神黃豆種子休眠及解除方法研究
2024-06-08 00:21:03楊江敏李富榮余貴湘張恩向董詩凡蔡文良刀保輝
安徽農業科學 2024年10期
楊江敏 李富榮 余貴湘 張恩向 董詩凡 蔡文良 刀保輝

摘要 為探究神黃豆種子的休眠原因及解除方法,對神黃豆種子的生物學特性和萌發抑制物進行觀測,采用熱水、濃硫酸和赤霉素(GA3)浸泡處理,打破種子休眠。結果表明:神黃豆種子為橢圓形,種皮黃褐色,千粒重為378.66 g,含水量為9.97%,種子生活力為100%,發芽潛力較大。完整種子浸泡215 h后,吸水率僅為0.57%,透水性較差。種皮和種胚的甲醇浸提液,對白菜種子的萌發和幼苗根長的生長具有顯著的抑制作用。經98%濃硫酸浸泡10 min,種子發芽率最高(53.33%),用不同溫度熱水浸泡至自然冷卻,萌發促進作用較小,發芽率最高僅為5.56%,GA3浸泡處理種子未萌發。種皮吸水障礙是神黃豆種子休眠的主要原因。4 ℃層積過程中,可溶性糖含量變化較小,25 ℃層積過程中,可溶性糖含量先上升后下降,可溶性蛋白質和粗脂肪含量總體呈下降趨勢,且25 ℃層積過程中的減少速率更快,說明種子解除休眠過程中需要消耗大量營養物質。
關鍵詞 神黃豆;種子休眠;解除休眠
中圖分類號 S722? 文獻標識碼 A
文章編號 0517-6611(2024)10-0133-04
doi:10.3969/j.issn.0517-6611.2024.10.028
Study on Dormancy and Release of Cassia agnes Brenan Seed
YANG Jiang-min,LI Fu-rong,YU Gui-xiang et al
(Dehong Prefecture Forestry Science Research Institute,Ruili,Yunnan 678600)
Abstract In order to explore the dormancy reason and dormancy release method of Cassia agnes seeds,in this research,we mainly studied the biological characteristics and germination inhibitors of C.agnes seeds,hot water,concentrated sulfuric acid and gibberellin (GA3) were used to break the seed dormancy.The results showed that the seed was elliptic,the seed skin was yellowish-brown,smooth and hard,the 1 000-seed weight was 378.66 g,the water content was 9.97%,the seed viability was 100%,and the germination potential was great.After soaking the whole seeds for 215 h,the water absorption rate was only 0.57%,which was significantly different from that of seeds broken skin.The methanol extract from the seed coat and seed embryo of C.agnes had significant inhibitory effects on the germination of Chinese cabbage seeds and the growth of seedling roots.The germination rate was higher (53.33%) when the seeds were soaked in 98% concentrated sulfuric acid for 10 min.Soaking seeds in the water with different temperature until natural cooling had little promoting effect on germination,and the highest germination rate was only 5.56%.The seeds soaked in gibberellin (GA3) can not promote the germination of seeds.The main reasons of C.agnes seeds dormancy were the water absorption barrier.During the stratification at four temperature,the soluble sugar content had little change,in the process of stratification at twenty-five temperature,the soluble sugar content first increased and then decreased,while the soluble protein and crude fat contents showed a decrease trend and the rate of decrease at twenty-five temperature stratification was faster,indicating that a large amount of nutrients were needed in the process of seed dormancy removal.
