高寒氣候區生長季NDVI與晝夜不對稱增溫的Copula分析
2024-06-28 10:04:06李忠良何光鑫李勛
大氣科學學報 2024年3期
李忠良 何光鑫 李勛


摘要 利用1982—2016年的青海地區歸一化植被指數和氣象數據,基于馬爾科夫鏈蒙特卡羅的Copula函數方法,深入探索晝夜增溫不對稱性與植被活動之間的復雜關系,揭示了晝夜增溫和NDVI之間的聯合概率分布及其季節性差異。研究結果表明,晝夜增溫與NDVI之間的關系在不同季節呈現顯著差異。尤其在秋季,晝夜增溫對NDVI的影響最為顯著,其次是夏季和春季。通過Copula函數模型,發現晝夜增溫與NDVI在特定溫度區間內呈現正相關,表明適宜的溫度條件下晝夜增溫對植被生長具有促進作用。然而,當晝夜增溫超過某一閾值時,其對NDVI的促進作用轉變為抑制作用,從而限制了植被的生長。同時,還揭示了重現期與晝夜增溫及NDVI之間的關系。在較低的重現期下,晝夜增溫與NDVI的聯合概率較高,表明在這些條件下,植被生長良好的情況出現的頻率較高。反之,較高的重現期對應于晝夜增溫與NDVI較低的聯合概率,表明植被生長受到抑制。本研究通過Copula函數提供了一個全新的視角來理解晝夜增溫與植被動態之間的相互作用,強調了氣溫變化對植被生長影響的復雜性。
關鍵詞晝夜增溫;歸一化植被指數(NDVI);非對稱性增溫;Copula;重現期
氣候變暖導致極端氣候事件頻發,使得陸地生態系統對全球變化的響應備受關注(Gong et al.,2017)。植被作為連接大氣、土壤水分和能量循環的重要組成部分(Kalisa et al.,2019),通過生物物理和地球化學活動,在調節區域和全球氣候變化中發揮重要作用(Chen et al.,2018)。歸一化植被指數(Normalized Difference Vegetation Index,NDVI)作為表征地表植被生長狀況的重要指標,是地表環境與氣候變化的指示器。研究植被的動態變化及其對氣候變化的響應,對于深入理解地表輻射平衡(Silva et al.,2017;Zhang et al.,2020)、碳氮循環(Del Grosso et al.,2018)、維持生態系統穩定性(Lu et al.,2019)以及動態監測陸地生態系統變化(He et al.,2017)等方面具有重要意義。溫度作為控制植物生理活動的重要因素之一(Cong et al.,2017;Zhang H X et al.,2018),近幾十年由于生長季溫度升高導致北半球植被生態系統的光合作用能力增強更為顯著(Wu et al.,2016;Reich et al.,2018;Park et al.,2019)。由于地理要素的復雜性以及氣候要素間相互作用,全球變暖存在明顯的不一致性(Davy et al.,2017),表現為晝夜增溫速率非對稱性和季節增溫速率不一致性。生長季的氣溫變化與植被活動之間存在緊密聯系(Piao et al.,2006;Kong et al.,2017),對植被的影響隨著晝夜交替表現出時間上非一致性的特征。全球大部分地區夜間增溫速率快于白天,季節上的增溫也存在同樣特征(Karl et al.,1993;Easterling et al.,1997;Davy et al.,2017;Barton and Schmitz,2018)。北半球高緯度地區夏季增溫速率大于春季和秋季,這種非對稱性變暖在未來幾十年會持續增強(Su et al.,2015)。晝夜溫度對植被生理活動的影響機制不同,其增溫的非對稱性對植被的影響程度不同,植被活動對溫度的變化表現出不確定性(Shen et al.,2018;Deng et al.,2019),溫度的變化特征如何影響植物的生理活動亟須進一步研究。
關于氣候變化對植被影響的研究主要集中于氣候的平均態對植被的影響(劉向培等,2015;Zhang Y L et al.,2018;Li et al.,2019;Meng et al.,2020)。氣候變暖使得極端氣候事件對NDVI的影響逐漸增強(Tan Z Q et al.,2015;Marcelino et al.,2020),溫度極值對植被變化的影響具有區域差異性和多樣性(Iglesias et al.,2012;Tan J G et al.,2015;Baumbach et al.,2017;Treharne et al.,2020)。多數學者基于傳統線性方法研究氣候因子與NDVI的相互作用關系,但此方法不能較好地反映時間序列的非平穩、非線性特征(Baumbach et al.,2017),在環境空間異質性較大的地區此問題更加凸顯(Miller,2012)。
大多數植物的光合作用發生在白天,但呼吸作用晝夜持續進行,晝夜溫度及其增溫速率的不一致性對植被的生理活動的影響是不同的。近50年地表夜間增溫速率為白天的1.4倍(Peng et al.,2013),晝夜增溫速率的差異性使得植被的固碳活動也呈現出非對稱性(Atkin et al.,2013)。晝夜增溫通過影響植被生態系統的結構、功能和過程,加強了植被對晝夜增溫速率不對稱性的響應(Xia et al.,2018;Du et al.,2019;Ma et al.,2019a)。目前,關于晝夜增溫速率不對稱性對植被動態影響的研究較少(Xia et al.,2018),且忽略了季節的不一致性對植被的影響(Rossi and Isabel,2017;Wang et al.,2020),可能掩蓋植被對晝夜增溫響應的季節性差異。因此,晝夜增溫的非對稱性及其季節性差異對植被活動影響的研究仍然較為匱乏。
青海是我國生物物種形成和演化中心之一,是氣候變化的“敏感區”和生態環境的“脆弱區”(Chang et al.,2018;Tang et al.,2018)。青海特殊的高原氣候和環境,使得高原植被的變化能夠比其他地區更快速地揭示全球氣候變化。由于地理和社會環境的差異,不同地區植被覆蓋動態變化特征有所不同。全球變暖加劇了區域的晝夜增溫變化差異,由此使得植被對晝夜增溫的響應在區域尺度上表現得更為明顯。深入探討高寒氣候區域的植被對晝夜增溫不對稱性的響應,可以豐富全球變化的區域響應研究,對于了解區域生態環境的變化趨勢,進而制定合理的生態環境保護政策和措施具有一定的參考價值。同時,有助于加深理解晝夜增溫對農作物的作用機制,對于改進作物模型也提供了理論支撐。迄今為止,青海地區長時間尺度的植被動態變化及其對極端氣候的響應尚不明確。本文應用Copula方法分析其NDVI時空動態變化,研究其對晝夜增溫非對稱性和季節性增溫不一致性的響應,揭示了晝夜增溫的非對稱性對植被活動的控制機制,為深入理解地表水熱過程及預測植被動態變化提供科學依據,對認識該地區生態系統演變、保護當地生態環境等具有重要意義。
1 研究區與研究方法
1.1 研究區概況
青海地處青藏高原東北部,位于89°35′~103°04′E、31°9′~39°19′N,是長江、黃河、瀾滄江的發源地,享有“中華水塔”的美譽。地勢由西向東呈階梯型下降趨勢,平均海拔3 000 m。青海省屬高原大陸性氣候,具有氣溫低、晝夜溫差大、降雨少而集中、日照長、太陽輻射強等特點。年平均氣溫在-5.6~8.6 ℃,降水量從東南向西北遞減,季節分配不均勻。受地形和氣候條件限制,植被類型多以高寒灌叢、高寒草甸及高寒草原為主,荒漠和山地草原次之,森林植被則比較少。
1.2 數據及預處理
