產氣莢膜梭菌致病機制及其治療性抗體研究進展
2024-05-07 08:53:18趙品楠羅龍龍
中國藥理學與毒理學雜志 2024年4期
趙品楠,羅龍龍
(軍事科學院軍事醫學研究院毒物藥物研究所,國家安全特需藥品全國重點實驗室,北京 100850)
產氣莢膜梭菌(Clostridiumperfringens,CP)又稱魏氏梭菌(C.Welchii),主要存在于土壤、污水以及人和動物腸道中,可引發多種疾病,包括人氣性壞疽、食物中毒、動物壞死性腸炎和腸毒血癥等,嚴重危害公共衛生安全[1]。美國病理學家William H.Welch 于1982 年在1 例尸檢過程中分離得到CP,CP 的獨特表現是在被感染的血管中能觀察到氣泡。開放性創傷使CP 感染的控制更加困難。第一次世界大戰(一戰)期間,約1%的士兵死于CP感染[2]。二戰期間,由于戰爭波及到更多國家,武器機械化程度更高,殺傷性更強,導致的傷亡人數和受傷程度大幅上升,戰傷使感染CP 的比例大幅增加,氣性壞疽發生率由一戰的1%增長至10%[3]。國內同樣發生過CP群體感染事件。2008年汶川地震時,由于大面積開放性創傷、惡劣環境及炎熱天氣,CP迅速傳播,出現近百例氣性壞疽患者[4]。另外,CP可以引起大規模禽類壞死性腸炎和羊腸毒血癥,給養殖業造成巨大的經濟損失[1]。因此,CP對公共衛生的影響不容小覷,如何有效控制CP 感染引起的各種疾病的發生,找到有效的防治方案迫在眉睫。目前,針對CP 感染的主要治療手段還是及時清創并大量使用抗生素,但由于多重耐藥菌株的大量出現、耐藥譜的擴大以及不同血清型混合感染等問題,亟需新型的預防和治療策略。本文簡述CP 的主要致病因子、致病機制和基因工程抗體治療研究進展,為CP感染防治提供參考。
1 主要致病因子和致病機制
CP 是一種革蘭陽性粗大桿菌,有莢膜,無鞭毛,不能運動;芽孢位于次級端,呈橢圓形;抗熱能力很強,可在114 ℃的高溫下存活1~4 h,這也是其廣泛存在、傳播力強的主要原因。CP 屬于厭氧菌,但并非絕對厭氧,在微量氧的情況下也能生長。該細菌培養有兩大特點:①菌落在血平板上形成雙層溶血環(內環由θ 毒素引起完全溶血,外環由α 毒素引起不完全溶血);②在牛乳培養基中因分解乳糖產酸而使酪蛋白凝固,同時產生大量氣體,將凝固的酪蛋白沖成蜂窩狀,該現象被稱為“洶涌發酵”。
水解酶和外毒素是CP 最主要的致病因子。CP 可分泌近20 余種毒素,其中最主要的致病毒素為α,β,ε 和τ 毒素及腸毒素(C.perfringensenterotoxin,CPE)和壞死性腸炎B 樣毒素(necrotic enteritis B-like toxin,NetB)。另外,根據其主要產生的毒素可將CP 分為7 種血清型,分別是A 型(α 毒素)、B 型(α,β 和ε 毒素)、C 型(α 和β 毒素)、D 型(α 和ε 毒素)、E 型(α 和τ 毒素)、F 型(α 毒素和CPE)和G 型(α 毒素和NetB)[2]。由于各型CP 的宿主和主要致病毒素不同,其導致的疾病也不同,其中A型為最常見類型,主要引起人氣性壞疽、食物中毒和家禽壞死性腸炎等;B,C 和G 型導致綿羊和反芻動物壞死性腸炎等;D 和E 型引起動物腸毒血癥;F型引起人食物中毒和抗生素相關性腹瀉等。
1.1 α毒素
1.1.1 分子結構
研究發現,各型CP 均可產生α 毒素。α 毒素是A 型CP最主要的毒力因子,也是第1個被發現既有酶活性又有毒素特性的細菌蛋白[5]。α毒素由370個氨基酸組成,具有磷脂酶C 活性,編碼基因位于細菌染色體的穩定區域,包括β 折疊組成的膜結合C端結構域、α 螺旋組成的催化N 端結構域和含有神經節苷脂結合位點的中心環結構域[6],在鈣離子存在時結合至宿主細胞膜上[7],通過破壞細胞膜結構導致細胞裂解壞死。N端和C端單獨存在時無溶血活性,只有兩者共同作用時α毒素才具有毒性[8]。
