芍藥雌配子發(fā)育和2n雌配子誘導1)
2024-05-11 09:40:38孫榕澤朱紹才崔雅琦趙家庚王宇暄于曉南
東北林業(yè)大學學報 2024年5期
孫榕澤 朱紹才 崔雅琦 趙家庚 王宇暄 于曉南
(北京林業(yè)大學,北京,100083)
多倍體植物通常具有莖稈健壯、花色豐富、花瓣多、葉片厚、抗性強的特點[1]。有性多倍化育種一直是芍藥屬(Paeonia)植物創(chuàng)制新品種的重點關(guān)注領(lǐng)域,即通過誘導配子減數(shù)分裂時期紊亂,從而形成2n配子參與受精培育多倍體的一種手段。已有研究證明,2n配子在植物系統(tǒng)進化中起重要作用,在傳遞雜合性和上位性中具有特殊價值[2]。同時,通過2n配子途徑培育的多倍體具有高雜合性和遺傳多樣性的特征[3]。目前,已有部分植物通過有性多倍化培育出多倍體品種[4-6]。
雖然已陸續(xù)有研究在多個物種中報道了天然2n配子的存在[7-8],但其發(fā)生概率低,難以被人為利用。人工誘導2n配子產(chǎn)生已成為一種多倍體育種的有效方法,包括2n花粉和2n雌配子的誘導。其中,有關(guān)2n花粉誘導的報道詳盡且深入[9-10],2n雌配子的探索則較為淺顯。但2n花粉的花粉競爭力差,難以競爭過單倍性花粉,導致多倍體得率較低[10]。而2n雌配子誘導在避免競爭問題的同時,還具有母系遺傳的優(yōu)勢[11]。大多數(shù)被子植物的細胞質(zhì)幾乎全部遺傳自母本[12],因此,誘導具有優(yōu)良性狀的母本2n雌配子,可能使其細胞質(zhì)遺傳物質(zhì)增加,進而遺傳給后代,使后代表現(xiàn)出更近似于母本的優(yōu)良性狀。
誘導2n配子產(chǎn)生,把握最佳處理時期是關(guān)鍵。大孢子母細胞經(jīng)歷2次減數(shù)分裂,產(chǎn)生功能大孢子,進入胚囊發(fā)育期,經(jīng)歷3次有絲分裂,最終形成雌配子。其他物種的研究表明,對減數(shù)分裂時期(大孢子發(fā)生期)[13-14]和胚囊發(fā)育期進行誘導都可能產(chǎn)生2n雌配子[15]。