1.2.1 氣象數據預處理
研究采用青海1982年1月1日—2016年12月31日的氣溫數據,包括最高氣溫和最低氣溫2個要素。根據氣象數據時間序列長度,數據完整性(缺失值小于5%)和空間分布情況,選擇青海省內氣象站點。通過質量控制測試識別所有站點的異常值,并以質量控制標志進行標記。
根據青海省植被生長規律,將生長季時段內的4—5月、6—8月、9—10月分別劃分為春季、夏季和秋季,大致對應植被從變綠到成熟,成熟到衰老,衰老到休眠3個生理過程(Tan J G et al.,2015)。考慮到大部分地區每日極端最高氣溫出現在14:00(北京時,下同)左右,而每日極端最低氣溫出現在日出前,因此分別將最高氣溫(Tmax)、最低氣溫(Tmin)作為晝夜溫度。
1.2.2 NDVI數據預處理
NDVI數據的時間跨度為1982—2016年,分別采用AVHRR數據和MODIS數據。其中,AVHRR數據來自美國航空航天局戈達德太空中心,數據時間跨度為1982—2010年。MODIS地表反射率數據來自NASA MODIS陸地產品組開發的Terra衛星每日地表反射率產品MOD09GA,本研究使用了2010—2016年生長季(4—10月)MOD09GA地表反射率數據。
對于AVHRR數據,通過輻射定標、云檢測與過濾、大氣校正及雙向反射率分布函數(BRDF)處理后生成逐日格網(5 km)數據集。對于MOD09GA數據,由于AVHRR NDVI和MODIS NDVI數據的空間分辨率不同,為了與LTDR數據時空相匹配,將MODIS 1-2波段地表反射率數據空間分辨率重采樣為5 km。然后經過質量檢驗、圖像鑲嵌、子集提取、裁剪、格式和投影轉換等預處理過程,得到MODIS NDVI數據集。這樣就把AVHRR數據集的時間跨度延長為1982—2016年。為了盡可能消除每日NDVI圖像中云或噪聲的影響,通過最大合成法將LTDR和MODIS逐日NDVI分別合成為逐旬NDVI數據。
1.3 方法介紹
1.3.1 最大值合成法
最大值合成法(Maximum Value Composite,MVC)由Holben(1986)提出,MVC具體公式如下:
NDVIi=max(NDVIij)。(1)
式中:NDVIi是指第i個月或者第i年的NDVI;NDVIij是指第i月的第j個時間段的NDVI數據或者第i年的第j月的NDVI數據。
1.3.2 Copula函數理論
Copula函數由Sklar(Genest and Favre,2007)提出,它可以利用邊緣分布和相關性框架構建多維聯合分布Copula函數模型(Li et al.,2013)。在眾多的刻畫兩個變量相關關系的統計方法和模型中,選擇以Copula函數來刻畫變量之間的相依關系,是因為Copula函數具有明顯的優勢:Copula函數擬合的變量相依關系可以無須假定變量符合某些統計分布特征;Copula給出的是變量之間的聯合概率分布函數,而非兩者之間簡單的相關系數,這有利于細致研究變量在不同區間上的動態相關關系;Copula函數可以獲得兩個變量在極值區域上的相關關系,為討論變量之間極值相關性提供了途徑。研究選取了8種Copula函數簇(Genest and Rivest,1993;Sraj et al.,2015;Sadegh et al.,2017,2018):1)BB1(Ben Nasr and Fateh,2019);2)Clayton(Clayton,1978);3)Frank(Li et al.,2013);4)Gaussian(Li et al.,2013);5)Gumbel(Li et al.,2013);6)Joe(Li et al.,2013);7)t(Li et al.,2013);8)Tawn(Gródek et al.,2019),這8種形式的Copula函數由于其性能,一直是相關模型的常用選擇。利用選取的Copula函數建立氣候要素與NDVI的二維聯合分布。
利用Copula聯合概率分布研究兩個變量之間的關系,需要經過參數估計(Chen and Sun,2015)、最優Copula函數簇的選擇和評估、相關性分析和估計單變量的邊緣分布函數(Singh et al.,2017;Sadegh et al.,2018)、構建雙變量聯合概率分布(Chen and Sun,2015)和回歸期的計算(Singh and Zhang,2018)等5個步驟。
2 結果分析
2.1 季節性晝夜增溫和NDVI趨勢
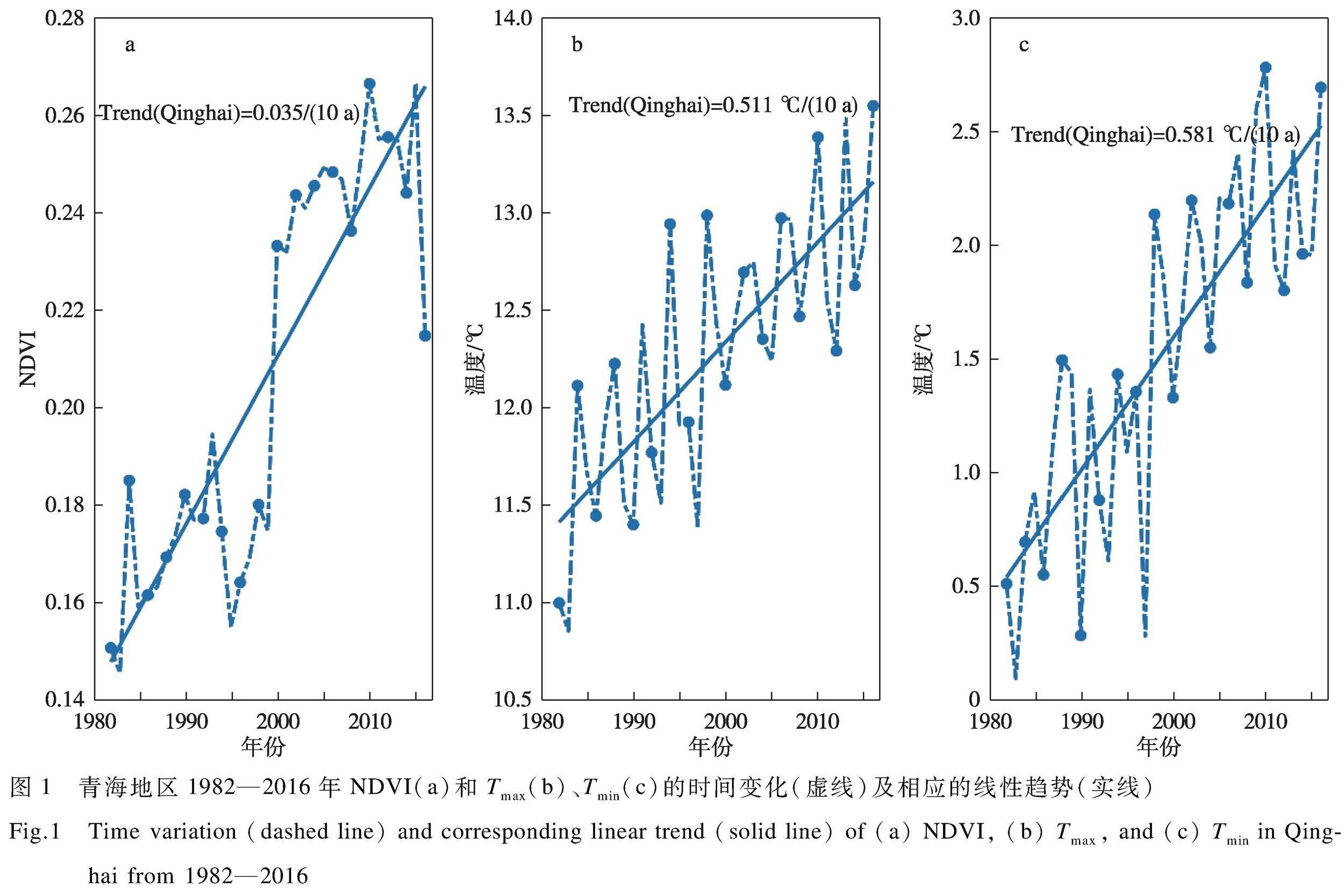
青海省獨特的自然環境決定了其植物區系和植被類型特征,境內植被具有典型的高寒性特點,草地為當地的主要植被類型,生態系統的結構和功能較為單一。因此,其典型的高原高寒氣候對植被的影響與其他區域具有明顯差異。圖1展現了青海省1982—2016年生長季NDVI和晝夜溫度變化情況。
近35年青海省植被表現為明顯增高趨勢,NDVI增長速率達到0.035 (10 a)-1,說明當地生態環境呈現逐漸好轉的發展態勢。青海NDVI年際變化較大,但波動較小。在2000年左右,NDVI發生了一個明顯的躍遷,這可能與當時全球變暖加劇有關。青海省晝夜溫度均呈現出增加趨勢,而且表現為晝夜增溫的不對稱性,最低溫的增加速率大于最高溫。其中,最高溫的增溫速率為0.511 ℃/(10 a),最低溫的增溫速率為0.581 ℃/(10 a),最低溫增溫速率為最高溫的1.14倍。青海晝夜溫度的年際變化較大,且波動也較大,受全球變暖影響明顯。
圖2為青海省1982—2016年生長季時段內不同季節的NDVI變化趨勢。NDVI表現為夏季最高,其次為秋季,春季次之。NDVI的增長速率由大到小依次為夏季>春季>秋季。不同季節的NDVI表現出明顯的年際波動,春季尤為劇烈,其標準差和變異系數均很大;而秋季的年際波動次之,夏季的年際波動較為平緩。主要是由于青海的水熱條件所決定,當地的景觀以草地為主,夏季植物生長茂盛,增長相對其他季節不明顯。而春季氣溫波動較大,生長季的開始和結束時間會發生明顯的振蕩。此外,NDVI的飽和效應對于夏季NDVI波動幅度較小也會產生很大影響。總體而言,青海地區各季節NDVI呈現增長趨勢,且在2000年前后產生了巨大躍遷。
圖3為青海地區1982—2016年NDVI突變識別檢驗的結果,可知,青海地區的NDVI整體上呈現先下降后上升的變化趨勢,其中1997年是NDVI產生突變的年份。植被生長所需的能量來自太陽輻射,受緯度地帶分異的影響,其生長狀況和分布格局受氣候變化的影響較大。通過研究全球區域內NDVI與氣候的相關性,發現溫度對NDVI的變化有顯著影響,特別在北半球中緯度以上地區,存在著季節尺度上的春、秋季的NDVI受溫度影響尤為顯著。小范圍的氣候突變不易引起局地的NDVI發生突變,而大范圍的氣候突變現象,例如厄爾尼諾事件的發生會導致NDVI產生突變。1998年發生厄爾尼諾/南方濤動(ENSO)事件,全球溫度大幅度升高。中國東部沿海地區冬季和春季溫度的升高會促進植物的光合作用、延長植被的生長期,有利于植被的生長發育和營養物質的累積。因此,青海地區的NDVI于1997年發生了巨大的躍遷。