1.1.2 致病機制
α 毒素同時含磷脂酶和鞘磷脂酶活性,可水解質膜上的卵磷脂和鞘磷脂,產生二酰基甘油和神經酰胺[6];還可通過與Gi型GTP結合蛋白的相互作用激活內源性磷脂酶和鞘磷脂酶[9],通過一系列級聯反應和信號通路破壞細胞膜結構的完整性,引起細胞裂解,并促使白細胞介素8(interlukin-8,IL-8)、活性氧和鈣離子升高,從而表現出毒素的細胞毒性、致死性和溶血活性等[10]。此外,α 毒素作用不僅限于膜破壞,還能特異性結合神經節苷脂類糖,隨后促進其聚集和酪氨酸激酶A的激活,觸發IL-8釋放,繼而通過招募和激活中性粒細胞引起細胞急性炎癥,最終導致細胞死亡[11-12](圖1)。Oda等[13]報道,IL-8升高可促進嗜中性粒細胞與細胞外基質蛋白黏附,促使嗜中性粒細胞在血管內皮上積聚,最終導致氣性壞疽發生。還有研究報道,α 毒素可誘導外周血中性粒細胞分化[14],該作用歸因于外周血中性粒細胞中含神經節苷脂類糖的脂筏改變[15]。在病理方面,α 毒素能夠影響免疫細胞如嗜中性粒細胞向感染組織轉移(機制目前未知),因此其可潛在地減少感染部位的病原體清除。α 毒素還可導致血管收縮,減少組織的血液供應,從而產生有利于CP 生長的低氧環境。另外,α 毒素所引發的代謝中的炎癥級聯反應(花生四烯酸和蛋白激酶C)是導致細胞死亡的主要原因[2]。人氣性壞疽主要由A 型CP 導致,尤其發生在戰爭或地震環境中。由于α 毒素能分解肌肉和組織中的糖類物質,從而產生大量氣體,造成氣腫;同時血管通透性增加,局部發生水腫,擠壓周圍軟組織和血管,影響血供,造成組織壞死。
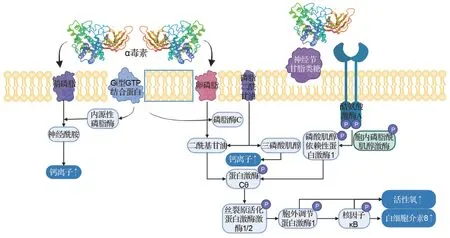
圖1 α毒素致病機制.↑:升高;:磷酸化.
1.2 腸毒素
1.2.1 分子結構
流行病學研究表明,CPE 是引起F 型CP 的人類食物中毒及非食源性腹瀉的主要毒素[16],該毒素缺乏與其他毒素一級氨基酸序列的同源性,但在結構上屬于成孔毒素家族[17]。CPE基因位于染色體或質粒上,毒素的表達僅發生在孢子形成期間[18]。CPE由含有319個氨基酸的單一蛋白組成[19],CPE 蛋白由無毒的C端受體結合結構域和N端細胞毒性結構域組成,其在孔形成期間介導寡聚化和膜插入[20]。
1.2.2 致病機制
緊密連接蛋白是CPE 的細胞受體,對于維持由上皮和內皮細胞形成的緊密連接的結構和功能具有重要作用[21]。初始CPE 與緊密連接蛋白的結合導致小復合物形成,6 個小復合物的相互作用可導致CPE 低聚和質膜上孔的形成,最終形成六聚體復合物1(hexamer complex-1,CH-1)[22]。該復合物將來自CPE 的β-發卡環組裝成β-桶狀結構,其插入膜中形成增強離子內流的活性孔,從而導致細胞死亡[23](圖2)。CPE 孔可滲透小分子,特別是陽離子[24]。CPE 較低劑量即可引起鈣離子內流,誘導鈣蛋白酶活化觸發胱天蛋白酶3 介導的細胞凋亡;而CPE 較高劑量引起大量鈣離子內流,可誘導強烈的鈣蛋白酶活化并導致混合系列蛋白激酶樣結構域依賴性壞死性細胞凋亡。CPE 對人類健康危害極大,在腸道中其不但引起上皮細胞死亡,導致體液積聚和電解質損失,導致腹瀉;而且還誘導腸上皮組織損傷,包括絨毛縮短、上皮壞死和脫屑,使得CPE 進入血循環并進入肝,在肝中與緊密連接蛋白3 結合,誘導可能危及生命的高鉀血癥[25]。典型F 型CP 的CPE 黏附于腸黏膜上皮,阻礙營養物質的輸運和吸收,且通過改變細胞膜的通透性,造成小分子的流失進而引起腹瀉或食物中毒。
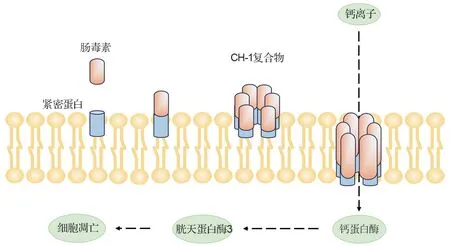
圖2 腸毒素致病機制.CH-1:六聚體復合物1.
