瓊凹大葉蟬精子發生及精子超微結構
2024-06-08 17:41:17陳實銳陳哲晶鄭欣然李佳靈張貝貝
植物保護 2024年3期
關鍵詞:結構
陳實銳 陳哲晶 鄭欣然 李佳靈 張貝貝
摘要
昆蟲精子超微結構在不同類群間變異較大,在昆蟲分類、種群進化研究及系統發育分析中具有重要意義。研究葉蟬科昆蟲精子發生及精子超微結構,可為葉蟬科及頭喙亞目系統發育分析提供更多特征依據。本研究采用光學和電子顯微鏡技術,觀察了瓊凹大葉蟬Bothrogonia?qiongana?Yang?et?Li的精子發生和精子超微結構。精子形成時期,精子細胞內高爾基復合體形成精子頂體;線粒體融合衍變為線粒體衍生物;中心粒產生軸絲。單層微管結構圍繞精子各組成部分,在精子成熟后消失,對精子細胞伸長起關鍵作用。成熟精子細長、線狀,頭部頂端由繩狀物質連接形成梳狀精子束。頂體圓錐狀,具有頂體腔,末端延伸出2條突起插入細胞核頂端。細胞核近柱狀,末端平截,側緣凹陷,通過片層狀中心粒側體與鞭毛線粒體衍生物連接。尾部鞭毛由1條9+9+2型軸絲,2條對稱線粒體衍生物,2條較短副體,2條細小附結構及鞭毛中心物質組成。瓊凹大葉蟬副體橫切橢圓形、長度較短僅出現在鞭毛起始區,與角頂葉蟬亞科昆蟲差異較大。結合已報道頭喙亞目類昆蟲,我們推斷副體結構可為葉蟬科及頭喙亞目系統發育分析提供有用的特征。
關鍵詞
頭喙亞目;?大葉蟬亞科;?生殖細胞;?精子形成;?超微結構;?副體
中圖分類號:
Q?964
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2023271
Spermatogenesis?and?sperm?ultrastructure?of?Bothrogonia?qiongana?Yang?et?Li?(Hemiptera:?Cicadellidae)
CHEN?Shirui1,?CHEN?Zhejing1,?ZHENG?Xinran1,?LI?Jialing2,?ZHANG?Beibei1*
(1.?College?of?Forestry,?Hainan?University,?Danzhou?571737,?China;?2.?Wuzhishan?Branch?of?the?Management
Office?of?Hainan?Tropical?Rainforest?National?Park,?Wuzhishan?572200,?China)
Abstract
Sperm?ultrastructure?displays?a?great?variability?among?insect?groups?and?holds?great?significance?in?the?classification,?evolution,?and?phylogenetic?analysis?of?insects.?The?study?of?spermatogenesis?and?sperm?ultrastructure?in?Cicadellidae?can?provide?additional?valuable?characters?for?phylogenetic?analysis?of?both?Cicadellidae?and?Auchenorrhyncha.?Spermatogenesis?and?sperm?ultrastructure?of?the?leafhopper?Bothrogonia?qiongana?Yang?et?Li?were?investigated?using?light?and?electron?microscopy.?During?spermiogenesis,?the?acrosome?originated?from?the?Golgi?apparatus,?while?the?mitochondrial?derivatives?formed?through?the?aggregation?and?fusion?of?numerous?mitochondria?in?the?spermatid?cytoplasm.?The?flagellar?axoneme?was?generated?by?the?centriole.?Spermatid?components?were?surrounded?by?a?single?layer?of?microtubules,?which?played?a?critical?role?in?the?spermatid?elongation?during?spermiogenesis,?which?then?disappeared?in?mature?spermatozoa.?The?spermatozoa?exhibited?an?elongated,?filiform?structure.?The?sperm?heads?were?arranged?in?a?row?with?a?ropelike?structure,?forming?a?pectinate?sperm?bundle.?The?conical?acrosome?possessed?a?subacrosomal?space?and?formed?two?acrosomal?processes?inserting?into?both?sides?of?the?anterior?part?of?nucleus.?The?nucleus?was?cylindrical,?with?a?flattened?base,?and?its?marginal?depression?connected?to?the?two?mitochondrial?derivatives?of?the?flagellum?through?a?lamellate?centriolar?adjunct.?The?flagellum?was?composed?of?a?9+9+2?axoneme,?two?symmetrical?mitochondrial?derivatives,?two?short?accessory?bodies,?two?small?subellipsoidal?structures,?and?a?centerflagellar?material.?Two?ovoid?and?short?accessory?bodies?observed?only?in?the?flagellums?initial?region?of?Bothrogonia?qiongana,?were?quite?different?from?those?of?species?in?Deltocephalinae.?Combined?with?the?reported?data?from?other?auchenorrhynchans,?these?accessory?bodies?likely?provide?valuable?characters?for?phylogenetic?analysis?of?Cicadellidae?and?Auchenorrhyncha.