由圖4可見,青海省不同季節的Tmax均表現為明顯上升趨勢,Tmax增溫幅度呈現明顯差異,在春、夏、秋3季的增長率分別為0.602 ℃/(10 a)、0.529 ℃/(10 a)和0.392 ℃/(10 a),增溫速率表現為春季>夏季>秋季。同理,Tmin在各個季節均表現為顯著增長趨勢,其增長率分別為0.564 ℃/(10 a)、0.652 ℃/(10 a)和0.491 ℃/(10 a),增溫速率由大到小依次為夏季>春季>秋季。不同季節的晝夜增溫速率呈現出季節差異性。青海地區春、夏和秋的Tmin與Tmax增溫速率之比分別為0.9、1.23和1.25。3個地區晝夜增溫在不同季節均表現出不對稱變化特征,存在明顯的季節性差異。
圖5為青海地區春、夏、秋3季的晝夜溫差變化趨勢可知,青海省晝夜溫差的變化呈現出季節性差異,春、夏、秋3季的晝夜溫差波動很大。春季的晝夜溫差呈上升趨勢,增加速率為0.038 ℃/(10 a),而夏季和秋季的晝夜溫差則表現為相反的變化規律,其中夏季晝夜溫差的下降速率大于秋季。這表明春季晝夜溫差逐漸加大,而夏秋兩季晝夜溫差逐漸縮小。結合圖1和圖4可知,近35年來青海最高溫的升溫速率小于最低溫。春季白晝增溫速率大于夜間,夏季和秋季則是夜間增溫速率大于白晝。
表1為青海不同季節晝夜增溫差異與NDVI的相關系數。所有季節的晝夜溫差和NDVI的相關關系均通過了統計學顯著性檢驗。春季和秋季的晝夜溫差與NDVI呈現負相關,夏季的晝夜溫差與NDVI呈現正相關。由于青海的高寒氣候特征,加之海拔較高,在長期的生態演化中其植物生長的溫度三基點較低。夏季由于輻射強,日照時數長,地表的熱通量大,溫度就處于植被適宜生長的溫度區間。晝夜溫差大能更好地滿足植物生長的溫周期需求。秋季由于夜間增溫為白天的1.25倍,導致夜溫較高,增加了呼吸作用對有機物的消耗,不利于根的生長和細胞分裂素的合成,降低了整株植物的生長速率。
2.2 構建最高溫、最低溫和NDVI Copula函數簇
本文分別對青海地區不同季節最高溫、最低溫和NDVI進行相關分析,并采用Kendall秩相關系數、Pearson相關系數、Spearman秩相關系數3種方法度量變量間的相關性,計算結果見表2和表3,均通過P<0.01統計學顯著性檢驗。
根據表2和表3可知,青海地區春季、夏季和秋季晝夜增溫與NDVI的相關性均呈現為極顯著正相關關系(P<0.01)。最高溫、最低溫和NDVI相關性強弱呈現出秋季>夏季>春季的規律。各季節最高溫和NDVI的相關性都強于最低溫和NDVI的相關性。由此可知,晝夜增溫在整體上對青海地區NDVI的影響程度較強,尤其是秋季晝夜增溫對NDVI影響最大。白天增溫和夜間增溫對青海地區植被的影響差異顯著,白天增溫影響強于夜間增溫。
不同單變量邊緣分布函數組合擬合Copula函數時,需要對Copula函數結構進行優選,利用AIC、BIC和RMSE3個標準選擇晝夜溫度與NDVI的最優Copula函數簇。
由表4可知,青海地區3個季節的Tmax-NDVI最優Copula函數分別為BB1、BB1和Frank,它們對應的3種評價指標數值均小于其余Copula函數的評價指標數值。同理,對于Tmin-NDVI最優Copula函數分別為BB1、BB1和Frank,說明這些Copula函數的擬合效果最好,更適合描述Tmax-NDVI、Tmin-NDVI的聯合分布特征。因此,本文選用BB1 Copula、BB1 Copula和Frank Copula函數分別建立3個季節最高溫、最低溫和NDVI的二維聯合概率分布模型。
2.3 最高溫、最低溫和NDVI聯合概率分布特征
根據最優Copula函數,分別計算青海地區生長季時段內春季、夏季和秋季的Tmax-NDVI以及Tmin-NDVI的聯合概率分布及其重現期。
圖6顯示了青海1982—2016年生長季內春、夏和秋季最高溫和NDVI之間的聯合概率分布關系。從圖6中可以獲取任意點最高溫與NDVI的聯合概率。聯合概率分布函數可以清晰反映最高溫和NDVI在不同值域范圍上的相關關系。圖中最顯著的特征是數據的不對稱和傾斜的依存結構,并且Tmax、Tmin取極小值時對NDVI影響較小,取極大值時對NDVI影響顯著。
根據等值線圖可以得出聯合概率為0.1~0.9時最高溫與NDVI的區間分布情況。當最高溫取固定值,NDVI越大,或NDVI取值固定,最高溫越大,其聯合概率越大。同一等值線上不同區間的最高溫和NDVI的聯合概率也呈現明顯差異。出現最高溫極小值和NDVI極小值、最高溫極小值和NDVI極大值以及最高溫極大值和NDVI極小值這3種情景的聯合概率較小;而出現最高溫極大值和NDVI極大值的聯合概率較大。當青海春、夏和秋季最高溫位于分別位于10~14、17~24和12~20 ℃區間時,NDVI值較大的概率較高(大于0.6)相對較大,表明此時植被生長相對良好。
圖7為青海春季、夏季和秋季Tmax-NDVI聯合超越概率的等值線圖。根據等值線圖可以獲取最高溫與NDVI為任意值時的聯合超越概率。圖中顯示了聯合超越概率分別為0.1~0.9時最高溫與NDVI同時大于等于某一特定值的不同組合情況。最高溫與NDVI的取值越小,其聯合超越概率越大;反之亦然。說明最高溫與NDVI同時超過較小值發生的概率,大于同時超過較大值時的概率。綜上表明植物生長都存在適宜的溫度區間。
圖8表示青海地區春季、夏季和秋季Tmax-NDVI的聯合重現期的二維等值線。利用聯合重現期圖可以計算最高溫或者NDVI大于等于特定值對應的重現期。圖中反映了重現期分別為2、5、10、25和50 a時最高溫或者NDVI大于等于特定值的組合情況。聯合重現期越小,最高溫與NDVI的取值范圍越大,說明當最高溫一定時,NDVI出現較小重現期的可能性更大。春季、夏季和秋季Tmax位于較高區間:8~13、16~21和10~16 ℃,NDVI出現較小重現期的概率最大。上述氣溫區間下3個季節的NDVI值分別大于0.1、0.225和0.175,說明該氣溫條件下植物生長良好的情景出現頻次最高。
圖9表示青海春季、夏季和秋季Tmax-NDVI的同現重現期。由同現重現期圖可知最高溫與NDVI同時大于等于特定值的重現期。圖中展現了重現期為2、5、10、25、50 a時最高溫與NDVI同時大于等于特定值的組合情況。對于最高溫和NDVI取值固定的情況,它們的同現重現期明顯大于聯合重現期。對于同現重現期,給定的重現期越大,則最高溫與NDVI的值越大,甚至當最高溫與NDVI的值足夠大時,其同現重現期超過50 a。這說明氣溫超出植物生長的適宜區間后,NDVI的值較大的情景出現頻次較低,植物生長生長發育狀況較大概率比較差。
圖10顯示了青海1982—2016年生長季內春、夏、秋季最低溫和NDVI之間的聯合概率分布關系。從圖中可以獲取任意點的最低溫與NDVI的聯合概率。聯合概率分布函數可以清晰反映出最低溫和NDVI在不同值域范圍上的相關關系。圖中最顯著的特征是數據的不對稱和傾斜的依存結構,并且最低溫取極小值時對NDVI影響較小,取極大值時對NDVI影響顯著。
根據等值線圖可以得出聯合概率為0.1~0.9時最低溫與NDVI的區間分布情況。當最低溫取固定值,NDVI越大,或者當NDVI取值固定,最低溫越大,其聯合概率越大。同一等值線上不同區間的最低溫和NDVI的聯合概率也呈現明顯差異。出現最低溫極小值和NDVI極小值、最低溫極小值和NDVI極大值以及最低溫極大值和NDVI極小值這3種情景的聯合概率較小;而出現最低溫極大值和NDVI極大值的聯合概率較大。當青海3個季節最低溫分別位于-2~4、7~11和1~8 ℃區間時,NDVI的值較高概率相對較大(大于0.6),表明此時植被生長相對良好。