Key words Cassia agnes Brenan;Seed dormancy;Broken dormancy
基金項目 熱帶珍貴樹種種質資源庫建設項目。
作者簡介 楊江敏(1990—),女,云南芒市人,工程師,碩士,從事林業草原培育和經營研究。*通信作者,工程師,碩士,從事林業草原培育和經營研究。
收稿日期 2023-08-18;修回日期 2023-10-25
神黃豆(Cassia agnes Brenan)為豆科(Leguminosae)決明屬(Cassia)喬木植物[1],產于云南、廣西等省區以及中南半島的山地林中,喜高溫、陽光充足的環境[2]。神黃豆生長快,抗性強,樹形優美,夏季盛開時花團錦簇,具有濃郁的芳香,花期較長,具有較高的觀賞價值,是優良的園林綠化樹種,同時還具有一定的藥理作用[3-6]。神黃豆的合理開發與推廣應用對于豐富城市園林景觀及推進植物藥理研究具有積極意義。神黃豆在云南地區有天然分布,多生于茂密的叢林中,自然條件下種子萌發比較困難,不利于進一步開發利用。目前,國內對神黃豆的研究主要集中在果實、莖葉和莖部所含化合物的分離鑒定、藥理作用分析、引種栽培評價和樹種分類鑒別上,而有關神黃豆種子休眠原因及解除方法的研究鮮見報道。Nikolaeva[7]將種子休眠分為4種類型:種皮的阻礙作用引起休眠,種子成熟時胚未發育完全,種胚自身生理所致,混合因素引起的休眠。機械損傷種皮、熱水、濃硫酸、激素浸泡等方式能有效解除種子休眠[8-11],層積處理能促進種胚發育成熟,改變內源激素含量,增強種子內的呼吸作用等,打破種子休眠[12]。因此,筆者對神黃豆種子休眠原因、解除方法和層積過程中的生理指標變化進行研究,探索打破神黃豆種子休眠方法,以進一步豐富神黃豆植物學研究方面的新資料,為基礎研究提供理論參考。
1 材料與方法
1.1 試驗材料
以2019年10月采自云南省瑞麗市武甸村的神黃豆種子為試驗材料,采種后敲打莢果,取出帶瓤的種子,再剝去種瓤,得出凈種,備用。該試驗所用的種子均為同一家系。
1.2 試驗方法
1.2.1 種子形態特征。
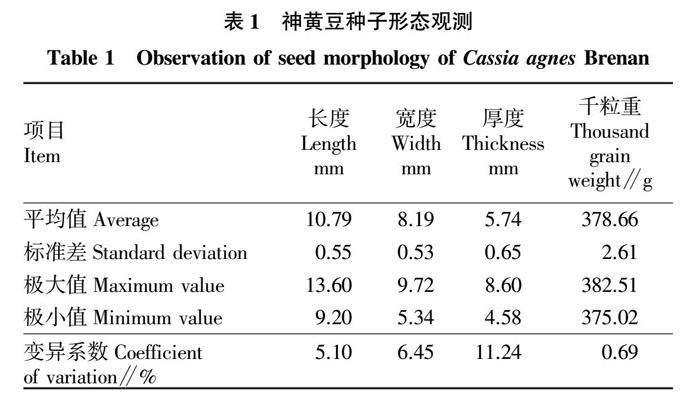
隨機選取100粒種子,觀察其形狀、顏色和光滑度,總結種子形態特征。用游標卡尺測定種子的長度、寬度和厚度(精確到0.01 mm)。采用百粒法測量種子的千粒重[13],測定9組。用SPSS 13.0進行種子的形態變異分析。
1.2.2 種子含水量和吸水率。
隨機選取20粒種子進行稱重,記為W1,烘干至種子重量保持不變,記為W2,重復3次。種子含水量(%)=(W1-W2)/W1×100%。
分別選取完整種子、破皮種子各20粒,稱重,于25 ℃的恒溫水浴箱中浸泡,間隔4 h取出稱重,夜晚8~10 h,直至重量穩定,依次記錄。吸水率=[(濕重-干重)/干重]×100%。
1.2.3 種子生活力。
參考李炎坤等[14]的研究方法,采用氯化三苯基四氮唑(TTC)染色法測定種子生活力。隨機取30粒種子,破皮處理后,用蒸餾水浸泡32 h,使種子充分吸脹,再將種子剖成2瓣,露出種胚,取其中一半放入0.1% TTC溶液中,避光染色12 h,染色結束后立即觀察染色情況,對照種子用蒸餾水煮沸15 min,重復3次。