Key?words
Auchenorrhyncha;?Cicadellinae;?germ?cell;?spermiogenesis;?ultrastructure;?accessory?body
葉蟬科Cicadellidae是昆蟲綱Insecta半翅目Hemiptera頭喙亞目Auchenorrhyncha中最大的類群,目前已知記載約1?500屬20?000余種[1],廣泛分布于世界各地,是生態系統中昆蟲物種最豐富的類群之一。該類昆蟲具有典型的刺吸式口器和較強的生殖力,其通過吸食植物汁液影響植物生長,一些種類還可以傳播植物病毒,是重要的農林業害蟲。然而,葉蟬科的系統發育關系至今仍存在較大爭議[214]。近年來,基于形態學和分子數據的系統發育研究認為葉蟬科是并系群,科內窄頰葉蟬亞科Ulopinae和圓痕葉蟬亞科Megophthalminae互為姊妹群,與角蟬分支(犁胸蟬科Melizoderidae、美角蟬科Melizoderidae和角蟬科Membracidae)聚成一支[5,8,14],而葉蟬科其他亞科支系間親緣關系仍處于爭論中。因此,若要完善葉蟬科的系統發育分析,還需尋找更多的特征依據。
精子是有性生殖動物中高度特化且形態最多樣化的一類特殊細胞[15],對種族的繁衍、延續至關重要。昆蟲精子因快速、趨異的進化特性,形態結構復雜多樣,尤其是精子的超微結構在不同類群間表現出顯著差異,可為昆蟲分類、進化及系統發育分析提供有效的特征依據[1622]。然而,關于葉蟬科昆蟲的精子超微結構研究迄今僅涉及少數種類,其中精子超微結構被完整研究的種類只有6種,包括玉米黃翅葉蟬Dalbulus?maidis?(Delong?&?Wolcott)、條沙葉蟬Psammotettix?striatus?(Linnaeus)、橫線頂帶葉蟬Exitianus?nanus?(Distant)、短二室葉蟬Balclutha?brevis?Lindberg、大青葉蟬Cicadella?viridis?(Linnaeus)和白邊大葉蟬Kolla?paulula?(Walker)[2328]。因此,亟須開展更多葉蟬科昆蟲的精子研究。
瓊凹大葉蟬Bothrogonia?qiongana?Yang?et?Li,?1980,隸屬于葉蟬科大葉蟬亞科Cicadellinae凹大葉蟬屬Bothrogonia,已知分布于中國華中、華南等地山間陰濕林區,尤其在海南地區廣泛分布,為害林木及近林區農作物,是一類重要潛在農林業害蟲[29]。本研究采用光學和電子顯微鏡技術,對瓊凹大葉蟬雄性生殖器官、精子發生及精子超微結構進行觀察研究,旨在為葉蟬科及頭喙亞目系統發育分析提供更多有用的特征依據。
1?材料與方法
1.1?供試標本
瓊凹大葉蟬雄成蟲于2020年,2021年的6月-8月在海南省五指山市海南熱帶雨林國家公園五指山片區(18°54′N,?109°40′E)掃網捕獲。網捕采集的成蟲裝入預先盛放有新鮮枝條的養蟲籠內,帶回實驗室備用。
1.2?光學顯微鏡樣品制備與觀察
用乙酸乙酯將新鮮雄性活體成蟲麻醉,放入盛有磷酸緩沖液(0.1?mol/L?PBS,?pH?7.2)的凹面皿中,在Olympus?SZX7體視顯微鏡下迅速解剖,獲得完整的雄性生殖系統,并置于裝有HQimagie?C630成像系統的Nikon?SMZ800N光學體視顯微鏡下觀察、拍照。
精子形態觀察:將快速解剖分離出來的貯精囊轉移至預先滴有磷酸緩沖液的潔凈載玻片上,破壁使囊內精子游離出來,加蓋蓋玻片后置于連有Nikon?DSFi3成像系統的Nikon?NI/E顯微鏡下觀察、拍照。
精子細胞核形態觀察:將貯精囊置于預先滴有蒸餾水的潔凈載玻片上,破壁后使精子游離出來,室溫條件下晾干后用4,6二脒基2苯基吲哚(DAPI,?Beyotime)對精子進行染色,蒸餾水沖洗干凈后在顯微鏡下觀察、拍照。
在同一顯微鏡下,隨機拍攝50個精子和精子細胞核,采用NISElements?(D?5.20.00)軟件測量長度,并結合Excel?2010軟件及SPSS進行精子長度統計、分析。
1.3?透射電子顯微鏡樣品制備與觀察
