proliferation and migration[J].Cell Mol Life Sci,2023,80(5):128.
[22] ZHANG J N,DUAN Z T,WANG X Q,et al.Screening and validation of candidate genes involved in the regulation of egg yolk deposition in chicken[J].Poult Sci,2021,100(6):101077.
[23] HU K,WANG C,XU Y F,et al.Interaction of lncRNA Gm2044 and EEF2 promotes estradiol synthesis in ovarian follicular granulosa cells[J].J Ovarian Res,2023,16(1):171.
[24] 鄭婉秋,李 爍,吳金峰,等.RNA解旋酶DDX5在病理生理作用中的研究進展[J].臨床與病理雜志,2023,43(3):623-627.
ZHENG W Q,LI S,WU J F,et al.Research progress in RNA helicase DDX5 in its pathophysiological effects[J].Journal of Clinical and Pathological Research,2023,43(3):623-627.(in Chinese)
[25] MAZUREK A,LUO W J,KRASNITZ A,et al.DDX5 regulates DNA replication and is required for cell proliferation in a subset of breast cancer cells[J].Cancer Discov,2012,2(9):812-825.
[26] 朱國勝,邱麗影,資 捷.DDX5和DDX17在卵巢癌組織中的表達及其臨床意義[J].醫學理論與實踐,2023, 36(10):1633-1635,1628.
ZHU G S,QIU L Y,ZI J.Expression of DDX5 and DDX17 in ovarian cancer tissues and their clinical significance[J].The Journal of Medical Theory and Practice,2023,36(10):1633-1635,1628.(in Chinese)
[27] 強 浩.綿羊IGF1R、IGF2R基因的多態性及其與生長性狀的關聯分析[D].銀川:寧夏大學,2022.
QIANG H.Polymorphism of IGF1R,IGF2R gene and its association analysis with growth traits in sheep[D].Yinchuan:Ningxia University,2022.(in Chinese)
[28] NIELSEN F C.The molecular and cellular biology of insulin-like growth factor II[J].Prog Growth Factor Res,1992, 4(3):257-290.
[29] WEBB R,CAMPBELL B K.Development of the dominant follicle:mechanisms of selection and maintenance of oocyte quality[J].Soc Reprod Fertil Suppl,2007,64:141-163.
[30] TKACHENKO O Y,WOLF S,LAWSON M S,et al.Insulin-like growth factor 2 is produced by antral follicles and promotes preantral follicle development in macaques[J].Biol Reprod,2021,104(3):602-610.
[31] NGUYEN P T T,CONLEY A J,SOBOLEVA T K,et al.Multilevel regulation of steroid synthesis and metabolism in the bovine placenta[J].Mol Reprod Dev,2012,79(4):239-254.
[32] NIE R X,ZHENG X T,ZHANG W H,et al.Morphological characteristics and transcriptome landscapes of chicken follicles during selective development[J].Animals (Basel),2022,12(6):713.
[33] LAVOIE H A,KING S R.Transcriptional regulation of steroidogenic genes:STARD1,CYP11A1 and HSD3B[J].Exp Biol Med (Maywood),2009,234(8):880-907.
[34] 楊安琪,李嘉誠,宋 穎,等.CYP19A1對兔卵巢顆粒細胞增殖和凋亡的影響[J].畜牧獸醫學報,2023,54(10):4209-4219.
YANG A Q,LI J C,SONG Y,et al.Effects of CYP19A1 on proliferation and apoptosis of rabbit ovary granulosa cells[J].Acta Veterinaria et Zootechnica Sinica,2023,54(10):4209-4219.(in Chinese)
[35] SECHMAN A,GRZEGORZEWSKA A K,GRZESIAK M,et al.Nitrophenols suppress steroidogenesis in prehierarchical chicken ovarian follicles by targeting STAR,HSD3B1,and CYP19A1 and downregulating LH and estrogen receptor expression[J]. Domest Anim Endocrinol,2020,70:106378.
[36] CONVISSAR S,WINSTON N J,FIERRO M A,et al.Sp1 regulates steroidogenic genes and LHCGR expression in primary human luteinized granulosa cells[J].J Steroid Biochem Mol Biol,2019,190:183-192.
(編輯 郭云雁)
