低溫對民豬骨骼肌線粒體能量代謝基因的影響
2024-07-12 12:36:41張冬杰馬守正汪亮李忠秋劉娣
西北農業學報 2024年7期
張冬杰 馬守正 汪亮 李忠秋 劉娣
doi:10.7606/j.issn.1004-1389.2024.07.001
https://doi.org/10.7606/j.issn.1004-1389.2024.07.001
收稿日期:2022-11-08? 修回日期:2023-02-08
基金項目:國家自然科學基金(32172696);黑龍江省省屬科研院所科研業務費項目(CZKYF2021-2-C025);國創中心先導科技項目(NCTIP-XD/C16)。
第一作者:張冬杰,女,研究員,研究方向為民豬資源保護研究與利用。E-mail:djzhang8109@163.com
通信作者:劉? 娣,女,博士,教授,研究方向為地方豬資源保護研究與利用。E-mail:liudi1963@163.com
摘? 要? 為了探究低溫環境下民豬肌肉組織內與線粒體能量代謝相關基因的變化情況,以解析民豬抗寒特性的遺傳機制。將9頭體質量相近的母豬隨機分成3組,每組3頭,在冬季將其中2組置于室外半敞篷舍內飼養,分別處理3 d(急性低溫處理組)和58 d(慢性低溫處理組),1組置于常規舍內飼養,作為對照組。以背肌為試驗材料,采用PCR array方法,對90個與線粒體能量代謝相關基因的表達水平進行檢測。結果表明,與對照組相比,急性低溫處理組基因的表達水平差異不顯著,但慢性低溫處理顯著改變? ATP5H、? ATP5L、? ATP6V1G3、? COX5A、? COX7A2、? COX7A2L、? NDUFS4、? SDHB、? UQCRQ和? GADD45B基因的表達水平,尤其是? GADD45B,達到了極顯著水平(P<0.01),這些基因均為編碼OXPHOS系統亞基基因,推測慢性低溫誘導了背肌ATP的生成。對? GADD45B基因進一步分析發現,該基因在哺乳動物中高度保守,基本符合分子進化規律,表達具有明顯的組織特異性。急性低溫處理對民豬背肌內ATP的生成影響不顯著,但慢性低溫處理誘導ATP的生成,以維持體溫恒定。
關鍵詞? 民豬;骨骼肌;低溫;線粒體;能量代謝
線粒體是細胞的能量工廠,通過氧化呼吸鏈為生物體提供95%的能量。線粒體氧化磷酸化過程中產生的能量一部分作為ATP直接被生物體利用以支持某些能量消耗過程,比如肌肉運動、胞內離子運輸等,剩余部分則主要作為熱能維持體溫。人們發現不同氣候條件下動物對熱量的需求并不相同,其線粒體受到純凈化選擇壓力也不同[1]。