但目前,有關(guān)芍藥屬雌配子發(fā)育的細胞學研究較少,僅在黃牡丹(Paeoniadelavayivar.lutea(Delavay ex Franch.) Finet &Gagnep.)和‘鳳丹’牡丹(Paeoniaostia‘Fengdan’)中有相關(guān)報道。其中,王雁等[16]完整觀察了黃牡丹雌配子的發(fā)育過程,并建立雌配子體發(fā)育與外部形態(tài)的相關(guān)性。‘鳳丹’牡丹卻難以觀察整個雌配子發(fā)育進程的典型結(jié)構(gòu),但花蕾形態(tài)也與大孢子母細胞的發(fā)育階段呈現(xiàn)一定的對應(yīng)關(guān)系[13]。因此,芍藥屬雌配子發(fā)育的細胞學觀察存在一定的困難,但與花蕾的外部形態(tài)可以建立相關(guān)性,以即時辨別芍藥雌配子發(fā)育階段。
此外,芍藥屬已通過高溫誘導的物理方法成功誘導2n雌配子,并獲得了三倍體植株[13]。化學藥劑誘導法是目前使用最廣泛,也是誘導率最高的方法[17]。其中,秋水仙素是應(yīng)用最廣泛的化學誘變劑,對旺盛分裂的細胞作用效果優(yōu)秀。因此,秋水仙素誘導2n雌配子的方法非常適合于芍藥。
本研究以二倍體芍藥品種‘朱砂判’(Paeonialactiflora‘Zhushapan’)為材料,在掌握其雌配子發(fā)育過程的基礎(chǔ)上,利用秋水仙素誘導2n雌配子的產(chǎn)生,探究處理時期、秋水仙素質(zhì)量濃度、注射次數(shù)對2n雌配子誘導率的影響,為芍藥人工誘導2n雌配子,培育多倍體種質(zhì)途徑提供理論參考。
1 材料與方法
試驗材料:本研究選用的試驗材料為育性良好、性狀優(yōu)良的二倍體芍藥品種‘朱砂判’(Paeonialactiflora‘Zhushapan’,2n=2x=10),選擇生長勢良好、無病蟲害的植株,種植于國家花卉工程技術(shù)研究中心北京市昌平區(qū)小湯山基地,常規(guī)田間管理。
芍藥花蕾形態(tài)特征觀察:2022年、2023年4月末至5月中旬,于田間拍照并記錄花蕾形態(tài)特征,包括花蕾直徑、花瓣顏色、萼片顏色和雌蕊長度。其中,花蕾直徑和雌蕊長度使用標準游標卡尺測量,單位為mm。花蕾直徑劃分組別見表1。