1.2.4 種子萌發特性。
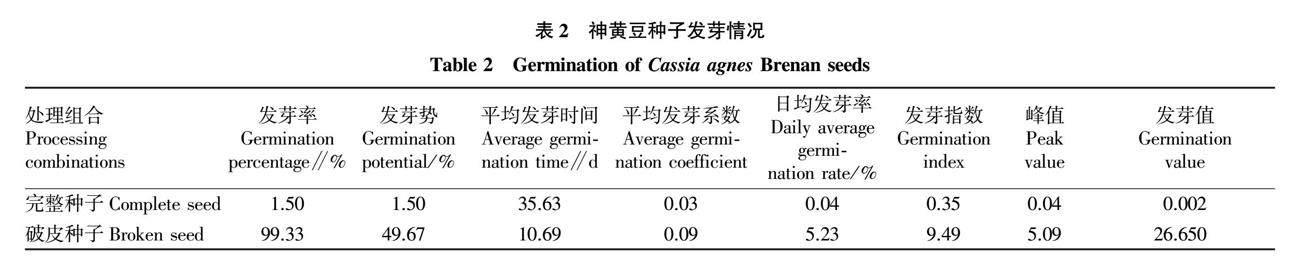
參考閆芳[15]的方法,選取600粒種子,將種子分為2組,每組300粒,其中1組擦破種皮。將完整種子和破皮種子分別置于鋪有濾紙的培養皿中,黑暗條件下于培養箱(溫度25 ℃,濕度10%)中進行萌發試驗。種子發芽以露白為標志[16],萌發期內按時統計發芽率。參考蔡年輝等[17]的方法計算發芽率、發芽勢、日均發芽率、平均發芽時間、平均發芽系數、發芽指數、峰值、發芽值。
1.2.5 種子內源抑制物質的粗提及生物測定。
參考趙敏等[18]的方法,將種子剝離為種皮和胚乳(含胚)2部分,分別制備0.1 g/mL浸提液,選取90粒白菜種子(云南祺合農業科技有限公司,醉貴妃Q-6,純度95.0%,凈度98.0%,發芽率85%以上),分為3組,每組30粒,均勻放置在培養皿中,分別加入4 mL不同濃度(50%、75%、100%)的浸取液,重復3次,在恒溫培養箱(18 ℃)中進行萌發,以蒸餾水處理為對照,24 h后統計發芽率(當胚根長度達到種子長度的1/2時,記為發芽),72 h后測量白菜幼苗的莖長和根長。
1.2.6 不同處理方法對神黃豆種子萌發的影響。
參考前人研究方法[19-21],分別用3種不同溶液對種子進行處理,熱水(100、90、80 ℃)浸泡至自然冷卻;濃硫酸溶液(98%、80%、70%)浸泡10 min,期間用玻璃棒持續攪拌,處理后在流水下沖洗種子1 h;GA3溶液(700、500、300mg/L)避光浸泡72 h,每個處理90粒種子,每組30粒,重復3次,置于25 ℃恒溫箱中進行萌發試驗。以上試驗以清水處理為對照,胚根突破種皮露白作為發芽標志,每天記錄發芽情況。
1.2.7 不同溫度層積處理對生理指標的影響。
取一定量的神黃豆種子放入50%多菌靈可濕性粉劑800倍液浸泡4 h,用清水沖洗后,按照3∶1的比例將種子與沙子混勻(含水量保持在50%左右),分別放入4和25 ℃的恒溫箱中進行層積處理,每30 d取樣1次,分別測定種子可溶性糖、可溶性蛋白質和脂肪的含量變化。可溶性糖含量采用蒽酮比色法測定,可溶性蛋白質含量采用考馬斯亮藍法測定,粗脂肪含量使用索氏提取法測定[22]。
1.2.8 數據分析。應用Microsoft Excel 2003進行試驗數據處理,方差分析選用SPSS 13.0,采用Tukey法進行多重比較。
2 結果分析
2.1 種子生物學特性
2.1.1 種子形態特征。
神黃豆種子為橢圓形,種皮黃褐色,光滑堅硬,長度為(10.79±0.55)mm,寬度為(8.19±0.53)mm,厚度為(5.74±0.65) mm,種子厚度的變異系數最大,達11.24%,長度和寬度相對穩定,千粒重為(378.66±2.61)g,9組數據的變異系數為0.69%,無明顯的變異趨勢,顆粒重量較平均(表1)。
2.1.2 種子含水量。經測定,
神黃豆種子的平均干種子重7.782 g,平均烘干重7.005 g,平均含水量為9.97%,含水量變化較小。