樣品預處理:用乙酸乙酯將新鮮雄性活體成蟲麻醉,在低溫環境下將其置于盛有2%多聚甲醛-2.5%戊二醛固定液的表面皿中,在體視顯微鏡下快速解剖。將精巢小管和貯精囊剪切為1?mm3的樣塊,投入盛有添加3%蔗糖溶液的多聚甲醛戊二醛固定液的離心管中,4℃條件下避光固定12?h以上。
透射電子顯微鏡樣品制備與觀察:預處理后的樣品室溫條件下用磷酸緩沖液漂洗,4℃條件下置于1%鋨酸中固定2?h,經磷酸緩沖液漂洗后用乙醇溶液(30%、50%、75%、80%、90%、100%)逐級脫水。脫水后的樣品依次經過不同濃度的乙醇LR?White樹脂混合物(乙醇和LR?White樹脂體積比為3∶1,1∶1和1∶3)及純LR?White樹脂滲透后包埋,并置于烘箱內于60℃條件下聚合48?h。聚合好的樣品膠粒使用Leica?EM?UC7超薄切片機進行定位及切片,切片厚度約70?nm,經醋酸雙氧鈾和檸檬酸鉛雙重染色后,在Hitachi?HT7700透射電子顯微鏡80?kV下觀察并拍照。
1.4?組織學觀察
聚合好的樣品膠粒經修整后,用裝有玻璃刀的切片機切成厚度約800?nm的半薄切片。將樣品切片置于預先滴有蒸餾水的潔凈載玻片水滴上,烤干后經0.5%甲苯胺藍染色,蒸餾水沖洗,再次烤干,最后在顯微鏡下觀察、拍照。
2?結果與分析
2.1?瓊凹大葉蟬雄性生殖器官
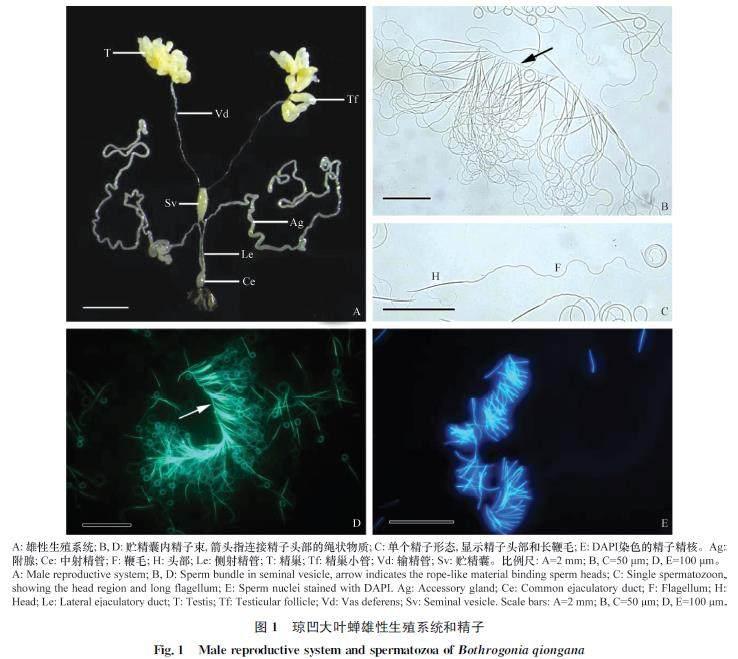
瓊凹大葉蟬雄性生殖系統由精巢、輸精管、貯精囊、附腺、射精管及外生殖器組成(圖1A)。其中1對精巢左右對稱,各由12個長度約為1.27?mm的淡黃色指狀精巢小管組成,基部連接輸精管。2條輸精管細長,前端略粗,后端較細,末端膨大形成長卵形、長寬比約為5∶1的淡黃色貯精囊,并與1對發達的附腺連接側射精管。2條側射精管末端匯合形成膨大、近球狀的中射精管,最后與外生殖器連接(圖1A)。
2.2?瓊凹大葉蟬精子發生
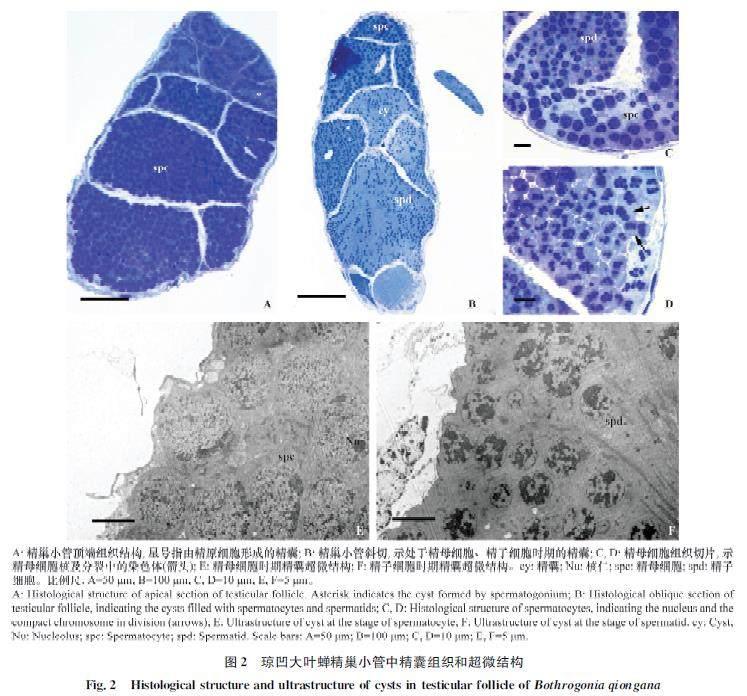
瓊凹大葉蟬精子發生在精巢小管中以精囊為單位同步進行細胞分裂、形變(圖2A~D)。精巢小管頂端精原細胞被非生殖性細胞包圍形成精囊(圖2A,星號),精原細胞在精囊中進行多次有絲分裂產生精母細胞(圖2A,?2E)。精母細胞經歷2次分裂產生精子細胞(圖2C,?2D,?2F),而精子細胞經過形變最終形成線狀成熟精子(圖1C,?2F)。精子發生早期,隨著精原細胞的產生、分裂及增殖,精巢小管內精囊逐漸增多,體積增大(圖2A~B),向基部移動。精原細胞經過多次連續有絲分裂后產生精母細胞,而處于精母細胞時期的精囊內細胞體積較大,具有直徑約8.3?mm的細胞核,且在核周細胞質內有大量為細胞分裂提供能量的線粒體(圖2C,?2E,?3A)。精子發生中期,精囊內精母細胞經歷2次分裂產生精子細胞(圖2B~D)。精子細胞相比于精母細胞,細胞核明顯較小,直徑約4.3?mm(圖2C,?2F)。
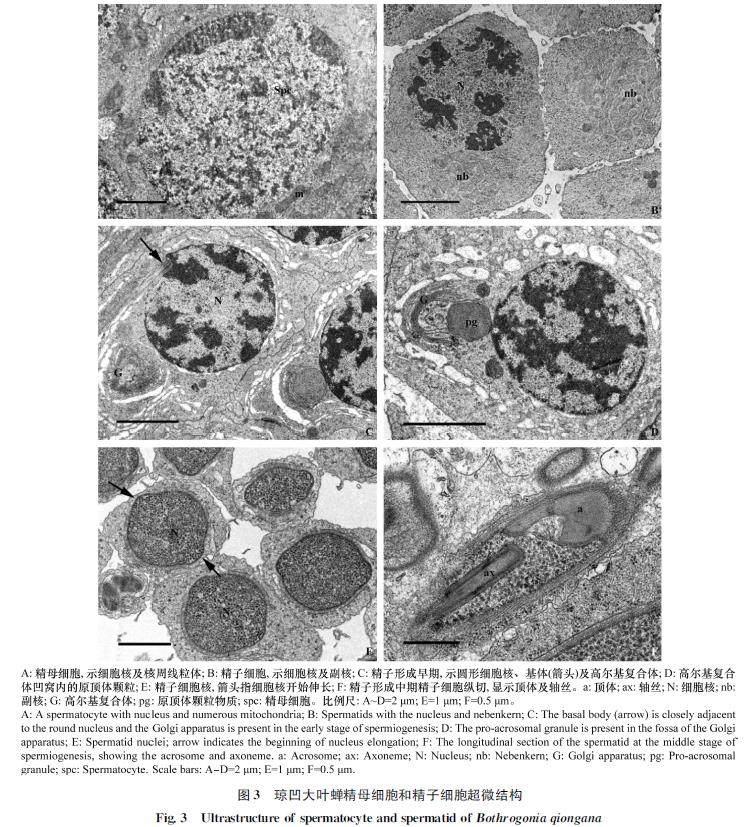
精子發生后期是精子細胞形變轉化為線狀精子的過程,又稱為精子形成時期。精子形成初期,精子細胞具有染色質較均勻的圓球狀細胞核,而進入精子形成早期細胞核內染色質開始小區域凝集,細胞質內線粒體聚集融合形成連鎖狀副核結構(圖3B)。隨著時間的延續,細胞質內出現明顯的高爾基復合體及原頂體結構,同時細胞核基部出現由中心粒形變形成的基體結構(圖3C)。原頂體顆粒出現在高爾基復合體凹窩內,體積逐漸增大,并隨高爾基復合體開始移向細胞核頂端位置(圖3C,?3D)。細胞核基部基體產生軸絲結構。隨著軸絲的出現,細胞質中副核開始移向軸絲兩側。精子形成中期,精子細胞細胞核由球狀開始向兩端伸長,核內的染色質繼續凝集(圖3E)。而細胞質內的原頂體顆粒與高爾基復合體一起移動至細胞核頂端,形成內部具腔的頂體結構(圖3F)。精子細胞核基部副核形成2條大小相等的線粒體衍生物,平行于軸絲結構(圖4C)。頂體、細胞核及線粒體衍生物等結構隨精子形成過程逐漸伸長(圖4A,?4D),外圍具有一圈明顯的單層微管結構(圖4B,?4C)。