表1 芍藥花蕾形態(tài)特征記錄
芍藥雌配子發(fā)育過程觀察:根據(jù)花蕾直徑分組,每一組分別用標準(FAA)固定液(V(體積分數(shù)50%的乙醇)∶V(冰醋酸)∶V(甲醛)=18∶1∶1)固定10個不同植株花蕾的全部心皮。固定后的心皮置于冰箱4 ℃保存,通過常規(guī)石蠟切片法對心皮進行細胞學觀察,切片厚度8~10 μm,番紅-固綠染色,切片置于徠卡光學顯微鏡下拍攝,同步統(tǒng)計發(fā)育時期及所占比例,建立雌配子發(fā)育過程與花蕾形態(tài)的關(guān)系。
秋水仙素誘導處理:采取注射法(即采用一次性注射器對花蕾緩慢注射試劑,至試劑從鱗片滲出為止),研究花蕾直徑、秋水仙素質(zhì)量濃度和注射次數(shù)3個因子對四倍體得率的影響。花蕾直徑(d)范圍為11 mm≤d<12 mm、14 mm≤d<15 mm、15 mm≤d<17 mm、23 mm≤d<26 mm、26 mm≤d<29 mm、29 mm≤d<32 mm。對照組為蒸餾水處理。對各直徑范圍的花蕾注射不同質(zhì)量濃度的秋水仙素(0、1、2、3 g/L),每個質(zhì)量濃度分為3組,分別注射1、2、3次,每組處理30朵花蕾。同步記錄花蕾存活數(shù)。
四倍體父本雜交:待試驗組和對照組雌蕊柱頭分泌粘液,去除母本花瓣,進行雜交工作。選擇花粉生活力良好及親和性良好的四倍體父本芍藥品種‘Cream Delight’授粉雜交[18],連續(xù)授粉3 d后,套硫酸紙袋以防止外來花粉污染,10 d后更換為雜交網(wǎng)袋,后續(xù)常規(guī)田間管理。
種子采收及常規(guī)管理:待雜種種子成熟后,根據(jù)試驗組合分別進行采收,通過赤霉素浸泡及沙藏處理促進萌發(fā)。種子生根后轉(zhuǎn)移至種植缽中繼續(xù)生長。在此過程中,記錄結(jié)實數(shù)量、種子總數(shù)、有胚種子數(shù)。
體細胞染色計數(shù)觀察:取雜種種子新生根尖,采用常規(guī)染色體壓片法,于徠卡光學顯微鏡下進行體細胞計數(shù)、觀察,統(tǒng)計四倍體后代數(shù)。倍性鑒定材料選取和處理參照朱煒等[18]的方法。