適應性產熱(adaptive thermogenesis)對恒溫動物至關重要,是維持核心體溫恒定的關鍵組成部分。當機體遭遇低溫刺激時,除棕色脂肪外,骨骼肌也是重要的產熱組織。它一方面可以通過肌肉的舒張和收縮產生熱量,當細胞質中的Ca2+濃度升高時,肌肉會收縮,反之則舒張[2]。Ca2+的轉運主要由肌質網鈣離子ATP酶 (sarco/endoplasmic reticulum Ca2+-ATPase, SERCA) 完成,它在轉運Ca2+時,會水解一個ATP,釋放熱量,同時將兩個Ca2+從細胞質泵回肌質網[3]。另一方面,肌肉線粒體中受調節的氧化磷酸化解偶聯也可發揮產熱作用[4]。冷馴化可以增加骨骼肌中線粒體的體積,這可能是通過Ca2+刺激線粒體生物發生產生的[5]。
豬是較為特殊的一種哺乳動物,新生仔豬對溫度的敏感性要大于牛、羊等其他新生家畜。但豬的? UCP1基因在進化過程中發生了突變,使其無法產生傳統意義上的棕色脂肪[6]。因此,寒冷環境下,豬是通過何種方式維持體溫恒定一直是未知的。相比之下,同樣發生? UCP1基因突變而無法產生棕色脂肪的鳥類,其低溫環境下的能量代謝機制的研究較為透徹[7]。對于鳥類,已經明確骨骼肌是其在低溫環境下的重要產熱組織[8],但豬由于相關研究報道較少,目前還無法確認肌肉和脂肪哪個是其重要產熱組織,或者是兩者兼具。但無可爭議的是,長期冷暴露或短期急性冷暴露均會導致機體產熱能力的增加,這是恒溫動物維持體溫、調節內穩態的關鍵[9]。
本研究以一種東北地區特有的民豬作為研究對象,該地方豬因長期生活在寒冷地區,具有明顯的抗寒特性[10]。使用PCR array技術對遭受慢性低溫處理和急性低溫處理的民豬背肌進行與線粒體能量代謝相關基因的篩選與分析,以期解析低溫環境下民豬骨骼肌線粒體的能量代謝變化情況。
1? 材料與方法
1.1? 試驗材料
9頭體質量為(60±5) kg的民豬母豬由黑龍江省農業科學院畜牧研究所民豬養殖場提供。
1.2? 試驗設計
2020年11月9日,試驗開始前,9頭個體均在供暖的豬舍內飼養,舍內溫度控制在(18±2) ℃。將9頭個體隨機分成3組,每組3頭。試驗開始后,隨機將一組置于室外半敞篷舍內飼養,記為慢性低溫處理組,半敞篷舍內溫度與外界環境溫度基本一致,白天最高溫度為5 ℃,夜晚最低溫度為-5 ℃。試驗進行55 d后,將剩余2組的1組置于室外半敞篷舍內飼養,記為急性低溫處理組,此時白天最高溫度降為-15 ℃,夜晚最低溫度為-24 ℃。再處理3 d后,試驗結束。整個試驗期內,3組個體均自由采食,保證充足的飲水。試驗結束后,將9頭個體同時屠宰,采集背肌、背脂、心臟、肝臟、脾臟、肺、腎臟等組織,置于液氮中保存,備用。