圖11為青海春季、夏季和秋季Tmin-NDVI聯合超越概率的等值線圖。根據等值線圖可以獲取最低溫與NDVI為任意值時的聯合超越概率。圖中顯示了聯合超越概率分別為0.1~0.9時最低溫與NDVI同時大于等于某一特定值的不同組合情況。最低溫與NDVI的取值越小,其聯合超越概率越大;反之亦然。這說明當最低溫與NDVI同時超過某個較小值發生的概率大于同時超過一個較大值對應的概率。青海3個季節在適宜的溫度條件下,植被生長相對良好的情景出現頻次最高。
圖12表示青海春季、夏季和秋季Tmin與NDVI的聯合重現期的二維等值線。從聯合重現期二維圖可知最低溫或者NDVI大于等于特定值的重現期情況,從聯合重現期的等值線圖可以查出給定重現期2、5、10、25和50 a,
最低溫或者NDVI大于等于特定值的各種組合情況。可以看出,聯合重現期越小,最低溫與NDVI的取值范圍越大,這說明當最低溫一定時,NDVI出現較小重現期的可能性更多。當青海省春季、夏季和秋季溫度值分別位于較高時的溫度區間-4~3、7~10和-0.5~6 ℃,較小重現期出現的概率最大,在這些溫度區間下,3個季節植物生長良好的情況出現頻次最高。
圖13表示青海春、夏、秋季最低溫與NDVI的同現重現期。從同現重現期二維圖可知最低溫與NDVI同時大于等于特定值的重現期,從等值線圖可以查出給定重現期2、5、10、25、50 a,最低溫與NDVI同時大于等于特定值的各種組合情況。相同組合下,最低溫與NDVI的同現重現期明顯大于聯合重現期,組合值越大,其同現重現期越成倍大于聯合重現期。對于同現重現期,給定重現期越大,則最低溫與NDVI的值越大,甚至當最低溫與NDVI的值足夠大時,其同現重現期超過50 a。由此可知,在適宜溫度條件下,最低溫能促進植被的生長。隨著最低溫上升,其同現重現期也越大,植被生長良好情景出現頻次越低。因此,最低溫過高時并不一定能促進植物的生長,超過一定閾值時,較大的概率會抑制植物生長。
3 討論
3.1 NDVI變化趨勢分析及原因
近35年來青海地區植被表現出明顯增高趨勢,說明當地生態環境呈現逐漸好轉的態勢,這與青海地區暖濕化的氣候變化趨勢及實施的一系列環境保護政策有關。2000年左右,其NDVI發生明顯躍遷,這可能與20世紀末全球變暖加劇有關(沈永平和王國亞,2013)。21世紀初青海地區啟動了生態治理工程,退耕還林(還草)、禁牧以及控制載畜量,促進草地恢復(劉櫟杉等,2014)。研究表明,青海地區NDVI雖然局部出現退化情況,但整體仍保持穩定趨勢(卓嘎等,2018;韓炳宏等,2019)。青海地區的年降水呈現增加趨勢,氣候在朝暖濕化演變過程中,植被覆蓋對氣溫響應的敏感性高于降水(張戈麗等,2010),可能是由于降水空間分布極不均勻(楊陽等,2020),溫度的升高有利于積雪和冰川融水。溫度通過改變外界環境和影響植物代謝活動作用于植被,氣候變暖改變了高寒植被生態系統的物候期,使得返青期提前(馬曉芳等,2016)。由于氣溫升高,尤其是晝夜溫度的增高深刻地影響了當地凍土地面的熱量條件,季節凍土層熱量條件顯著升高,凍土層深度變薄,凍結時間縮短,這些都直接作用于植被生長。
較低的NDVI反映了青海地區植被覆蓋較差。青海生長季相對短暫,地上生物量普遍較小,其平均值較低。另一方面,青海內部植被類型多樣,復雜的地形和氣候類型狀況使得區域間的植被生長情況具有顯著差異。有些地區由于增溫而改善了植被生長條件,有些地區因為生態環境惡化而導致NDVI降低,在整體區域上的平均值就有可能相互抵消而偏低,NDVI變化趨勢的波動性不大。
3.2 晝夜增溫非對稱性對植被活動影響
大部分植物的光合作用在白天進行,而呼吸作用貫穿全天。晝夜不對稱增溫速率通過作用于植被的生理過程進而影響植被動態變化(Jing et al.,2016;Yang et al.,2017;Wang et al.,2019)。北半球夏季白天增溫不利于溫帶干旱地區植被的生長,但春季白天增溫卻能夠利于寒帶地區植被NDVI的提高(Tan J G et al.,2015)。
青海地區春季、夏季和秋季的夜間最低溫增溫速率分別是白天最高溫增溫速率的0.94倍、1.23倍和1.25倍,季節性晝夜增溫呈現出明顯的不對稱特征。過去50年中,夜間全球表面溫度的升高明顯快于白天,并且Tmin比Tmax高1.4倍(Peng et al.,2013)。青海省夏、秋季變暖趨勢與全球趨勢一致,春季則相反。植物在夜間一般不能進行光合作用,Tmin升高只能引起植物呼吸作用的增強和體內養分代謝速率的增加,造成植物體內有機質損耗增加,表現為NDVI降低;白天植被進行光合作用,理想的水分條件以及Tmax的升高能夠促進植被葉片氣孔更大地張開,使植被在增大蒸騰的同時也增加了CO2進入葉片參與光合作用的概率,因此促進了植被生長,表現為NDVI升高。這表明熱量是影響北半球中高緯度地區植被生長的關鍵因子。
溫帶高寒、山地和荒漠植被對夜間溫度的響應較為積極(Xia et al.,2018)。最高溫和最低溫對植被生長發育都起促進作用,這是由于最高溫的升高有助于促進光合作用合成更多的有機物來滿足植物生長發育的需求。雖然夜間呼吸作用使得有機物凈累積量減少,但是其夜間自養呼吸加劇而產生的補償作用會使其生產力得以提高。青海地區Tmin的增加有助于次日植物體內碳封存的利用,促進植被生長,延長生長季時間,同時夜間增溫降低了霜凍的風險(Wan et al.,2009)。
對于增暖放大現象,存在一種解釋機制可以對晝夜增溫速率的不對稱性進行說明(Zhou et al.,2007)。植被通過覆蓋度來影響地表反照率的變化,進而影響地表能量平衡。在植被稀疏地區,其覆蓋度低,導致反照率降低,減少地表吸收的太陽輻射,由于云量減少而引起的增暖加速在一定程度上被抵消了(Knorr et al.,2001)。同時,植被覆蓋度降低也會導致地表發射率下降。夜間地表長波輻射減少,加速了夜間溫度升高(Zhou et al.,2003)。對于植被覆蓋度較高地區,由于地表反照率的升高導致白天地表氣溫上升加速;同時,地表發射率也表現出增加趨勢,使得夜間地表長波輻射增加,減緩了這類區域的夜間增溫速度。青海地區整體NDVI呈現出由西北向東南增加的趨勢,NDVI的波動性具有明顯的區域差異。其中柴達木盆地周邊及青南牧區西北部屬于植被覆蓋度較低區域,NDVI的波動性大;祁連山東部、青南牧區東南部和東部農業區的植被覆蓋度較高,NDVI波動性較小。因此,對于不同植被覆蓋度以及不同植被類型的區域,其晝夜增溫的不對稱性影響NDVI的程度是有差異的。
3.3 不同季節增溫非一致性對植被活動影響
青海地區NDVI表現為夏季最大,其次為秋季,春季最小。這表明夏季水熱條件最利草地植被的生長,而春季植被生長需要一定溫度的積累,易受倒春寒等天氣的影響使溫度升高較慢。
青海地區春季、夏季和秋季晝夜增溫與NDVI的相關性均呈現為極顯著正相關關系(P<0.01),這是因為熱量資源是植被生長的基礎。最高溫、最低溫和NDVI的相關性強弱均表現為秋季>夏季>春季。夏季的水熱條件均利于植被生長,氣候要素對NDVI影響差異不顯著,而春季溫度相對于秋季波動大,隨著溫度升高蒸發增加,春季的水熱組合并不一定利于植被生長。另外,溫度對NDVI的影響有一定的滯后性(王艷召等,2020),夏季溫度的積累有利于秋季的NDVI,若秋季寒潮強或影響早,會導致植被較早進入枯黃期。