數(shù)據(jù)處理:采用Office軟件、SPSS 24整合和分析數(shù)據(jù)。其中,花蕾存活率=(花蕾存活數(shù)/處理數(shù))×100%,結(jié)實率=(結(jié)實數(shù)/花蕾存活數(shù))×100%,有胚率=(有胚種子數(shù)/種子總數(shù))×100%,四倍體得率=(四倍體種子數(shù)/有胚種子數(shù))×100%。采用Adobe Photoshop CC 2019和Image-Pro Plus 6.0處理圖片。
2 結(jié)果與分析
2.1 芍藥雌配子發(fā)育過程
大孢子發(fā)生進程:芍藥胚囊發(fā)育類型為蓼型,倒生胚珠,具有雙珠被結(jié)構(gòu)(圖1)。

圖1 芍藥胚珠結(jié)構(gòu)縱切面
芍藥大孢子母細胞由珠心的一個細胞分化而來,細胞體積大,細胞質(zhì)濃厚,細胞器豐富。芍藥同一胚珠內(nèi)可能存在一至多個大孢子母細胞(圖2A、B),這些大孢子母細胞不同步進入減數(shù)分裂時期或退化。大孢子母細胞繼續(xù)發(fā)育,陸續(xù)進入第一次減數(shù)分裂(圖2C~F),第一次減數(shù)分裂結(jié)束,大孢子母細胞形成二分體,進入第二次減數(shù)分裂前期(圖2G),最終二分體分裂成縱向排列的四分體大孢子(圖2K)。多數(shù)胚珠中,僅合點端的大孢子具有功能性,發(fā)育成功能大孢子(圖2L),其余大孢子退化。極少數(shù)情況下,珠孔端大孢子繼續(xù)發(fā)育。大孢子母細胞完成減數(shù)分裂,進入胚囊發(fā)生階段。
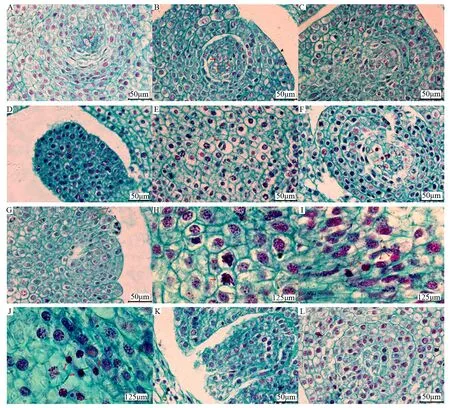
A為間期(單個大孢子母細胞);B為間期(多個大孢子母細胞);C為減數(shù)分裂I前期;D為減數(shù)分裂I中期;E為減數(shù)分裂I后期;F為減數(shù)分裂I末期;G為減數(shù)分裂II前期;H為減數(shù)分裂II中期;I為減數(shù)分裂II后期;J為減數(shù)分裂II末期;K為四分體;L為功能大孢子。圖中紅色箭頭表示處于該發(fā)育時期的細胞。圖2 芍藥大孢子發(fā)生過程