1.3? 總RNA的提取與反轉錄
使用Trizol分別提取每個個體不同組織的總RNA,微量分光光度計進行純度檢測,1%瓊脂糖凝膠電泳檢測所提RNA的濃度和完整性,質量濃度要求不低于100 ng/μL。使用PrimeScript RT reagent Kit with gDNA Eraser試劑盒將所提取的總RNA反轉錄成cDNA。
1.4? PCR array分析
豬線粒體能量代謝PCR array孔板訂制于上海沃吉基因科技有限公司,每個PCR array為一個96孔板,內含90個目的基因和3個內參基因以及3個陰性對照。陣列中所含的90個目的基因信息見表1。3個內參基因分別為ACTB、GAPDH和? HPRT1。
1.5? 實時熒光定量PCR(qRT-PCR)
96孔板中含有目的基因的引物,因此只需配置其他qRT-PCR反應成分,包括cDNA樣品0.5 μL,WCGENE mRNA qPCR mix (2×)5.3 μL,滅菌水3.2 μL。反應程序為:95 ℃ 30 s;40個循環:95 ℃ 5 s,60 ℃ 30 s。檢測結果根據2-ΔΔCt法計算不同低溫處理組目的基因相較于對照組的表達量變化情況,按照|log2FC|>1且P<0.05的標準篩選差異基因。
1.6? ??GADD45B基因的克隆測序與分子進化樹構建
使用Primer 5.0軟件,根據NCBI數據庫中已有的? GADD45B基因序列(XM_005654701)設計引物,用于擴增該基因的完整編碼區,引物序列為GS:5′-ATGACACTGGAAGAGCTCGTGG-3′,GA:5′-CAGCGTTCCTGGAGGGAGAT-3′。獲得民豬? GADD45B基因的完整編碼區序列后,使用DNAMAN軟件與已知序列進行比對,確定所擴增序列的準確性。使用NCBI數據庫的在線軟件BLAST,下載人、鼠、恒河猴、牛、羊、雞、家燕、鱷魚、青蛙和斑馬魚的GADD45B氨基酸序列。使用MEGA 11.0軟件對這些氨基酸序列進行整理和比對,并構建基于GADD45B氨基酸序列的分子進化樹。
1.7? ?GADD45B基因的組織表達譜