精子形成后期,細胞核基部中心粒周圍物質逐漸在細胞核與線粒體衍生物之間形成中心粒側體結構(圖4D)。精子細胞內組成精子的各部分結構繼續伸長,同時細胞核內染色質進一步凝集。精子形成末期,精子細胞的細胞核伸長呈圓柱狀,核內染色質變得致密(圖4E,?4F),核外微管層逐步消失,而精子細胞質膜內仍具有部分細胞質殘余物質(圖4F,星號)。隨著時間的推移,精子各組成部分完成伸長過程,其中精子頭部頂體呈帽狀結構插入細胞核頂端,頸區中心粒側體沿鞭毛延伸成副體,鞭毛軸絲和兩條線粒體衍生物隨鞭毛延伸至末端。精子形變成熟后,與形成末期精子細胞區別的典型特征為精子質膜內細胞質殘余物質消失不見,呈現透明狀(圖6B,星號)。
2.3?瓊凹大葉蟬精子超微結構
貯精囊內成熟精子長線狀,約(196.41±1.67)?μm(樣本量50,平均值±標準誤),主要由較短的頭部和細長的鞭毛組成(圖1C),其中精子頭部細胞核長約(39.01±0.22)?μm(圖1E),頭、尾長度比約為1∶4。瓊凹大葉蟬精子整體形態無多態性現象,貯精囊內成熟精子頭部頂端排列整齊由繩狀物質連成一排,形成梳狀精子束(圖1B,?1D)。
精子頭部主要由致密的長細胞核和位于細胞核頂端的頂體組成(圖5A,?5B)。頂體圓錐狀,長度較短,約3?μm,基部直徑約0.4?μm,由纖維管狀結構組成(圖5A,?5B)。頂體頂端通常插入胞外基質中,后端多呈帽狀插入精核頂端,頂體內形成頂體腔(圖5A,?B)。頂體腔直徑漸增,前端通常不與頂體同軸心,后端多不規則。頂體末端兩側多延伸成突起插入細胞核頂端,橫切圖上清晰可見(圖5A)。細胞核細長,圓柱狀(圖1C,?1E,?5C),由致密染色質組成。精核直徑自頂端處至基部漸增,0.4~0.6?μm,其橫切由前端橢圓形至特殊的不規則五邊形(圖5A,?6A,?6B)。
頭部細胞核與尾部鞭毛的連接區為精子頸區,主要由中心粒和中心粒側體組成。瓊凹大葉蟬精子細胞核末端平截、一側凹陷,由中心粒連接尾部軸絲,而凹陷區域出現中心粒側體連接鞭毛線粒體衍生物(圖5D,?5E)。從精子頸區的橫、縱切圖中可見,中心粒側體為致密片層狀結構,在精核凹陷區緊鄰細胞核,長度從凹陷區前端延伸至鞭毛軸絲區,連接精核和線粒體衍生物(圖5D,?5E,?6C,?6D,?7A)。
瓊凹大葉蟬精子尾部鞭毛較長,主要由1條軸絲,2條線粒體衍生物,2條副體,2條細小附結構及鞭毛中心物質組成(圖1C,?6E)。鞭毛中線粒體衍生物為1對長度和直徑大小對稱的結構,長度延伸至鞭毛末端,占據整個鞭毛的大部分區域。從縱切圖中可見,線粒體衍生物由一列垂直于縱軸、規則排列的嵴狀結構組成,其中嵴狀結構間隔13?nm,直徑38?nm(圖5E)。隨鞭毛長度向后延伸,線粒體衍生物橫切呈現出不同形狀,前端卵圓狀,中間兔耳狀,后端芒果狀(圖6C~F);且從橫切圖中可明顯看到線粒體衍生物具有電子稀疏區和電子致密區,其中底部電子致密區有晶狀體結構(圖7D)。此外,從橫、縱切圖中明顯可見線粒體衍生物基部區域中央具有電子稀疏的間隙區(圖6D,?6E,箭頭),且間隙區直徑隨線粒體衍生物長度延伸變大,之后消失不見(圖5E,?6D,?6E,?6F,?7B)。
鞭毛軸絲較長,始于中心粒延伸達整個鞭毛,具有典型的9+9+2型微管模式,即中央2個中心微管,內層9個雙微管,外圍9個副微管(圖6E,?6F)。其中軸絲副微管間有粗纖維,雙微管有動力蛋白臂及伸向中心微管的輻射鏈。沿鞭毛長度的延伸,鞭毛中副體和線粒體衍生物相繼消失,末端規則排列的軸絲結構逐漸變得雜亂無序(圖6A)直至消失。鞭毛中兩條副體平行于軸絲,位于軸絲和線粒體衍生物之間(圖6E)。副體結構細小,直徑約100?nm,橫切呈現規則的橢圓形或近圓形,同時副體內側出現細小附結構(圖6E,?7D)。副體長度較短,協同內側細小附結構隨鞭毛長度延伸而消失,僅出現在鞭毛起始區域,明顯短于軸絲和線粒體衍生物(圖6E,?6F)。此外,研究發現精子鞭毛基部,軸絲與線粒體衍生物中間還出現了鞭毛中心物質(圖7A~C)。
3?結論與討論