胚囊發(fā)生進程:進入胚囊發(fā)生階段后,功能大孢子體積增大,逐漸變得細長,產(chǎn)生液泡,形成單核胚囊(圖3A)。隨后發(fā)生3次有絲分裂,逐步形成二核胚囊(圖3B)、四核胚囊(圖3C)和八核胚囊(圖3D)。伴隨胚囊的成熟,珠孔端和合點端模糊,中央部分細長。此外,同一胚珠中可能同時存在不同發(fā)育階段的胚囊,多個胚囊結(jié)構(gòu)的堆疊使得胚囊結(jié)構(gòu)難以辨認(圖3E)。
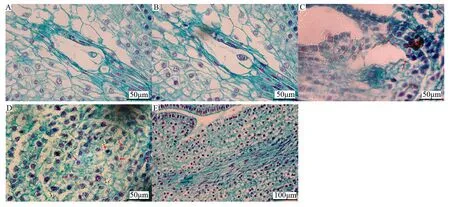
A為單核胚囊;B為二核胚囊;C為四核胚囊;D為八核胚囊;E為多個非典型胚囊結(jié)構(gòu)。圖中紅色箭頭表示處于該發(fā)育時期的細胞。圖3 芍藥胚囊發(fā)生過程
雌配子發(fā)育過程與花蕾形態(tài)的關(guān)系:芍藥雌配子發(fā)育過程與花蕾形態(tài)呈現(xiàn)一定的對應(yīng)關(guān)系,一個花蕾直徑范圍對應(yīng)一個主要的雌配子發(fā)育階段,即處于該發(fā)育階段的雌配子數(shù)量占全部雌配子數(shù)量的比例最大。同時,跟蹤觀察了不同花蕾直徑范圍對應(yīng)的其他花蕾形態(tài)特征,包括萼片顏色、花瓣顏色、花瓣軟硬程度和雌蕊長度。具體可見表1、2和圖4。因此,花蕾形態(tài)可以作為判斷芍藥雌配子發(fā)育過程的判斷指標。
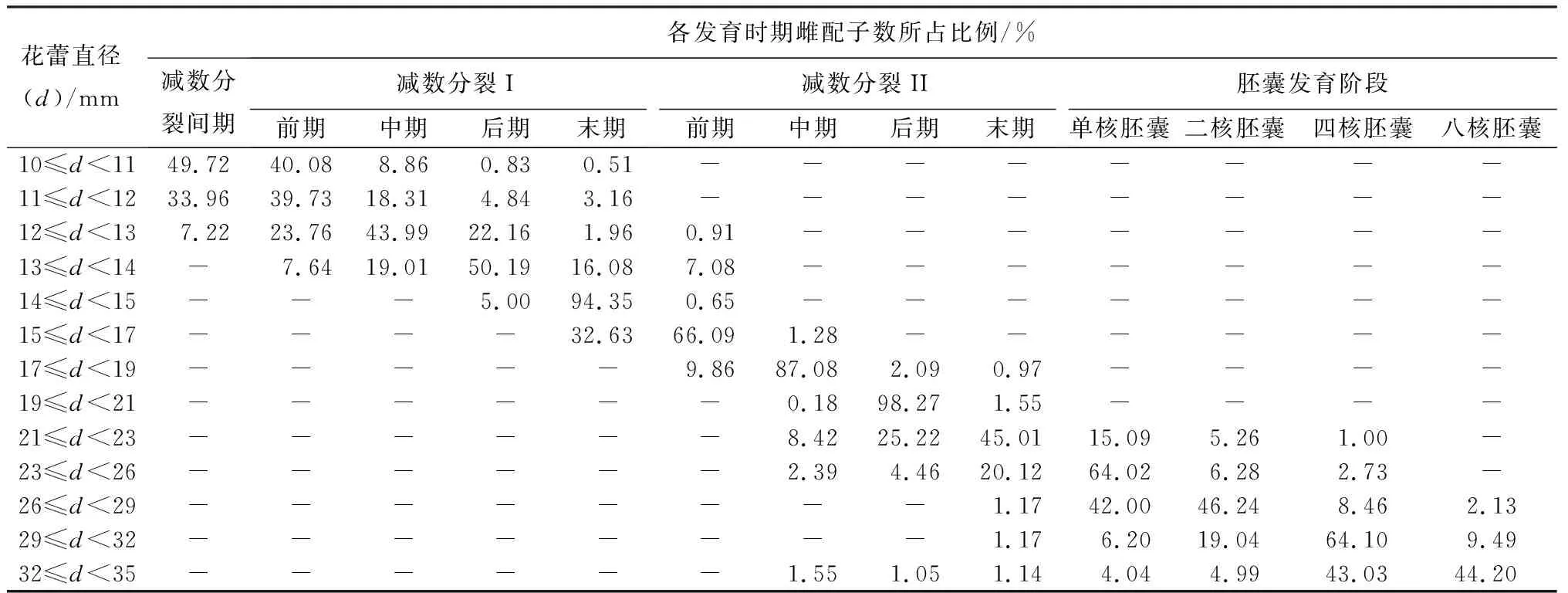
表2 不同直徑花蕾中各發(fā)育時期雌配子數(shù)所占比例

A為10 mm≤d<11 mm;B為11 mm≤d<12 mm;C為12 mm≤d<13 mm;D為13 mm≤d<14 mm;E為14 mm≤d<15 mm;F為15 mm≤d<17 mm;G為17 mm≤d<19 mm;H為19 mm≤d<21 mm;I為21 mm≤d<23 mm;J為23 mm≤d<26 mm;K為26 mm≤d<29 mm;L為29 mm≤d<32 mm;M為32 mm≤d<35 mm。d為花蕾直徑。圖4 芍藥花蕾形態(tài)發(fā)育進程