采用qRT-PCR的方法,檢測? GADD45B基因在民豬的心臟、肝臟、脾臟、肺、腎臟、背肌和背脂中的表達水平。所用引物序列為AS:5′- GCTGATGAATGTGGACCCT -3′,AA:5′- AAACGACTGGATGAG GGTG -3′。選擇β-actin基因作為內參基因,引物序列為F:5′-CGGGACCTGACCGACTACCT-3′,S:5′- GGGCCGTGATC TCCTTCTG -3′。qRT-PCR反應條件和反應程序同“1.5”。將該基因在腎臟的表達水平設為對照,其他組織與其相比較,判定表達水平高低,相對表達量的計算公式為2-△△Ct,其中△△Ct=(△Ct目的基因-△Ct內參基因)其他組織-(△Ct目的基因-△Ct內參基因)心臟。
2? 結果與分析
2.1? 慢性低溫處理后肌肉組織線粒體能量代謝相關基因的變化
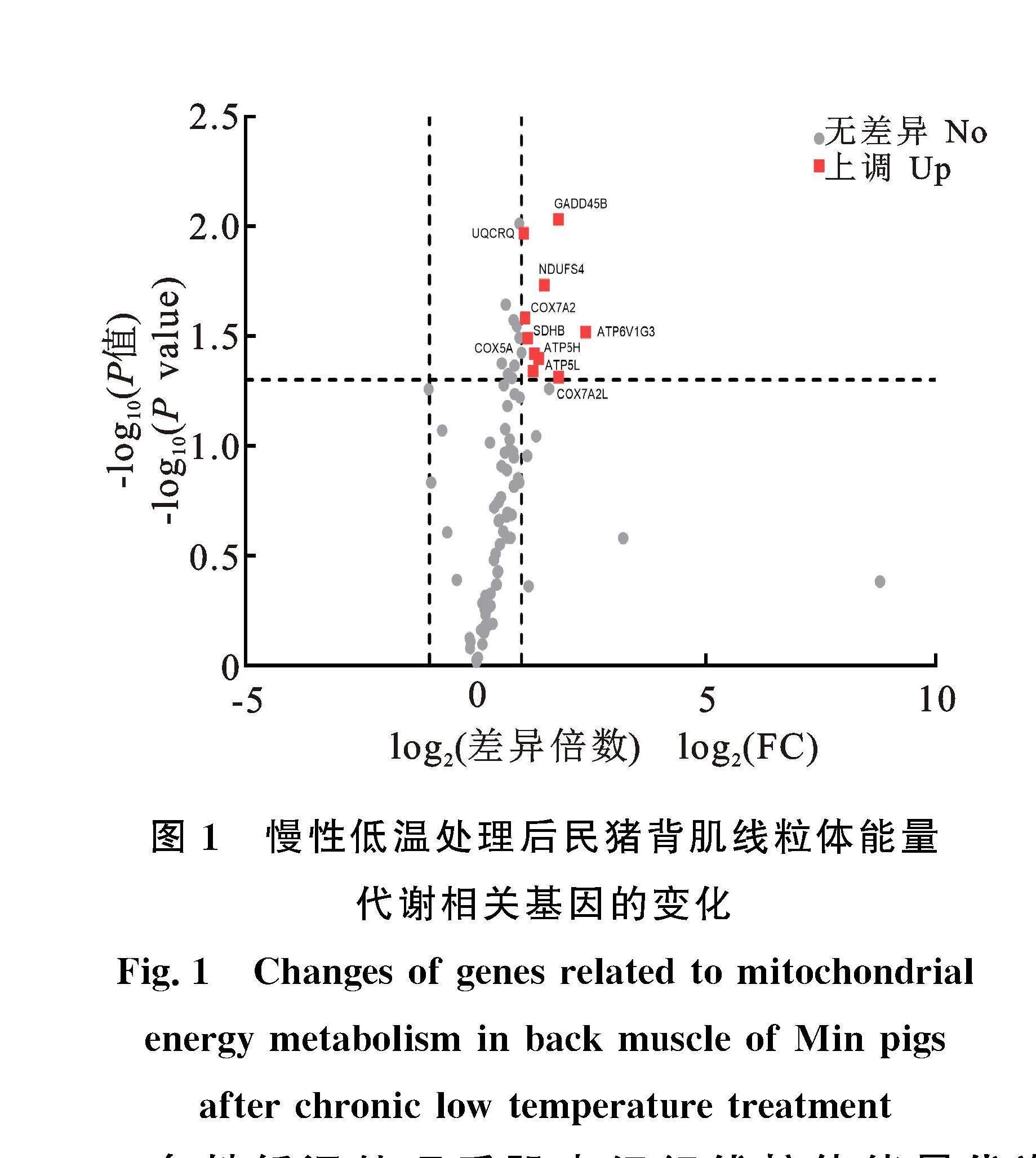
線粒體能量代謝陣列共包含90個目的基因,以ATP酶H+/K+轉運、ATP合成、細胞色素C氧化酶、 泛醌氧化還原酶(NADH)、琥珀酸脫氫酶復合體、泛醌細胞色素c還原酶等家族基因為主。民豬在遭受慢性低溫處理后,背肌內與線粒體能量代謝相關的90個基因,按照|log2FC|>1且P<0.05標準進行差異表達基因篩選,共10個基因發生顯著上調,其中? ATP5H、? ATP5L、? ATP6V1G3、? COX5A、? COX7A2、? COX7A2L、? NDUFS4、? SDHB和? UQCRQ顯著上調(P
2.2? 急性低溫處理后肌肉組織線粒體能量代謝相關基因的變化
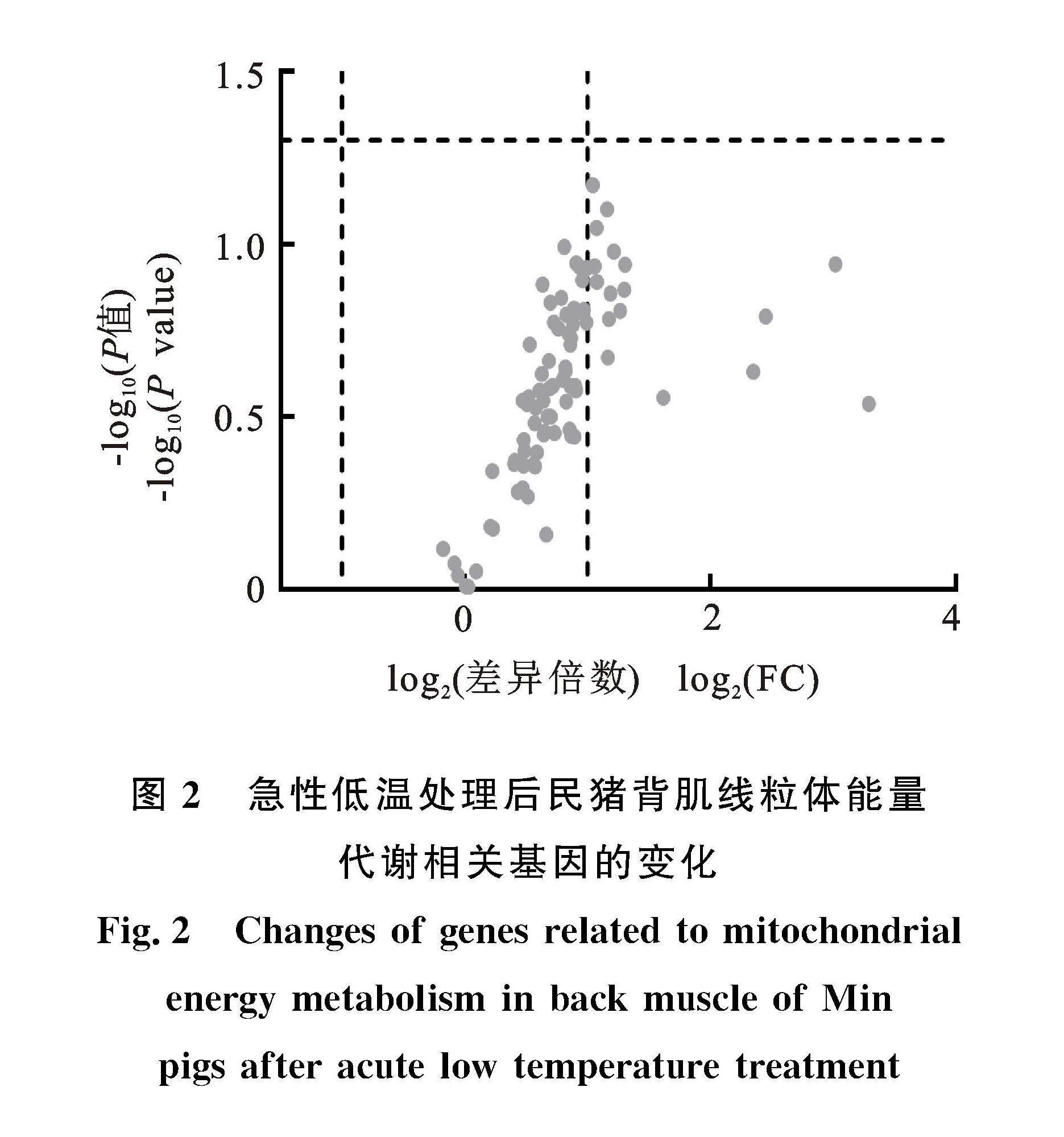
民豬在遭受急性低溫處理后,背肌內與線粒體能量代謝相關的90個基因,按照|log2FC|>1且P<0.05標準進行差異表達基因篩選,未發現發生顯著變化的基因(圖2)。說明急性低溫處理沒有顯著改變背肌內線粒體能量代謝。
2.3? ??GADD45B基因的克隆測序
使用引物對GS和GA,獲得民豬? GADD45B基因完整編碼區序列,共計483 bp,編碼160個氨
基酸。通過BLAST比對,下載NCBI數據庫中人(NP_056490.2)、鼠(NP_032681.1)、恒河猴(NP_001247803.1)、牛(NP_001035694.1)、羊(XP_004008677.1)、雞(XP_040548737.1)、家燕(XP_039942460.1)、鱷魚(XP_019393972.1)、青蛙(XP_040178585)和斑馬魚(NP_001012386)的GADD45B氨基酸序列。經比對發現,該基因在哺乳動物中高度保守,序列一致性達97.81%,在不同物種間屬于中度保守,序列一致性為? 81.99%(圖3)。
2.4? 基于GADD45B氨基酸序列構建分子進化樹
利用MEGA11.0軟件構建基于不同物種GADD45B氨基酸序列的分子進化樹(圖4)。哺乳動物中,豬與牛、羊的親緣關系最近,人和恒河