研究發現白天增溫往往能促進植被的光合作用,而更有利于提高植被的NDVI(Peng et al.,2013;Ma et al.,2019b)。本文發現晝夜增溫對青海地區植被的影響程度存在季節性差異。從季節晝夜增溫的影響程度來看,秋季晝夜增溫對青海地區植被的影響程度高于夏季和春季,并且白天增溫和夜間增溫對青海地區NDVI的影響程度也不同,表現為白天增溫影響強于夜間增溫。
青海地區春夏兩季晝夜溫度和NDVI呈現正相關,可能與全球變暖引起的大部分地區春季生長季普遍提前有關(Piao et al.,2007),也可能是白天增溫導致植被展葉期、返青期提前(Piao et al.,2015)。植物通常在日平均溫度上升到5 ℃的時候開始發育,進入夏季青海地區溫度迅速升高,這可促進酶的活性,加快植物發育進程。夏季是青海省水熱條件最好的季節,植被生長旺盛,晝夜增溫對植被生長的影響顯然大于春季。Peng et al.(2013)發現白天溫度升高有利于大部分寒帶、溫帶濕潤地區植被生長及其生態系統碳匯功能;夜晚溫度升高對植被的影響正好與白天相反。Shen et al.(2016)發現,夏季白天增溫利于青藏高原地區的植被生長,而該季節夜間溫度的上升對植被的生長起到消極作用。這可能是由于研究區域和時間的差異導致研究結果不同。或因為夜間增溫雖然會提升植被暗呼吸,增加其體內養分代謝速度,使其體內有機質損耗增加,從而降低植被生產力。與夏季相比,秋季晝夜溫差與NDVI的相關程度更高,這可能是因為夏季Tmax和Tmin在青海地區已接近植被生長的最適溫度,使得NDVI對它們的敏感程度較低。
3.4 利用Copula函數探索氣溫與NDVI動態變化
相關系數作為研究變量間相關性的方法被廣泛采用,但它無法判別變量在不同區間上的動態相關關系,而且無法排除變量之間存在“偽相關”的可能性。這些問題利用Copula函數理論可以得到很好的解決,采用Copula函數可以發現氣溫和NDVI更為細致和真實的動態變化規律。青海地區春季、夏季和秋季晝夜增溫在不同溫度區間條件下與對應的NDVI相依性強弱差異明顯,即對植物生長的影響差異較大。但當它們位于適宜溫度區間時,與NDVI的相依性較強,對植物生長的影響較大。當Tmax或者當Tmin位于適宜的溫度區間時,此時NDVI的較小重現期出現概率最大,說明植物生長良好的情況出現頻次最高。但當Tmax、Tmin與NDVI的值足夠大時,其重現期甚至會超過50 a,說明氣溫超過一定的閾值時,NDVI較高的值出現的概率降低,植物生長受抑制現象出現,氣溫越高,對植物生長抑制作用越大,其顯現良好狀況的概率越低。晝夜增溫以不同的擾動機制增強和抑制植被生長。Tmax、Tmin接近最佳溫度,顯著促進植被光合作用,延長植被生長季周期。當Tmax、Tmin增溫不斷加強,超出植被生長所需最佳溫度,植被生長受到抑制,Tmax、Tmin的持續升高會加快土壤水分蒸發,加重土壤干旱狀況(Melillo et al.,2002;Park et al.,2013),植被無法從土壤中得到生長必需的水分,植被生長對Tmax、Tmin的響應就會轉化為顯性負相關。
青海作為青藏高原的主要部分,是全球氣候變化的敏感區,高原高寒生態系統對氣候變化脆弱而敏感,又會對氣候系統產生反饋,是多圈層耦合過程中的關鍵因子。近30多年來,隨著青藏高原持續變暖,高原生長季植被活動呈持續增強趨勢(Huang et al.,2016;Zou et al.,2020)。增強的植被活動降低了地表生長季白天溫度,對生長季夜間溫度的影響不顯著,總體上降低了局地生長季平均溫度。這種局地降溫效應,主要是由于植被增加導致局地蒸騰作用增強,從而降低了地表能量;青藏高原植被活動對氣候變化形成了“負反饋”。高原植被對氣候的這種“負反饋”作用能夠減緩氣候變暖帶來的負面影響(Liu et al.,2018),但是當氣溫變暖強度超過高原高寒生態系統自身的調節適應能力后,會對當地生態系統造成重大影響。因此,需要密切關注當地的氣候變暖,動態監測NDVI,積極應對全球變暖帶來的挑戰。
由于遙感數據分辨率較低,同一地區植被類型的異質性等問題,難以取得理想結果。在未來研究中需引入更高分辨率的植被分類數據。針對不同植被類型提出不同氣候變化應對政策,因地制宜地建立合適的植被保護措施,將更有利于維護自然生態系統平衡。
4 結論
1)近35年來青海地區生長季內季節性晝夜增溫趨勢顯著,晝夜增溫速率具有不對稱性。春季白天的增溫速率快于夜間,夏、秋季夜間的增溫速率都快于白天。
2)青海地區各季節晝夜增溫對植被的影響在晝夜和季節尺度呈現出明顯非對稱性。晝夜增溫在整體上對青海地區植被影響程度較強,尤其是秋季晝夜增溫對NDVI影響最大,夏季次之,春季最小。白天增溫和夜間增溫對青海地區植被影響程度存在差異,白天增溫的影響大于夜間增溫。
3)青海地區各季節的Tmax和Tmin與NDVI的聯合概率分布函數均呈現一定的規律:當Tmax、Tmin一定,NDVI越大時,或者當NDVI一定,Tmax、Tmin越大時,其聯合概率越大,說明Tmax、Tmin取極小值時對NDVI影響較小,Tmax、Tmin取極大值時對NDVI的影響明顯。
4)重現期越小,晝夜增溫與NDVI的取值范圍越大,這說明當晝夜增溫一定時,NDVI出現較小重現期的可能性更多。晝夜增溫越大,則重現期越大,表明晝夜增溫超過一定的閾值,植物生長受抑制現象出現的概率更大。
參考文獻(References)
Atkin O K,Turnbull M H,Zaragoza-Castells J,et al.,2013.Light inhibition of leaf respiration as soil fertility declines along a post-glacial chronosequence in New Zealand:an analysis using the Kok method[J].Plant Soil,367(1):163-182.doi:10.1007/s11104-013-1686-0.
Barton B T,Schmitz O J,2018.Opposite effects of daytime and nighttime warming on top-down control of plant diversity[J].Ecology,99(1):13-20.doi:10.1002/ecy.2062.
Baumbach L,Siegmund J F,Mittermeier M,et al.,2017.Impacts of temperature extremes on European vegetation during the growing season[J].Biogeosciences,14(21):4891-4903.doi:10.5194/bg-14-4891-2017.
Ben Nasr I,Chebana F,2019.Multivariate L-moment based tests for Copula selection,with hydrometeorological applications[J].J Hydrol,579:124151.doi:10.1016/j.jhydrol.2019.124151.