2.2 秋水仙素誘導芍藥大孢子染色體加倍
部分被子植物大孢子減數(shù)分裂進程的細胞學觀察研究[17,19]表明,第一次減數(shù)分裂前期和第二次減數(shù)分裂前期誘導效率最佳。而牡丹的研究表明,第一次減數(shù)分裂末期也能作為大孢子染色體加倍的適宜時期[13]。故本研究誘導的花蕾直徑范圍為11 mm≤d<12 mm(減I前期主導期)、14 mm≤d<15 mm(減I末期主導期)、15 mm≤d<17 mm(減II前期主導期)。共處理1 080朵花蕾,收獲有胚種子2 322個。經(jīng)根尖染色體壓片鑒定(圖5),篩選出7個四倍體種子,均來自試驗組,對照組均未獲得四倍體種子(表3)。
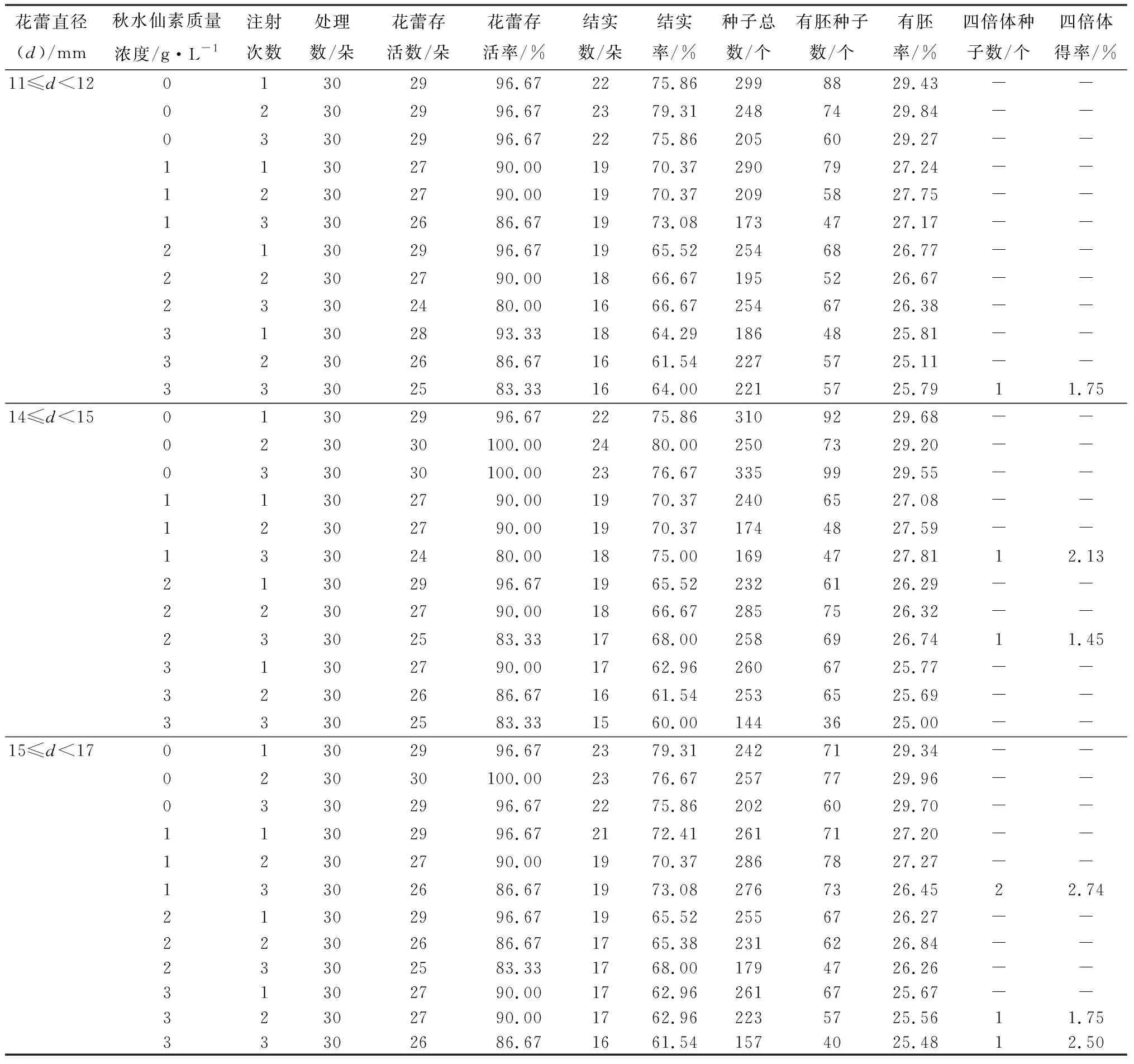
表3 秋水仙素誘導芍藥大孢子染色體加倍選育四倍體得率

圖5 芍藥大孢子染色體加倍獲得四倍體后代的體細胞染色體
由表3可知,3個減數(shù)分裂時期均成功獲得四倍體種子。進一步對2n雌配子誘導率減數(shù)分裂進程進行相關(guān)性分析,結(jié)果表明,2n雌配子誘導率和減數(shù)分裂I前期(r=0.496,p=0.284)、減數(shù)分裂I末期(r=0.547,p=0.168)、減數(shù)分裂II前期(r=0.044,p=0.925)的主導期雌配子數(shù)量占比(即主導期雌配子數(shù)量占全部雌配子數(shù)量的比)呈正相關(guān)。據(jù)此判斷,在減數(shù)分裂I前期、減數(shù)分裂I末期和減數(shù)分裂II前期均可以誘導雌配子加倍。其中,在減數(shù)分裂II前期,質(zhì)量濃度為1 g/L的秋水仙素注射3次的誘導效果最好,2n雌配子誘導率為2.74%。
2.3 秋水仙素誘導芍藥胚囊染色體加倍