猴的親緣關系最近,哺乳動物聚在一個大的分支上。家燕和雞同為鳥類,它們的親緣關系最近,但斑馬魚、鱷魚和青蛙在進化樹上的位置與它們的進化地位并不相符。斑馬魚作為魚類,應處于該進化樹的最低端,其次是兩棲類的青蛙,然后是爬行類的鱷魚和鳥類。
2.5? ?GADD45B基因在不同組織的表達
以民豬的腎臟組織為參照,利用相對定量的計算方法,比較? GADD45B基因在民豬不同組織的表達情況(圖5)。發現該基因具有明顯的組織表達特異性,在背肌和心臟中表達量非常低,其次是背脂,但在脾臟中表達量非常高,肺臟和肝臟次之。
3? 討? 論
骨骼肌具有很強的代謝靈活性和適應能力,能夠對不同能量需求的環境信號作出反應[10]。骨骼肌細胞中的線粒體通過氧化磷酸化(oxidative phosphorylation,OXPHOS)產生細胞行使功能所需的大部分能量,并參與許多關鍵的細胞過程,如凋亡、鈣穩態和產生活性氧(reactive oxygen species,ROS)。熱應激會改變骨骼肌細胞的糖酵解,在ATP需求突然增加后,補償性糖酵解機制被激活。熱應激誘導的熱休克蛋白70 (HSP70)抑制OXPHOS,同時促進糖酵解以補償ATP失衡[11]。目前關于細胞和分子對冷脅迫適應的復雜機制研究較少。此前發現,將小鼠的C2C12成肌細胞暴露在低溫下會增加代謝通量,分化肌管通過增加代謝率、ATP產生和糖酵解通量[12]來應對低溫。
本研究發現,短期的急性低溫處理(3 d)并未顯著改變這些基因的表達水平,而長期的慢性低溫處理(58 d)顯著誘導了與ATP合成相關基因的表達。線粒體ATP的產生是由OXPHOS系統介導的,該系統由4個線粒體多亞基復合體(CⅠ、CⅡ、CⅢ和CⅣ)和F0F1-ATP合成酶(CⅤ)組成。民豬在遭受長期低溫處理后,多個參與編碼該系統亞基的基因發生了顯著上調,其中? NDUFS4是編碼CⅠ亞基的基因,CⅠ是4個復合體中最大最復雜的一個蛋白,線粒體內大多數ATP是經過CⅠ的電子傳遞產生的[13]。在乳鼠的心肌細胞中敲減? NDUFS4,會造成ROS水平上升,線粒體膜電位、鈣離子攝取能力、細胞最大呼吸速率均出現下降,說明該基因在維持線粒體正常功能方面具有重要作用[14]。上調表達的? SDHB是編碼CⅡ亞基的基因,它既可參與ATP的生成,也可影響細胞內線粒體的數量[15]。編碼CⅢ亞基的? UQCRQ基因,編碼CⅣ亞基的? COX5A、? COX7A2和? COX7A2L基因,也發生了顯著上調。CⅣ負責將電子從細胞色素C傳遞到分子氧。