2.1.3 種子生活力。經TTC染色法測定,參試的神黃豆種子生活力100%(圖1),發芽潛力高。
2.1.4 種子萌發特性。
破皮處理和完整的神黃豆種子發芽率、發芽勢、平均發芽系數、日均發芽率、發芽指數、峰值和發芽值差異極顯著(P<0.01),平均發芽時間差異不顯著(P>0.05)。破皮處理種子的平均發芽率、發芽勢、發芽系數、日均發芽率、發芽指數、峰值和發芽值在發芽期內均大于完整種子,說明破皮處理能有效促進種子萌發(表2)。
2.2 神黃豆種子休眠原因
2.2.1 種子吸水率。破皮處理和完整種子吸水率差異極顯著(P<0.01)。破皮種子吸水率隨浸種時間的延長呈拋物線型變化,在0~48 h為快速吸水期,該階段吸水率達到116.7%,占吸水總量的75.8%,浸種167 h后,吸水量逐漸趨于飽和;完整種子浸泡215 h后,吸水率僅為0.57%,說明種子存在吸水障礙(圖2)。
2.2.2 種皮、種胚甲醇浸提液對白菜種子萌發和幼苗生長的影響。
由表3可知,種皮和種胚的甲醇浸提液對白菜種子的發芽率和幼苗根長的生長具有顯著的抑制作用(P<0.05),且隨著浸提液濃度的升高抑制作用逐漸增強。種胚甲醇浸提液對白菜種子的萌發抑制作用高于種皮,浸提液濃度為0.100 g/mL時,白菜種子的萌發率分別為15.56%和10.00%,相比CK分別下降了82.27百分點和88.61百分點,白菜幼苗的根長為3.27和5.46 mm,相比CK分別縮短了11.35和9.16 mm;種皮和種胚甲醇浸提液對白菜幼苗莖長的抑制作用不明顯。
2.3 種子休眠解除方法
經100、90、80 ℃熱水浸泡至自然冷卻,各處理間差異不顯著(P>0.05),100 ℃熱水處理的種子發芽率、發芽勢、發芽指數為5.56%、3.33%、0.23,萌發促進作用較小;98%、80%、70%濃硫酸處理10 min,期間用玻璃棒不斷攪拌,98%濃硫酸處理與80%濃硫酸處理間發芽勢的差異達到顯著水平(P<0.05),98%濃硫酸、80%濃硫酸處理與70%濃硫酸處理間的發芽率和發芽指數差異顯著(P<0.05),98%濃硫酸處理的種子發芽率、發芽勢、發芽指數最高,分別達到53.33%、36.66%、2.37,總體發芽情況較好;經GA3浸泡的種子觀測期內均未萌發(表4)。
2.4 不同溫度層積過程中生理指標的變化
2.4.1 可溶性糖含量的變化。
從圖3可知,不同溫度層積處理可溶性糖含量變化差異極顯著(P<0.01)。25 ℃層積過程中,0~30 d可溶性糖含量快速增加,30~60 d可溶性糖含量緩慢增加,在60 d時達到最大值86.7 mg/g,60~90 d可溶性糖含量迅速下降,120 d時下降至最小值3.3 mg/g。前期由于淀粉和脂肪的降解轉化,可溶性糖含量逐漸增加,第60天開始,部分種子開始萌發,種子呼吸作用增強,可溶性糖被胚利用或被種子呼吸作用消耗而降低。4 ℃層積過程中可溶性糖含量變化較小。
2.4.2 可溶性蛋白質含量的變化。
從圖4可知,不同溫度層積處理蛋白質含量變化差異極顯著(P<0.01),層積過程中,可溶性蛋白質含量總體呈下降趨勢,且25 ℃層積過程中下降速率較快,可溶性蛋白質含量持續減少,說明可溶性蛋白質被水解利用,為種子萌發提供能量。