芍藥功能大孢子經(jīng)過3次有絲分裂形成成熟胚囊。故3次有絲分裂給胚囊染色體加倍提供了可能性。因芍藥八核胚囊結(jié)構(gòu)的不典型性,本研究僅探討前兩次有絲分裂的誘導效果,即單核胚囊期(23 mm≤d<26 mm)、二核胚囊期(26 mm≤d<29 mm)和四核胚囊期(29 mm≤d<32 mm)。共處理1 080朵花蕾,獲得有胚種子數(shù)2 408個,經(jīng)根尖染色體壓片法鑒定(圖6),獲得9個四倍體種子(表4),均來自試驗組。對照組未獲得四倍體種子。
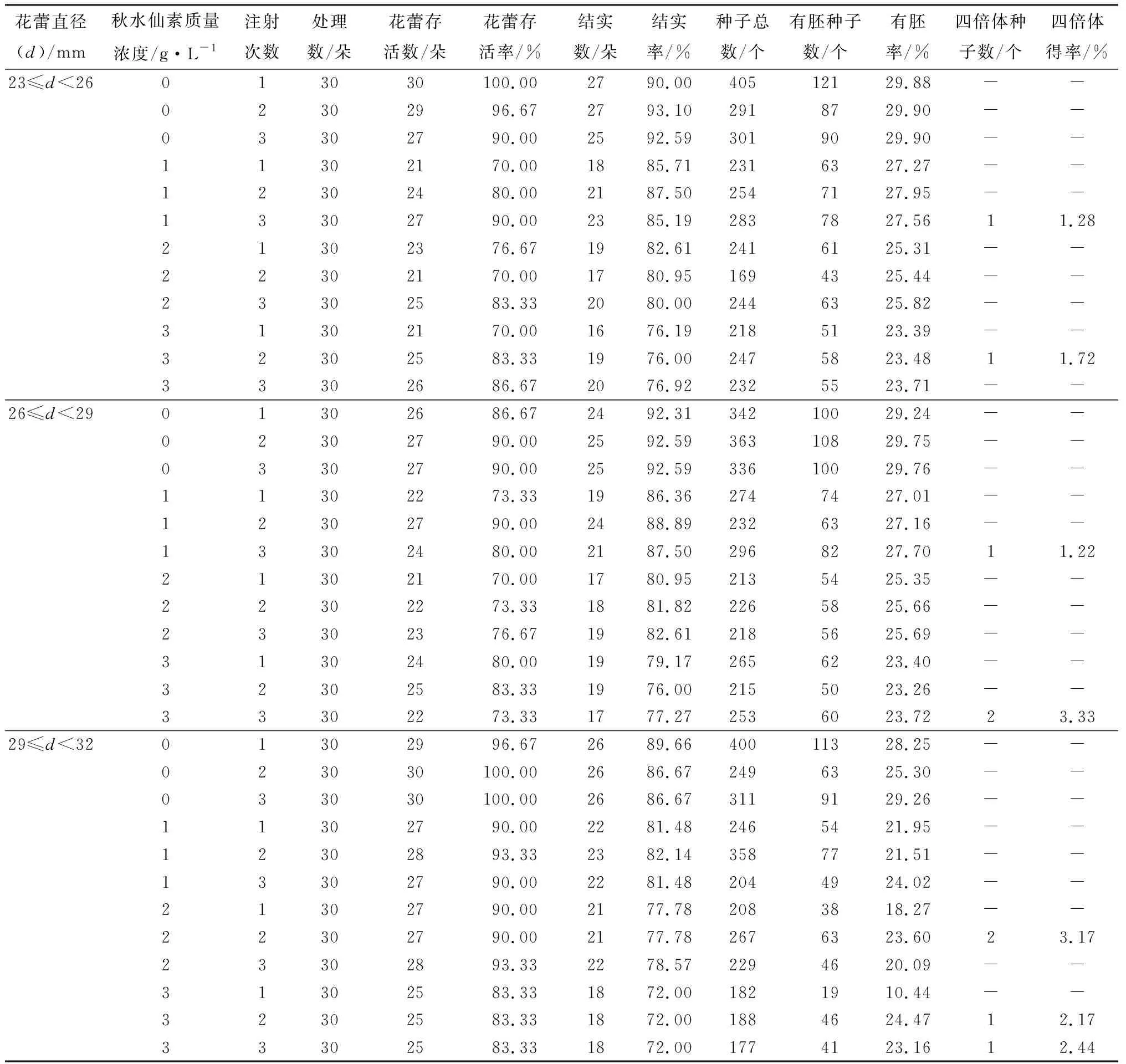
表4 秋水仙素誘導芍藥胚囊染色體加倍選育四倍體得率

圖6 芍藥胚囊染色體加倍獲得四倍體后代的體細胞染色體
單核胚囊期、二核胚囊期和四核胚囊期均能獲得一定比率的四倍體種子。進一步對胚囊發(fā)育時期中主導期雌配子數(shù)量占比和四倍體得率進行相關(guān)性分析,結(jié)果表明,四倍體得率與單核胚囊主導期雌配子數(shù)量占比(r=-6.980,p=0.258)呈負相關(guān),與二核胚囊(r=0.448,p=0.403)和四核胚囊(r=0.641,p=0.389)主導期雌配子數(shù)量占比呈正相關(guān)。綜上,在胚囊發(fā)育的前兩次有絲分裂過程施加處理均可獲得四倍體種子。其中,質(zhì)量濃度為3 g/L的秋水仙素對二核胚囊期的材料注射3次誘導效果最佳,誘導率為3.33%。該結(jié)果較之減數(shù)分裂時期處理組合的最佳誘導率略高,可視為芍藥2n雌配子人工誘導的最佳組合。
3 結(jié)論與討論
目前,芍藥屬植物配子發(fā)育規(guī)律的研究,主要集中在對雄配子的研究上[20-22]。本研究發(fā)現(xiàn),芍藥雌配子發(fā)育過程為蓼型。胚珠內(nèi)具有一至多個具有發(fā)育潛力,且發(fā)育不同步的大孢子母細胞。即胚珠內(nèi)可同時觀察到不同發(fā)育時期的大孢子、胚囊和退化細胞。當大孢子進入胚囊發(fā)育期,伴隨3次有絲分裂,胚囊拉伸為細長的結(jié)構(gòu)。加州芍藥(Paeoniacalifornica)的研究結(jié)果稱這種細長結(jié)構(gòu)為“珠孔端擴大、合點端模糊,中央部分極細長”[23]。但在本研究中,難以通過石蠟切片法觀察到形態(tài)清晰的典型成熟胚囊結(jié)構(gòu)。