Chang B,He K N,Li R J,et al.,2018.Trends,abrupt changes,and periodicityof streamflow in Qinghai Province,the northeastern Tibetan Plateau,China[J].Pol J Environ Stud,27(2):545-555.doi:10.15244/pjoes/76037.
Chen C,He B,Guo L L,et al.,2018.Identifying critical climate periods for vegetation growth in the Northern Hemisphere[J].J Geophys Res Biogeosci,123(8):2541-2552.doi:10.1029/2018JG004443.
Chen H P,Sun J Q,2015.Changes in drought characteristics over China using the standardized precipitation evapotranspiration index[J].J Climate,28(13):5430-5447.doi:10.1175/jcli-d-14-00707.1.
Clayton D G,1978.A model for association in bivariate life tables and its application in epidemiological studies of familial tendency in chronic disease incidence[J].Biometrika,65(1):141-151.doi:10.1093/biomet/65.1.141.
Cong N,Shen M G,Yang W,et al.,2017.Varying responses of vegetation activity to climate changes on the Tibetan Plateau grassland[J].Int J Biometeorol,61(8):1433-1444.doi:10.1007/s00484-017-1321-5.
Davy R,Esau I,Chernokulsky A,et al.,2017.Diurnal asymmetry to the observed global warming[J].Int J Climatol,37(1):79-93.doi:10.1002/joc.4688.
Del Grosso S J,Parton W J,Derner J D,et al.,2018.Simple models to predict grassland ecosystem C exchange and actual evapotranspiration using NDVI and environmental variables[J].Agric For Meteorol,249:1-10.doi:10.1016/j.agrformet.2017.11.007.
Deng G R,Zhang H Y,Guo X Y,et al.,2019.Asymmetric effects of daytime and nighttime warming on boreal forest spring phenology[J].Remote Sens,11(14):1651.doi:10.3390/rs11141651.
Du Z Q,Zhao J,Liu X J,et al.,2019.Recent asymmetric warming trends of daytime versus nighttime and their linkages with vegetation greenness in temperate China[J].Environ Sci Pollut Res Int,26(35):35717-35727.doi:10.1007/s11356-019-06440-z.
Easterling D R,Horton B,Jones P D,et al.,1997.Maximum and minimum temperature trends for the globe[J].Science,277(5324):364-367.doi:10.1126/science.277.5324.364.
Genest C,Rivest L P,1993.Statistical inference procedures for bivariate Archimedean Copulas[J].J Am Stat Assoc,88(423):1034-1043.doi:10.1080/01621459.1993.10476372.
Genest C,Favre A C,2007.Everything you always wanted to know about Copula modeling but were afraid to ask[J].J Hydrol Eng,12(4):347-368.doi:10.1061/(asce)1084-0699(2007)12:4(347).
Gong Z N,Zhao S Y,Gu J Z,2017.Correlation analysis between vegetation coverage and climate drought conditions in North China during 2001—2013[J].J Geogr Sci,27(2):143-160.doi:10.1007/s11442-017-1369-5.
Gródek-Szostak Z,Malik G,Kajrunajtys D,et al.,2019.Modeling the dependency between extreme prices of selected agricultural products on the derivatives market using the linkage function[J].Sustainability,11(15):4144.doi:10.3390/su11154144.
韓炳宏,周秉榮,顏玉倩,等,2019.2000—2018年間青藏高原植被覆蓋變化及其與氣候因素的關系分析[J].草地學報,27(6):1651-1658. Han B H,Zhou B R,Yan Y Q,et al.,2019.Analysis of vegetation coverage change and its driving factors over Tibetan Plateau from 2000 to 2008[J].Acta Agrestia Sin,27(6):1651-1658.doi:10.11733/j.issn.1007-0435.2019.06.023.(in Chinese).
He B,Chen A F,Jiang W G,et al.,2017.The response of vegetation growth to shifts in trend of temperature in China[J].J Geogr Sci,27(7):801-816.doi:10.1007/s11442-017-1407-3.