2.4.3 粗脂肪含量的變化。
由圖5可知,不同溫度層積處理粗脂肪含量變化差異極顯著(P<0.01)。
4 ℃層積過程中,0~30 d粗脂肪含量下降較快,與層積前相比下降約42.2%;25 ℃層積過程中,0~30 d粗脂肪含量與層積前相比下降約67.7%。粗脂肪含量前期快速下降,后期變化幅度較小,且25 ℃層積過程中的下降速率大于4 ℃層積處理,說明粗脂肪不斷分解轉化,為種子萌發提供能量。
3 討論與結論
種子萌發的第一步是吸水[23]。曹運梅等[9]報道了銅錢樹種子果殼和種皮存在機械障礙,未經處理種子較難萌發。神黃豆完整種子浸泡215 h,吸水率僅為0.57%,說明種皮結構致密,透水性差,阻礙了種子的吸水。種子內源抑制物也是引起休眠的主要原因之一[24-25]。劉洋等[26]報道了烏蘇里鼠李種子內部物質對種子萌發存在抑制作用。筆者在研究中發現,神黃豆種皮和種胚的甲醇浸提液對白菜種子的萌發和幼苗根長的生長具有顯著的抑制作用,且隨著濃度的升高抑制作用逐漸增強,說明神黃豆種皮和種胚均存在抑制白菜種子萌發的物質,至于該物質成分及其對神黃豆自身萌發是否具有抑制作用有待于后續研究。王付娟等[27-28]研究表明,熱水和濃硫酸浸泡能夠有效促進野生大豆、美麗決明和絨果決明種子萌發。神黃豆種子經不同溫度熱水浸泡至自然冷卻,最高發芽率僅為5.56%;98%濃硫酸和80%濃硫酸浸泡處理種子10 min能有效提高種子萌發率;經不同濃度GA3溶液浸泡處理后,種子均未萌發,可能是種皮阻礙了激素的作用。賴小連等[29]研究表明,刻傷夏蠟梅的種皮能夠有效提高發芽率。萌發試驗中,擦破神黃豆種皮,種子萌發率達到99.33%,說明種皮的不透水性阻礙了種子萌發。淀粉、蛋白質、脂肪等種內貯藏物質在相應酶的作用下分解并轉化助力種子萌發,為種胚及幼苗的成長提供必要的營養物質與能量[30]。層積過程中,可溶性糖、可溶性蛋白質和粗脂肪含量總體呈下降趨勢,為種子萌發提供了能量來源。
綜上所述,擦破種皮和98%濃硫酸、80%濃硫酸浸泡處理10 min,能夠有效提高神黃豆種子的發芽率。種皮堅硬、透性差是導致神黃豆種子休眠的主要原因,種皮和種胚甲醇浸提液對白菜種子萌發和幼苗生長影響試驗中,雖然存在一定的抑制作用,但對神黃豆種子休眠的影響還需進一步確定。可溶性糖、可溶性蛋白質和粗脂肪不斷分解轉化,表明貯藏物質參與并調節了神黃豆種子休眠解除的過程,但不同溫度層積處理后種子發芽率與各生理指標的內在聯系還需進一步試驗和研究。
參考文獻
[1] 《中國高等植物彩色圖鑒》編委會等.中國高等植物彩色圖鑒:第4卷[M].北京:科學出版社,2015:306-307.
[2] 胡勇,陳麗瓊,朱霞,等.決明屬化學成分與藥理作用研究進展[J].現代中藥研究與實踐,2013,27(5):69-71.
[3] 張淑紅.神黃豆[J].植物雜志,2003(6):28.
[4] 劉興劍,湯詩杰,竇劍,等.幾種熱帶蘇木科觀賞植物的引種觀察[J].江蘇農業科學,2009,37(5):193-195.