瓊凹大葉蟬具有葉蟬科已報道種類的精子形成及形態結構的共同特征:1)精子細胞內高爾基復合體形成精子頂體,線粒體融合形變為線粒體衍生物,中心粒產生軸絲結構;2)單層微管結構在精子形成過程中圍繞精子各組成部分,在精子成熟時消失;3)成熟精子細長、線狀,無多態性,頭部聚集形成精子束;4)圓錐狀頂體長度短,內部具有頂體腔,末端兩側多延伸2條突起插入細胞核頂端;5)圓柱狀細胞核染色質致密,末端平截、側邊多凹陷;6)頸區中心粒連接軸絲與平截細胞核,片層狀中心粒側體于細胞核凹陷區連接細胞核與線粒體衍生物;7)尾部鞭毛細長,多由1條9+9+2型軸絲,2條副體,2條線粒體衍生物組成;8)2條副體結構相對較小,隨鞭毛長度延伸最先消失;9)線粒體衍生物大小相等,由嵴狀結構組成,內部多具有晶狀體結構[2328,30]。但瓊凹大葉蟬也有其獨特的特征,即橫切呈五邊形的細胞核、電子稀疏間隙區直徑增大的線粒體衍生物。
線粒體衍生物作為昆蟲精子鞭毛中重要的結構組成部分之一,由昆蟲精子細胞內線粒體衍化形成,為精子運動提供能量。本研究中瓊凹大葉蟬線粒體衍生物基部區域具有直徑明顯增大的電子稀疏間隙區。在已報道葉蟬科種類中僅白邊大葉蟬具有該結構[27],但不同的是白邊大葉蟬間隙區直徑均一、無明顯變化趨勢。半翅目昆蟲除頭喙亞目廣蠟蟬類(間隙區小、不明顯,橫切多呈圓形),其他類群同大多數昆蟲一樣線粒體衍生物中無類似的間隙區結構[19,3132]。基于線粒體衍生物間隙區,我們認為大葉蟬亞科中凹大葉蟬屬與邊大葉蟬屬Kolla的親緣關系近,而非大葉蟬屬,這與孟澤洪等[33]根據線粒體Cyt?b基因得出的凹大葉蟬屬與邊大葉蟬屬親緣關系較近的研究結果相符合,但由于大葉蟬亞科昆蟲精子結構涉及種類較少,仍有待于進一步研究。此外,瓊凹大葉蟬精子鞭毛中出現了2條位于線粒體衍生物、軸絲和副體之間的細小附結構,以及位于鞭毛基部軸絲、線粒體衍生物之間的鞭毛中心物質,這與玉米黃翅葉蟬和短二室葉蟬的一致,也說明以上兩種結構并非角頂葉蟬亞科獨有的特征[26,28]。關于2條特殊的細小附結構,研究者曾認為該結構可能是由軸絲周圍膜池殘余物形成[26]。盡管也有類似的細小附結構出現于異翅亞目、長翅目、直翅目昆蟲精子鞭毛中[22,3437],但它們是否為同一類結構以及它們的來源、功能目前都尚不清楚,仍需開展更多深入的研究。
瓊凹大葉蟬精子發生過程中精子細胞外圍繞的單層微管結構,僅出現在精子形成期各組成部分伸長過程中,在精子成熟時消失不見,與已報道種類一致[27,30]。研究發現微管結構對精子細胞的形變和伸長起重要作用,且該結構與中心粒周圍物質密切相關[38]。精子發生過程中中心粒周圍物質參與了微管生成(主要為γ微管蛋白),而其剩余部分則會在精子形成后期沉積形成中心粒側體。大多數昆蟲的中心粒側體會沿著鞭毛向后伸長形成2條副體結構,雖然二者之間的直接聯系已被證實[17,38],但它們的具體功能還尚不清楚。本研究中瓊凹大葉蟬片層狀中心粒側體連接細胞核與線粒體衍生物,延伸至鞭毛軸絲區后形成2條長度較短、橫切呈橢圓形的副體結構。現有研究結果表明,葉蟬科昆蟲精子副體在不同亞科類群間差異較大,在角頂葉蟬亞科副體長度較長幾乎延伸至鞭毛端部,橫切呈不規則三角形[2526,28];而在大葉蟬亞科副體長度較短僅出現在鞭毛起始區域,橫切呈橢圓形[24,27]。此外,副體結構在頭喙亞目不同類群間也具有較大差異,其中蟬總科和沫蟬總科昆蟲缺少副體結構[3943],蠟蟬總科昆蟲副體結構異常發達[3132,44],角蟬總科昆蟲副體結構相對較為發達[2428,45]。由于在不同類群間表現出較大變異,我們推測副體結構很有可能為葉蟬科分類及半翅目頭喙亞目高級階元的系統發育分析提供有用的特征。