電子傳遞與ATP合成是偶聯在一起的,即氧化磷酸化過程。此外,編碼F0F1-ATP合成酶亞基的3個基因,即? ATP5H、? ATP5L和? ATP6V1G3也發生了顯著上調,ATP合成酶在呼吸過程中通過電子傳遞鏈(electron transport chain,ETC)釋放的能量先轉換為跨膜質子(H+)梯差,之后質子流順質子梯差通過ATP合成酶使ADP+Pi合成ATP[16]。這些基因的顯著上調,說明低溫處理下,民豬骨骼肌線粒體內的ATP合成增加了。
對發生極顯著上調表達的? GADD45B基因進一步分析發現,該基因在正常情況下的脾臟中高表達,這與其參與TH1介導的免疫應答相關,該基因的缺失,會導致小鼠脾臟顯著增大[17]。本研究中,民豬在遭受慢性低溫處理后,骨骼肌中的? GADD45B基因發生了極顯著的上調,具體作用機理未知。目前已知? GADD45B是肌肉發生的重要調控因子,它的缺失可降低p38 MAPK的磷酸化水平,從而下調肌肉調節因子(muscle regulatory factors,MRFs)和過氧化物酶體增殖物激活受體γ輔激活因子1Α(PPARGC1A)的表達,導致肌源性分化和線粒體生物發生的抑制[18]。此外,該基因還參與細胞的DNA修復和抗凋亡。
4? 結? 論
民豬是一個具有明顯抗寒特性的地方豬種,通過PCR array技術高通量篩選了不同程度的低溫處理下民豬骨骼肌線粒體能量代謝相關基因的變化情況。發現急性低溫處理并未改變民豬骨骼肌內線粒體能量代謝相關基因的表達,但慢性低溫處理誘導了多個編碼OXPHOS系統的亞基基因,包括? ATP5H、? ATP5L、? ATP6V1G3、? COX5A、? COX7A2、? COX7A2L、? NDUFS4、? SDHB和? UQCRQ基因(P<0.05),以及發生極顯著變化的? GADD45B基因(P<0.01),推測這些基因的上調表達將最終實現ATP合成的增加。發生極顯著變化的? GADD45B基因,氨基酸序列在哺乳動物中高度保守,基本符合分子進化規律,表達具有明顯的組織特異性。
參考文獻? Reference:
[1]? SUN Y B,SHEN Y Y,IRWIN D M,et al.Evaluating the roles of energetic functional constraints on teleost mitochondrial-encoded protein evolution[J].Molecular Biology and Evolution,2011,28(1):39-44.