[5] 王閃閃,黃文忠,曾廣智,等.神黃豆莖葉中酚性成分研究[J].云南民族大學學報(自然科學版),2019,28(2):105-108.
[6] 成功,龔濟達,薛達元,等.云南省隴川縣景頗族藥用植物傳統知識現狀[J].云南農業大學學報,2013,28(1):1-8.
[7] NIKOLAEVA M G.Physiology of deep dormancy in seeds (Fiziologiya glubokogo pokoya semyan)[M].Jerusalem:Israel Program for Scientific Translations,1969.
[8] BASKIN J M,BASKIN C C.A classification system for seed dormancy[J].Seed science research,2004,14(1):1-16.
[9] 曹運梅,張建,向云,等.銅錢樹種子休眠特性及破眠技術研究[J].種子,2019,38(8):73-76.
[10] 李兵兵,魏小紅,徐嚴.麻花秦艽種子休眠機理及其破除方法[J].生態學報,2013,33(15):4631-4638.
[11] 程廣有,唐曉杰,高紅兵,等.東北紅豆杉種子休眠機理與解除技術探討[J].北京林業大學學報,2004,26(1):5-9.
[12] 孫躍春,樊奮成,張英俊.預冷打破種子休眠的研究進展[J].種子,2004,23(10):52-53.
[13] 國家技術監督局.農作物種子檢驗規程 其他項目檢驗:GB/T 3543.7—1995[S].北京:中國標準出版社,1996:11-12.
[14] 李炎坤,卓一南,曾湘達,等.崗梅種子生活力測定方法的優化[J].種子,2018,37(11):128-131.
[15] 閆芳.黃瑞香種子休眠機理研究[D].蘭州:甘肅農業大學,2016.
[16] 魏琳,朱仁斌,程積民,等.不同處理對本氏針茅種子萌發影響的研究[J].種子,2012,31(7):25-27.
[17] 蔡年輝,許玉蘭,鄧麗麗,等.莖干通直與扭曲云南松種子萌發特性比較[J].西南林業大學學報,2017,37(5):9-14.
[18] 趙敏,王炎.膜莢黃芪種子萌發抑制物質特性的初步研究[J].中草藥,2001,32(7):643-646.
[19] 武承云.不同處理方法對甘肅鳶尾種子萌發的影響[J].青海農林科技,2021(1):93-95.
[20] 羅小燕,李欣勇,孫英男,等.不同處理對山螞蝗種子萌發和幼苗生長的影響[J].熱帶農業科學,2017,37(11):16-21.
[21] 趙冰,董進英,張冬林.溫度、光照和赤霉素濃度對秀雅杜鵑種子萌發的影響[J].種子,2014,33(5):26-30.
[22] 李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000:207-208.
[23] MURDOCH A J.Seeds:Ecology,biogeography,and evolution of dormancy and germination[J].Crop science,2000,40(2):564-565.
[24] 司倩倩,臧德奎,劉丹,等.玉鈴花種子休眠原因及其內源激素含量變化[J].北方園藝,2017(6):91-95.
[25] 付楠,宋慧,王淑君,等.種子的休眠與破除研究進展[J].安徽農業科學,2018,46(24):10-12,15.
[26] 劉洋,郭嬌,王娟,等.烏蘇里鼠李種子休眠機理及萌發特性研究[J].西北農林科技大學學報(自然科學版),2020,48(12):127-135.
[27] 王付娟,劉書含,李淑梅,等.野生大豆種皮障礙休眠解除及萌發的研究[J].大豆科學,2019,38(5):733-739.
[28] 陳瑩,孫苗苗,鐘麗媚,等.不同浸種處理對美麗決明和絨果決明種子萌發的效果研究[J].種子,2019,38(5):70-73.
[29] 賴小連,顏立紅,顏玉娟,等.夏蠟梅種子形態結構和種子萌發研究[J].種子,2021,40(2):86-90.
[30] 廖云嬌,李雪,董學會.不同變溫層積過程中東北紅豆杉種子生理生化特性和胚形態的變化[J].中國農業大學學報,2010,15(1):39-44.