精子發生過程中精原細胞經過一系列分裂、形變最終成為成熟精子。節肢動物尤其是六足類雄性個體精子成熟后通常發生一定程度的精子接合形成精子束,以提高交配期間精子傳輸率,從而更有效地延續種族。關于精子接合方式,昆蟲有初級接合(源于自身精子發生過程,由單個精原細胞產生的全部精子頭部嵌入精囊細胞或包裹于帽狀物質內形成精子束)和次級接合(精子發生完成后游離精子通過特定物質接合在一起形成精子束)2種方式,其中次級接合被認為比初級接合更為進化[46]。已報道頭喙亞目類昆蟲貯精囊中均有精子束存在,且精子頭部多插入胞外基質。本研究中瓊凹大葉蟬精子在貯精囊內由繩狀物質連接頭部頂端形成梳狀精子束,與已報道葉蟬的彗星狀、沫蟬的球狀精子束差異較大,與蟬科昆蟲的繩狀精子束相類似[2526,28,3940,43]。結合精子發生特點及精子束形態,我們推斷瓊凹大葉蟬精子接合方式為次級接合,與Barcellos等[28]認為的沫蟬總科和角蟬總科精子初級接合方式觀點不同,與Higginson等[46]的觀點相一致。盡管研究證實凹大葉蟬精子束繩狀連接物質為透明蛋白質類物質[47],但其來源、作用機理仍不清楚。明確頭喙亞目各類群間精子接合方式、接合物質來源及其作用機理或許可為研究頭喙亞目種群進化提供相應的研究思路,但由于頭喙亞目昆蟲精子結構研究涉及類群種類較少,亟須開展更多的研究。
參考文獻
[1]?BARTLETT?C?R,?DEITZ?L?L,?DMITRIEV?D?A,?et?al.?The?diversity?of?the?true?hoppers?(Hemiptera:?Auchenorrhyncha)?[M]∥FOOTTIT?R?G,?ADLER?P?H.?Insect?biodiversity:?science?and?society,?Ⅱ.?New?Jersey:?John?Wiley?&?Sons?Ltd,?2018:?501590.
[2]?DIETRICH?C?H,?DDITZ?L?L.?Superfamily?Membracoidea?(Homoptera:?Auchenorrhyncha).?Ⅱ.?Cladistic?analysis?and?conclusions?[J].?Systematic?Entomology,?1993,?18:?297311.
[3]?HAMILTON?K?G?A.?The?grounddwelling?leafhoppers?Myerslopiidae,?new?family,?and?Sagmatiini,?new?tribe?(Homoptera:?Membracoidea)?[J].?Invertebrate?Taxonomy,?1999,?13:?207235.
[4]?CRYAN?J?R,?URBAN?J?M.?Higherlevel?phylogeny?of?the?insect?order?Hemiptera:?is?auchenorrhyncha?really?paraphyletic??[J].?Systematic?Entomology,?2012,?37:?721.
[5]?DIETRICH?C?H,?ALLEN?J?M,?LEMMON?A?R,?et?al.?Anchored?hybrid?enrichmentbased?phylogenomics?of?leafhoppers?and?treehoppers?(Hemiptera:?Cicadomorpha:?Membracoidea)?[J].?Insect?Systematics?and?Diversity,?2017,?1(1):?5772.
[6]?SONG?Nan,?CAI?Wanzhi,?LI?Hu.?Deeplevel?phylogeny?of?Cicadomorpha?inferred?from?mitochondrial?genomes?sequenced?by?NGS?[J/OL].?Scientific?Reports,?2017,?7:?10429.?DOI:?10.1038/s41598017111320.
[7]?JOHNSON?K?P,?DIETRICH?C?H,?FRIEDRICH?F,?et?al.?Phylogenomics?and?the?evolution?of?hemipteroid?insects?[J].?Proceedings?of?the?National?Academy?of?Sciences?of?the?United?States?of?America,?2018,?115(50):?1277512780.