[2] ?MACLENNAN D H,KRANIAS E G.Phospholamban: a crucial regulator of cardiac contractility[J].Nature Reviews Molecular Cell Biology,2003,4(7):566-577.
[3]? TOYOSHIMA C,NAKASAKO M,NOMURA H,et al.Crystal structure of the calcium pump of sarcoplasmic reticulum at 2.6 A resolution[J].Nature,2000,405(6787):647-655.
[4]? MAHALINGAM S,CHEVIRON Z A,STORZ J F,et al.Chronic cold exposure induces mitochondrial plasticity in deer mice native to high altitudes[J].The Journal of Physiology,2020,598(23):5411-5426.
[5]? MAURYA S K,HERRERA J L,SAHOO S K,et al.Sarcolipin signaling? promotes? mitochondrial biogenesis and oxidative? metabolism in skeletal? muscle[J].Cell Reports,2018,24(11):2919-2931.
[6]? HOU L,SHI J,CAO L,et al.Pig has no uncoupling protein 1[J].Biochemical and Biophysical Research Communication,2017,487(4):795-800.
[7]? SWANSON D L,ZHANG Y,JIMENEZ A G.Skeletal muscle and metabolic flexibility in response to changing energy demands in wild birds[J].Frontiers in Physiology,2022,3:961392.
[8]? NOWACK J,GIROUD S,ARNOLD W,et al.Muscle non-shivering thermogenesis and its role in the evolution of endothermy[J].Frontiers in Physiology,2017,8:889.
[9]? TATE K B,WEARING O H,IVY C M,et al.Coordinated changes across the O2 transport pathway underlie adaptive increases in thermogenic capacity in high-altitude deer mice[J].Proceedings? Biological Sciences,2020,287(1927):20192750.
[10]? 楊月瑩,劉 ?娣.民豬耐寒基因目標風險最小特征搜索[J].西北農業學報,2013,22(10):15-21.
YANG Y? Y,LIU D.Screening for anti freezing genes in pig with target risk minimized feature searching algorithm[J].Acta Agriculturae Boreali-occidentalis Sinica,2013,22(10):15-21.
[11]? WESTERBLAD H,BRUTON J D,KATZ A.Skeletal muscle:energy metabolism,fiber types,fatigue and adaptability[J].Experimental Cell Research,2010,316:3093-3099.
[12]? SAJJANAR B,SIENGDEE P,TRAKOOLJUL N,et al.Cross-talk between energy metabolism and epigenetics during temperature stress response in C2C12 myoblasts[J].International Journal of Hyperthermia,2019,36(1):776-784.
[13]? LITTLE A G,SEEBACHER F.Thermal conditions experienced during differentiation affect metabolic and contractile phenotypes of mouse myotubes[J].American Journal of Physiology Regulation,Integrative and Comparative Physiology,2016,311: R457-R465.
[14]? 李鳳杰,沈麗君,方合志,等.線粒體呼吸鏈復合體Ⅰ[J].中國細胞生物學學報,2014,36(8):1153-1161.