Holben B N,1986.Characteristics of maximum-value composite images from temporal AVHRR data[J].Int J Remote Sens,7(11):1417-1434.doi:10.1080/01431168608948945.
Huang K,Zhang Y J,Zhu J T,et al.,2016.The influences of climate change and human activities on vegetation dynamics in the Qinghai-Tibet Plateau[J].Remote Sens,8(10):876.doi:10.3390/rs8100876.
Li C,Singh V P,Mishra A K,2013.A bivariate mixed distribution with a heavy-tailed component and its application to single-site daily rainfall simulation[J].Water Resour Res,49(2):767-789.
Iglesias A,Garrote L,Quiroga S,et al.,2012.A regional comparison of the effects of climate change on agricultural crops in Europe[J].Climate Change,112(1):29-46.doi:10.1007/s10584-011-0338-8.
Jing P P,Wang D,Zhu C W,et al.,2016.Plant physiological,morphological and yield-related responses to night temperature changes across different species and plant functional types[J].Front Plant Sci,7:1774.doi:10.3389/fpls.2016.01774.
Kalisa W,Igbawua T,Henchiri M,et al.,2019.Assessment of climate impact on vegetation dynamics over East Africa from 1982 to 2015[J].Sci Rep,9(1):16865.doi:10.1038/s41598-019-53150-0.
Karl T R,Knight R W,Gallo K P,et al.,1993.A new perspective on recent global warming:asymmetric trends of daily maximum and minimum temperature[J].Bull Amer Meteor Soc,74(6):1007-1023.doi:10.1175/1520-0477(1993)074<1007:anporg>2.0.co;2.
Knorr W,Schnitzler K G,Govaerts Y,2001.The role of Bright Desert Regions in shaping North African climate[J].Geophys Res Lett,28(18):3489-3492.doi:10.1029/2001gl013283.
Kong D D,Zhang Q,Singh V P,et al.,2017.Seasonal vegetation response to climate change in the Northern Hemisphere (1982—2013)[J].Glob Planet Change,148:1-8.doi:10.1016/j.gloplacha.2016.10.020.
Li C,Singh V P,Mishra A K,2013.A bivariate mixed distribution with a heavy-tailed component and its application to single-site daily rainfall simulation[J].Water Resour Res,49(2):767-789.doi:10.1002/wrcr.20063.
Li H,Liu L,Liu X,et al.,2019.Greening implication inferred from vegetation dynamics interacted with climate change and human activities over the southeast Qinghai-Tibet Plateau[J].Remote Sens,11(20):2421.doi:10.3390/rs11202421.
Liu D,Li Y,Wang T,et al.,2018.Contrasting responses of grassland water and carbon exchanges to climate change between Tibetan Plateau and Inner Mongolia[J].Agric For Meteorol,249:163-175.doi:10.1016/j.agrformet.2017.11.034.
劉櫟杉,延軍平,李雙雙,2014.2000—2009年青海省植被覆蓋時空變化特征[J].水土保持通報,34(1):263-267. Liu L S,Yan J P,Li S S,2014.Spatial-temporal characteristics of vegetation restoration in Qinghai Province from 2000 to 2009[J].Bull Soil Water Conserv,34(1):263-267.(in Chinese).
劉向培,劉烈霜,史小康,等,2015.中國東北植被動態變化及其與氣候因子的關系[J].大氣科學學報,38(2):222-231. Liu X P,Liu L S,Shi X K,et al.,2015.Dynamic variation of vegetation over Northeast China and its relationship with climate factors[J].Trans Atmos Sci,38(2):222-231.doi:10.13878/j.cnki.dqkxxb.20120509001.(in Chinese).
Lu Q,Zhao D S,Wu S H,et al.,2019.Using the NDVI to analyze trends and stability of grassland vegetation cover in Inner Mongolia[J].Theor Appl Climatol,135(3):1629-1640.doi:10.1007/s00704-018-2614-2.
Ma L Q,Qin F,Wang H,et al.,2019a.Asymmetric seasonal daytime and nighttime warming and its effects on vegetation in the Loess Plateau[J].PLoS One,14(6):e0218480.doi:10.1371/journal.pone.0218480.
Ma L Q,Xia H M,Meng Q M,2019b.Spatiotemporal variability of asymmetric daytime and night-time warming and its effects on vegetation in the Yellow River Basin from 1982 to 2015[J].Sensors,19(8):1832.doi:10.3390/s19081832.
馬曉芳,陳思宇,鄧婕,等,2016.青藏高原植被物候監測及其對氣候變化的響應[J].草業學報,25(1):13-21. Ma X F,Chen S Y,Deng J,et al.,2016.Vegetation phenology dynamics and its response to climate change on the Tibetan Plateau[J].Acta Prataculturae Sin,25(1):13-21.doi:10.11686/cyxb2015089.(in Chinese).
Marcelino J,Silva J P,Gameiro J,et al.,2020.Extreme events are more likely to affect the breeding success of lesser kestrels than average climate change[J].Sci Rep,10(1):7207.doi:10.1038/s41598-020-64087-0.
Melillo J M,Steudler P A,Aber J D,et al.,2002.Soil warming and carbon-cycle feedbacks to the climate system[J].Science,298(5601):2173-2176.doi:10.1126/science.1074153.
Meng L,Gao S,Li Y S,et al.,2020.Spatial and temporal characteristics of vegetation NDVI changes and the driving forces in Mongolia during 1982—2015[J].Remote Sens,12(4):603.doi:10.3390/rs12040603.
Miller J A,2012.Species distribution models[J].Prog Phys Geogr Earth Environ,36(5):681-692.doi:10.1177/0309133312442522.
Park T,Chen C,Macias-Fauria M,et al.,2019.Changes in timing of seasonal peak photosynthetic activity in northern ecosystems[J].Glob Chang Biol,25(7):2382-2395.doi:10.1111/gcb.14638.
Park W A,Allen C D,Macalady A K,et al.,2013.Temperature as a potent driver of regional forest drought stress and tree mortality[J].Nat Climate Change,3:292-297.doi:10.1038/nclimate1693.
Peng S S,Piao S L,Ciais P,et al.,2013.Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation[J].Nature,501(7465):88-92.doi:10.1038/nature12434.
Piao S L,Friedlingstein P,Ciais P,et al.,2006.Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades[J].Geophys Res Lett,33(23).doi:10.1029/2006gl028205.
Piao S L,Friedlingstein P,Ciais P,et al.,2007.Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades[J].Glob Biogeochem Cycles,21(3).doi:10.1029/2006gb002888.
Piao S L,Tan J G,Chen A P,et al.,2015.Leaf onset in the Northern Hemisphere triggered by daytime temperature[J].Nat Commun,6:6911.doi:10.1038/ncomms7911.
Reich P B,Sendall K M,Stefanski A,et al.,2018.Effects of climate warming on photosynthesis in boreal tree species depend on soil moisture[J].Nature,562(7726):263-267.doi:10.1038/s41586-018-0582-4.
Rossi S,Isabel N,2017.Bud break responds more strongly to daytime than night-time temperature under asymmetric experimental warming[J].Glob Chang Biol,23(1):446-454.doi:10.1111/gcb.13360.
Sadegh M,Moftakhari H,Gupta H V,et al.,2018.Multihazard scenarios for analysis of compound extreme events[J].Geophys Res Lett,45(11):5470-5480.doi:10.1029/2018gl077317.
Sadegh M,Ragno E,AghaKouchak A,2017.Multivariate Copula Analysis Toolbox (MvCAT):describing dependence and underlying uncertainty using a Bayesian framework[J].Water Resour Res,53(6):5166-5183.doi:10.1002/2016wr020242.