[8]?SKINNER?R?K,?DIETRICH?C?H,?WALDEN?K?K?O,?et?al.?Phylogenomics?of?Auchenorrhyncha?(Insecta:?Hemiptera)?using?transcriptomes:?examining?controversial?relationships?via?degeneracy?coding?and?interrogation?of?gene?conflict?[J].?Systematic?Entomology,?2020,?45:?85113.
[9]?XUE?Qingquan,?DIETRICH?C?H,?ZHANG?Yalin.?Phylogeny?and?classification?of?the?leafhopper?subfamily?Eurymelinae?(Hemiptera:?Cicadellidae)?inferred?from?molecules?and?morphology?[J].?Systematic?Entomology,?2020,?45:?687702.
[10]CAO?Yanghui,?DIETRICH?C?H.?Phylogenomics?of?flavobacterial?insect?nutritional?endosymbionts?with?implications?for?Auchenorrhyncha?phylogeny?[J].?Cladistics,?2021,?38(1):?3858.
[11]DU?Yimin,?LIANG?Zonglei,?DIETRICH?C?H,?et?al.?Comparative?analysis?of?mitochondrial?genomes?of?Nirvanini?and?Evacanthini?(Hemiptera:?Cicadellidae)?reveals?an?explicit?evolutionary?relationship?[J].?Genomics,?2021,?113:?13781385.
[12]WANG?Xianyi,?WANG?Jiajia,?DAI?Renhuai.?Structural?features?of?the?mitogenome?of?the?leafhopper?genus?Cladolidia?(Hemiptera:?Cicadellidae:?Coelidiinae)?and?phylogenetic?implications?in?Cicadellidae?[J].?Ecology?and?Evolution,?2021,?11(18):?1255412566.
[13]YU?Tinghao,?ZHANG?Yalin.?Two?complete?mitochondrial?genomes?of?Mileewinae?(Hemiptera:?Cicadellidae)?and?a?phylogenetic?analysis?[J/OL].?Insects,?2021,?12(8):?668.?DOI:?10.3390/insects12080668.
[14]HU?Yanhua,?DIETRICH?C?H,?SKINNER?R?K,?et?al.?Phylogeny?of?Membracoidea?(Hemiptera:?Auchenorrhyncha)?based?on?transcriptome?data?[J].?Systematic?Entomology,?2023,?48(1):?97110.
[15]BIRKHEAD?T?R,?HOSKEN?D?J,?PITBICK?S.?Sperm?morphological?diversity?[M]∥BIRKHEAD?T?R,?HOSKEN?D?J,?PITBICK?S.?Sperm?biology:?an?evolutionary?perspective.?London:?Elsevier,?2009:?68149.
[16]奚耕思,?鄭哲民.?蝗總科精子超微形態比較及其在系統分類上的意義[J].?昆蟲分類學報,?1997,?19(1):?19.
[17]JAMIESON?B?G?M,?DALLAI?R,?AFZELIUS?B?A.?Insects:?Their?spermatozoa?and?phylogeny?[M].?Plymouth:?Science?Publishers,?1999:?447480.
[18]DALLAI?R.?The?contribution?of?the?sperm?structure?to?the?reconstruction?of?the?hexapod?phylogeny?[J].?Proceedings?of?Arthropodan?Embryological?Society?of?Japan,?2009,?43:?2338.
[19]DALLAI?R.?Overview?on?spermatogenesis?and?sperm?structure?of?Hexapoda?[J].?Arthropod?Structure?&?Development,?2014,?43:?257290.
[20]DALLAI?R,?GOTTARDO?M,?BEUTEL?R?G.?Structure?and?evolution?of?insect?sperm:?new?interpretations?in?the?age?of?phylogenomics?[J].?Annual?Review?of?Entomology,?2016,?61:?123.
[21]GOTTARDO?M,?DALLAI?R,?MERCATI?D,?et?al.?The?evolution?of?insect?sperm—an?unusual?character?system?in?a?megadiverse?group?[J].?Journal?of?Zoological?Systematics?and?Evolutionary?Research,?2016,?54:?237256.
[22]張貝貝.?長翅目昆蟲精子發生和精子超微結構研究[D].?楊凌:?西北農林科技大學,?2016.
[23]FOLLIOT?R,?MAILLET?P?L.?Ultrastructure?de?la?spermiogenèse?et?du?spermatozode?de?divers?insects?Homoptères?[M]∥BACCETTI?B.?Comparative?spermatology.?New?York:?Academic?Press,?1970:?289300.
[24]ZHANG?Beibei,?DAI?Wu.?Ultrastructure?of?the?spermatozoa?of?Cicadella?viridis?(Linnaeus)?and?its?bearing?on?the?phylogeny?of?Auchenorrhyncha?[J].?Micron,?2012,?43(9):?978984.
[25]SU?Mei,?DIETRICH?C?H,?ZHANG?Yalin,?et?al.?Ultrastructure?of?the?spermatozoa?of?Psammotettix?striatus?(Linnaeus)?and?Exitianus?nanus?(Distant)?(Hemiptera:?Auchenorrhyncha:?Cicadellidae:?Deltocephalinae)?[J].?Arthropod?Structure?&?Development,?2014,?43:?559570.
[26]VITALE?D?G?M,?DURSO?V,?VISCUSO?R.?Morphostructural?investigation?on?spermatozoa?and?sperm?bundles?of?the?leafhopper?Balclutha?brevis?Lindberg?1954?(Hemiptera:?Deltocephalinae)?[J].?Zoomorphology,?2016,?135:?441451.