同時,在多胚囊胚珠中,胚囊結(jié)構(gòu)的堆疊使得胚囊結(jié)構(gòu)的觀察更加困難。這種多胚囊發(fā)育的特性與黃牡丹雌配子發(fā)育觀察結(jié)果不一致。黃牡丹的多個大孢子母細胞在經(jīng)減數(shù)分裂后,形成多個功能大孢子,但在同一胚珠內(nèi)只觀察到了一個胚囊結(jié)構(gòu)[16]。此外,在本研究中,雖觀察到芍藥的多胚囊結(jié)構(gòu),但目前仍未有報道顯示,芍藥屬種子具多胚結(jié)構(gòu)。故推測在雌配子發(fā)育和胚胎發(fā)育的過程中可能存在“淘汰”機制:①成熟胚囊形成前,不同胚囊之間相互存在競爭和選擇,只有那些在競爭中取得優(yōu)勢的胚囊才能進一步發(fā)育成熟,獲得受精的資格;②多個胚囊均能發(fā)育成熟并受精,除某一獲得競爭優(yōu)勢的胚胎外,其余胚胎的淘汰必然發(fā)生在胚胎發(fā)育的某一時期。
本研究建立花蕾形態(tài)與芍藥雌配子發(fā)育過程的關(guān)聯(lián)性,能夠做到對雌配子發(fā)育階段的即時性判斷,最大程度提高2n雌配子的誘導率。其中,不同花蕾直徑范圍對應(yīng)一個雌配子發(fā)育主導時期,主導時期雌配子數(shù)量占比對2n雌配子誘導率存在一定影響。如,在對芍藥大孢子染色體加倍的研究中,最佳誘導時期在減數(shù)分裂II前期,表明而非減數(shù)分裂I前期。花蕾直徑在11 mm≤d<12 mm范圍內(nèi),大孢子發(fā)生主導時期(減數(shù)分裂I前期)雌配子數(shù)量占比僅有39.73%,有33.96%的大孢子母細胞處于間期階段。而花蕾直徑在15 mm≤d<17 mm范圍內(nèi),大孢子發(fā)生主導時期(減數(shù)分裂II前期)所占比例高達66.09%。
后續(xù)工作中,需進一步探索更適宜芍藥雌配子發(fā)育過程的石蠟切片技術(shù),可輔以其他技術(shù)手段,如胼胝質(zhì)沉積定位等[24],以追求更準確、具體的發(fā)育過程細胞學觀察,建立更高效的芍藥2n雌配子誘導體系。同時,對芍藥雌配子發(fā)育和胚胎發(fā)育過程中可能存在的“淘汰”機制進行深入研究。