Shen M G,Piao S L,Chen X Q,et al.,2016.Strong impacts of daily minimum temperature on the green-up date and summer greenness of the Tibetan Plateau[J].Glob Chang Biol,22(9):3057-3066.doi:10.1111/gcb.13301.
Shen X J,Liu B H,Henderson M,et al.,2018.Asymmetric effects of daytime and nighttime warming on spring phenology in the temperate grasslands of China[J].Agric For Meteorol,259:240-249.doi:10.1016/j.agrformet.2018.05.006.
沈永平,王國亞,2013.IPCC第一工作組第五次評估報告對全球氣候變化認知的最新科學要點[J].冰川凍土,35(5):1068-1076. Shen Y P,Wang G Y,2013.Key findings and assessment results of IPCC WGI fifth assessment report[J].J Glaciol Geocryol,35(5):1068-1076.doi:10.7522/j.issn.1000-0240.2013.0120.(in Chinese).
Silva P F,de Sousa Lima J R,Antonino A C D,et al.,2017.Seasonal patterns of carbon dioxide,water and energy fluxes over the Caatinga and grassland in the semi-arid region of Brazil[J].J Arid Environ,147:71-82.doi:10.1016/j.jaridenv.2017.09.003.
Singh V P,Zhang L,2018.Copula-entropy theory for multivariate stochastic modeling in water engineering[J].Geosci Lett,5(1):6.doi:10.1186/s40562-018-0105-z.
Singh V,Sivakumar B,Cui H J,2017.Tsallis entropy theory for modeling in water engineering:a review[J].Entropy,19(12):641.doi:10.3390/e19120641.
Sraj M,Bezak N,Brilly M,2015.Bivariate flood frequency analysis using the Copula function:a case study of the Litija Station on the Sava River[J].Hydrol Process,29(2):225-238.doi:10.1002/hyp.10145.
Su H X,Feng J C,Axmacher J C,et al.,2015.Asymmetric warming significantly affects net primary production,but not ecosystem carbon balances of forest and grassland ecosystems in Northern China[J].Sci Rep,5:9115.doi:10.1038/srep09115.
Tan J G,Piao S L,Chen A P,et al.,2015.Seasonally different response of photosynthetic activity to daytime and night-time warming in the Northern Hemisphere[J].Glob Chang Biol,21(1):377-387.doi:10.1111/gcb.12724.
Tan Z Q,Tao H,Jiang J H,et al.,2015.Influences of climate extremes on NDVI (normalized difference vegetation index) in the Poyang Lake basin,China[J].Wetlands,35(6):1033-1042.doi:10.1007/s13157-015-0692-9.
Tang L Y,Duan X F,Kong F J,et al.,2018.Influences of climate change on area variation of Qinghai Lake on Qinghai-Tibetan Plateau since 1980s[J].Sci Rep,8:7331.doi:10.1038/s41598-018-25683-3.
Treharne R,Bjerke J W,Tmmervik H,et al.,2020.Development of new metrics to assess and quantify climatic drivers of extreme event driven Arctic browning[J].Remote Sens Environ,243:111749.doi:10.1016/j.rse.2020.111749.
Wan S Q,Xia J Y,Liu W X,et al.,2009.Photosynthetic overcompensation under nocturnal warming enhances grassland carbon sequestration[J].Ecology,90(10):2700-2710.doi:10.1890/08-2026.1.
Wang Y Q,Luo Y,Shafeeque M,2019.Interpretation of vegetation phenology changes using daytime and night-time temperatures across the Yellow River Basin,China[J].Sci Total Environ,693:133553.doi:10.1016/j.scitotenv.2019.07.359.
Wang Y J,Shen X J,Jiang M,et al.,2020.Vegetation change and its response to climate change between 2000 and 2016 in marshes of the Songnen Plain,Northeast China[J].Sustainability,12(9):3569.doi:10.3390/su12093569.
王艷召,王澤根,王繼燕,等,2020.近20年中國不同季節植被變化及其對氣候的瞬時與滯后響應[J].地理與地理信息科學,36(4):33-40. Wang Y Z,Wang Z G,Wang J Y,et al.,2020.Transient and lagged response of seasonal vegetation changes to climate in China during the past two decades[J].Geogr Geo Inf Sci,36(4):33-40.doi:10.3969/j.issn.1672-0504.2020.04.006.(in Chinese).
Wu X C,Liu H Y,Li X Y,et al.,2016.Seasonal divergence in the interannual responses of Northern Hemisphere vegetation activity to variations in diurnal climate[J].Sci Rep,6:19000.doi:10.1038/srep19000.
Xia H M,Li A N,Feng G,et al.,2018.The effects of asymmetric diurnal warming on vegetation growth of the Tibetan Plateau over the past three decades[J].Sustainability,10(4):1103.doi:10.3390/su10041103.
楊陽,時璐,劉弢,等,2020.青海省西北諸河61a來降水演變特征及空間分布研究[J].中國農村水利水電(8):159-165. Yang Y,Shi L,Liu T,et al.,2020.The evolution characteristics and spatial distribution of precipitation in northwest region of Qinghai in recent 61 years[J].China Rural Water Hydropower(8):159-165.doi:10.3969/j.issn.1007-2284.2020.08.031.(in Chinese).
Yang Z Y,Shen M G,Jia S G,et al.,2017.Asymmetric responses of the end of growing season to daily maximum and minimum temperatures on the Tibetan Plateau[J].J Geophys Res Atmos,122(24):13,278-13,287.doi:10.1002/2017JD027318.
張戈麗,歐陽華,張憲洲,等,2010.基于生態地理分區的青藏高原植被覆被變化及其對氣候變化的響應[J].地理研究,29(11):2004-2016. Zhang G L,Ouyang H,Zhang X Z,et al.,2010.Vegetation change and its responses to climatic variation based on eco-geographical regions of Tibetan Plateau[J].Geogr Res,29(11):2004-2016.(in Chinese).
Zhang H X,Chang J X,Zhang L P,et al.,2018.NDVI dynamic changes and their relationship with meteorological factors and soil moisture[J].Environ Earth Sci,77(16):582.doi:10.1007/s12665-018-7759-x.
Zhang W,Dscher R,Koenigk T,et al.,2020.The interplay of recent vegetation and sea ice dynamics:results from a regional earth system model over the Arctic[J].Geophys Res Lett,47(6):e85982.doi:10.1029/2019gl085982.
Zhang Y L,Wang X,Li C H,et al.,2018.NDVI dynamics under changing meteorological factors in a shallow lake in future metropolitan,semiarid area in North China[J].Sci Rep,8(1):15971.doi:10.1038/s41598-018-33968-w.
Zhou L,Dickinson R E,Tian Y,et al.,2003.A sensitivity study of climate and energy balance simulations with use of satellite-derived emissivity data over Northern Africa and the Arabian Peninsula[J].J Geophys Res,108(D24).doi:10.1029/2003jd004083.
Zhou L M,Dickinson R E,Tian Y H,et al.,2007.Impact of vegetation removal and soil aridation on diurnal temperature range in a semiarid region:application to the Sahel[J].Proc Natl Acad Sci USA,104(46):17937-17942.doi:10.1073/pnas.0700290104.
卓嘎,陳思蓉,周兵,2018.青藏高原植被覆蓋時空變化及其對氣候因子的響應[J].生態學報,38(9):3208-3218. Zhuoga,Chen S R,Zhou B,2018.Spatio-temporal variation of vegetation coverage over the Tibetan Plateau and its responses to climatic factors[J].Acta Ecol Sin,38(9):3208-3218.doi:10.5846/stxb201705270985.(in Chinese).
Zou F L,Li H D,Hu Q W,2020.Responses of vegetation greening and land surface temperature variations to global warming on the Qinghai-Tibetan Plateau,2001—2016[J].Ecol Indic,119:106867.doi:10.1016/j.ecolind.2020.106867.