[27]ZHANG?Beibei,?ZHANG?Chunni,?DIETRICH?C?H,?et?al.?Anatomy?and?ultrastructure?of?male?reproductive?system?in?two?leafhoppers?Cicadella?viridis?(Linnaeus)?and?Kolla?paulula?(Walker)?(Hemiptera:?Auchenorrhyncha:?Cicadellidae)?[J].?Annals?of?the?Entomological?Society?of?Amercia,?2016,?109(3):?463472.
[28]BARCELLOS?M?S,?DIAS?G,?LINONETO?J.?Redescription?of?the?sperm?morphology?of?Dalbulus?maidis?(Hemiptera:?Cicadellidae:?Deltocephalinae)?[J].?Tissue?&?Cell,?2018,?51:?5661.
[29]倪俊強,?楊茂發,?孟澤洪.?凹大葉蟬屬分類研究進展[J].?貴州農業科學,?2010,?38(11):?143146.
[30]KITAJIMA?E?W,?CRUZLANDIM?C.?An?electron?microscopic?study?of?the?process?of?differentiation?during?spermiogenesis?in?the?corn?leafhopper?Dalbulus?maidis?Del.?&?W.?(Homoptera:?Cicadellidae)?[J].?Biological?Reviews,?1972,?8:?519.
[31]江珍,?秦道正.?透明疏廣蠟蟬精子超微結構[J].?昆蟲學報,?2018,?61(2):?246254.
[32]JIANG?Zhen,?LIU?Jianing,?QIN?Daozheng.?Sperm?ultrastructure?of?Pochazia?shantungensis?(Chou?&?Lu)?and?Ricania?speculum?(Walker)?(Hemiptera,?Ricaniidae)?with?phylogenetic?implications?[J].?Zookeys,?2019,?880:?4359.
[33]孟澤洪,?楊茂發,?周玉鋒,?等.?基于Cyt?b?基因序列的大葉蟬亞科部分種類分子系統學分析[J].?貴州農業科學,?2013,?41(7):?15.
[34]杜喜翠,?石福明.?中華蟈螽與中華螽斯精子超微結構研究?(直翅目螽斯總科)?[J].?動物分類學報,?2005,?30(3):?461466.
[35]張貝貝,?謝莎,?花保禎.?劉氏蝎蛉的精子發生和精子超微結構[J].?昆蟲學報,?2017,?60(4):?389400.
[36]BACCETTI?B.?Spermatozoa?and?phylogeny?in?orthopteroid?insects?[M]∥BACCETTI?B.?Evolutionary?biology?of?orthopteroid?insects.?Chichester:?Halsted?Press,?1987:?12112.
[37]MERCATI?D,?GIUSTI?F,?DALLAI?R.?A?novel?membrane?specialization?in?the?sperm?tail?of?bug?insects?(Heteroptera)?[J].?Journal?of?Morphology,?2009,?270:?825833.
[38]DALLAI?R,?PAOLI?F,?MERCATIA?D,?et?al.?The?centriole?adjunct?of?insects:?Need?to?update?the?definition?[J].?Tissue?&?Cell,?2016,?48:?104113.
[39]CHAWANJI?A?S,?HODGSON?A?N,?VILLET?M?H.?Sperm?morphology?in?four?species?of?African?platylieurine?cicadas?(Hemiptera:?Cicadomorpha:?Cicadidae)?[J].?Tissue?&?Cell,?2005,?37:?257267.
[40]CHAWANJI?A?S,?HODGSON?A?N,?VILLET?M?H.?Sperm?morphology?in?five?species?of?cicadettine?cicadas?(Hemiptera:?Cicadomorpha:?Cicadidae)?[J].?Tissue?&?Cell,?2006,?38:?373388.
[41]崔貝貝,?魏琮.?四種蟬科昆蟲的精子形態?(半翅目:?蟬科)?[J].?昆蟲學報,?2018,?61(8):?9911002.
[42]CUI?Beibei,?WEI?Cong.?Ultrastructure?of?spermatozoa?in?three?cicada?species?from?China?(Hemiptera,?Cicadomorpha,?Cicadidae)?[J].?Zookeys,?2018,?776:?6180.
[43]HODGSON?AN,?RIDGEWAY?J?A,?VILLET?M?H.?Sperm?ultrastructure?and?spermatodesm?morphology?of?the?spittle?bug?Locris?transversa?(Thunberg?1822)?(Hemiptera:?Cercopidae)?[J].?Invertebrate?Reproduction?&?Development,?2016,?60(2):?8794.
[44]戴華國,?宋小玲,?吳小毅.?稻褐飛虱精子的超微結構[J].?南京農業大學學報,?1996,?19(2):?109111.
[45]ARAU'JO?V?A,?BO?S?N,?MOREIRA?J,?et?al.?Ultrastructural?characterization?of?the?spermatozoa?of?Aethalion?reticulatum?Linnaeus?1767?(Hemiptera:?Auchenorrhyncha:?Aethalionidae)?[J].?Micron,?2010,?41(4):?306311.
[46]HIGGINSON?D?M,?PITNICK?S.?Evolution?of?intraejaculate?sperm?interactions:?do?sperm?cooperate??[J].?Biological?Reviews,?2011,?86:?249270.
[47]HAYASHI?F,?KAMIMURA?Y.?The?potential?for?incorporation?of?male?derived?proteins?into?developing?eggs?in?the?leafhopper?Bothrogonia?ferruginea?[J].?Journal?of?Insect?Physiology,?2002,?48:?153159.
(責任編輯:楊明麗)
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