LI F J,SHEN L J,FANG H ZH,et al.Mitochondrial respiratory? complex I[J].Chinese Journal of Cell Biology,2014,36(8):1153-1161.
[15]? 李? 杰,王愛玲,楊謀廣.低表達NADH脫氫酶[輔酶Q]鐵硫蛋白4對乳鼠心肌細胞線粒體功能的影響[J].安徽醫學,2017,38(8):959-962.
LI J,WANG A L,YANG M G.Effect of low expression of protein?? NDUFS4 on mitochondria function of ventricular myocytes in neonatal rats[J].Anhui Medical Journal,2017,38(8):959-962.
[16]? 陳立蘭,狄? 文.琥珀酸脫氫酶B亞基對卵巢癌細胞線粒體DNA拷貝數影響的研究[J].現代婦產科進展,2022,31(3):161-165.
CHEN L L,DI W.Study on the effect of succinate dehydrogenase B subunit on the mitochondrial DNA copy number in ovarian cancer cells[J].Progress in Obstetrics and Gynecology,2022,31(3):161-165.
[17]? ZHAO R Z,JIANG S,ZHANG L,et al.Mitochondrial?? electron transport chain,ROS generation and uncoupling (Review)[J].International Journal of Molecular Medicine,2019,44(1):3-15.
[18]? DENG K,FAN Y,LIANG Y,et al.FTO-mediated demethylation of?? GADD45B promotes myogenesis through the activation of p38 MAPK pathway[J].Molecular Therapy-Nucleic Acids,2021,26:34-48.
Effect of Low Temperature on Mitochondrial Energy Metabolism Genes in Skeletal Muscle of Min Pig
ZHANG Dongjie, MA Shouzheng, WANG Liang, LI Zhongqiu and LIU Di
(Institute of Animal Husbandry, Heilongjiang Academy of Agricultural Science,Harbin? 150086,China)
Abstract? In order to explore the changes of genes related to mitochondrial energy metabolism in muscle tissue of Min pigs under low temperature environment, and to analyze the genetic mechanism of cold resistance of Min pigs.Nine female Min pigs with similar mass? were randomly divided into three groups, with three individuals in each group.In winter, two groups of individuals were raised in outdoor semi-open houses for 3 days (acute low temperature treatment group) and 58 days (chronic low temperature treatment group), respectively, and one group was raised in normal houses as the control group.The expression level of 90 genes related to mitochondrial energy metabolism in the skeletal muscle of Min pig was detected by PCR array method.The acute low temperature treatment did not significantly change the expression levels of these genes, but chronic low temperature treatment significantly changed the expression levels of?? ATP5H,?? ATP5L,?? ATP6V1G3,? ?COX5A,?? COX7A2,?? COX7A2L,?? NDUFS4,?? SDHB,?? UQCRQ and?? GADD45B genes at a very significant level (P<0.01), especially?? GADD45B.These genes were all genes encoding OXPHOS system subunits,? suggesting that chronic low temperature treatment induced ATP production.Further analysis of?? GADD45B gene showed that the gene was highly conservative in mammals, basically in line with the molecular evolutionary law, and its expression had obvious tissue specificity.Acute low temperature treatment did not change the ATP production in skeletal muscle of Min pig, but the chronic low temperature treatment induced ATP production to maintain body temperature constant.
Key words? Min pig; Skeletal muscle; Low temperature; Mitochondrial; Energy metabolism
Received ??2022-11-08??? Returned? 2023-02-08
Foundation item? The National Natural Science Foundation of China (No.32172696); Scientific Research Fund for Heilongjiang Provincial Research Institutions (No.CZKYF2021-2-C025); National Center of Technology Innovation for Pigs (No.NCTIP-XD1C16).
First author? ZHANG Dongjie, female, research fellow.Research area: research and protection of the Min pig.E-mail: djzhang8109@163.com
Corresponding?? author? LIU Di, female, Ph.D,professor.Research area: research and protection of the indigenous pig breeds.E-mail:liudi1963@163.com
(責任編輯:顧玉蘭? Responsible editor:GU Yulan)