1.3 ε毒素
1.3.1 分子結構
ε 毒素屬于氣溶素樣成孔毒素家族,是B 型和D 型CP 菌的主要致病毒素,可引起腸毒血癥。腸毒血癥是一種高度致死性疾病,對家養反芻動物,特別是綿羊養殖具有重大影響[26]。在小鼠中其半數致死劑量(LD50)為100 ng·kg-1,是繼肉毒桿菌毒素和破傷風毒素之后已知的第3種最有效的細菌毒素[27]。由于這種高效力,ε 毒素被認為是一種潛在的生物戰劑,并已被美國疾病控制和預防中心歸為B 類生物戰劑[28]。ε 毒素具有3 個由β-折疊組成的結構域。結構域Ⅰ含有1 個大的α-螺旋,隨后是1 個環和1 個短α-螺旋。在結構域Ⅰ和Ⅱ之間,存在另1 個α-螺旋,隨后是1 個環。結構域Ⅱ是由2 個反向平行的β-折疊和β-發夾組成的β-夾心。結構域Ⅲ也是具有2 個β-折疊的β-夾心結構,且這2個折疊中的第2個形成毒素的C端[29]。
1.3.2 致病機制
ε 毒素發揮毒性作用的過程大致為:ε 毒素被胰蛋白酶和胰凝乳蛋白酶激活[26];活化的ε 毒素與位于細胞膜區域中的受體結合[30],促進中性鞘磷脂酶激活,導致鞘磷脂水解和神經酰胺生成[31],從而促進寡聚體形成。寡聚化導致七聚體孔,誘導鉀離子快速丟失,氯離子和鈉離子進入,隨后鈣離子增加,最終導致細胞損傷[31-32](圖3)。有研究表明,這些七聚體也被內化并引起細胞損傷[33]。腸中ε毒素的作用可導致體液聚積和黏膜出血,小腸和大腸的組織病理變化包括黏膜充血和糜爛及中度多形核細胞浸潤等。ε 毒素除被腸道吸收外,其他靶器官如腎、肺和腦中也發現了該毒素[34]。
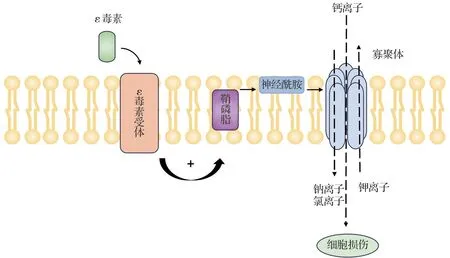
圖3 ε毒素致病機制.+:促進.
2 臨床治療和耐藥性
目前,臨床上針對CP 感染最常用的治療方法還是大劑量使用抗生素,對于嚴重的氣性壞疽需采取緊急外科處理,包括及時反復沖洗傷口和切除壞死組織等。但由于該菌導致的疾病具有發病急、病程短和死亡率高等特點,臨床用藥具有一定的滯后性,很難達到較好的治療效果。另外,盡管抗生素可用于大部分細菌感染,大幅降低細菌感染后的死亡率,然而全球許多常見菌正在對抗生素產生耐藥性,導致長期慢性感染性疾病頻發,CP 也不例外。青霉素、四環素、氯霉素和甲硝唑是臨床治療CP 感染的常規藥物[35],但由于抗生素的不規范使用和濫用,使得CP 耐藥性不斷增強甚至發生多重耐藥。2004 年,瑞典、丹麥和挪威分離的CP 菌株均存在對四環素的耐藥,耐藥率分別為10%,29%和80%,并且通過PCR 擴增到了2 個抗性基因tetA(P)和tetB(P)[36],它們分別編碼外排蛋白和核糖體保護型蛋白[37],通過干擾抗生素與核糖體的結合發揮作用。Gholamiandehkordi 等[38]從肉雞中分離的CP對四環素耐藥率高達66%。Kartalidis 等[39]報道,CP 中存在糖肽類抗生素耐藥基因,且萬古霉素耐藥基因的存在已得到證實。眾所周知,抗生素抑菌機制主要是抑制細菌細胞壁合成或抑制細菌蛋白質合成,包括細菌滅活酶的產生、細菌藥物靶點改變和細菌細胞膜通透性改變等[40]。而CP 主要致病物質是其產生的外毒素和水解酶,并非細菌本身。因此,抗生素對CP 感染的治療既達不到完全有效又會產生耐藥性。目前疫苗預防和抗體治療已成為研究的突破點。
3 基因工程治療性疫苗和抗體
隨著科學技術發展和醫藥研究進入新階段,抗體藥物呈現蓬勃發展的態勢,在腫瘤、免疫系統疾病和感染性疾病等方面均取得突破性的進展,也因此逐漸成為新藥開發的熱點。單克隆抗體藥物經歷了從鼠源單抗、人鼠嵌合抗體、人源化抗體到全人源化抗體的發展[41],上述技術的迭代逐步優化抗體藥物的免疫原性和半衰期,對疾病和治療效果呈跨越式提高,不良反應也明顯降低[42]。傳統的單克隆抗體具有完整的“Y”字形抗體結構,包括與抗原結合的Fab 段和誘導抗體依賴的細胞介導的細胞毒性作用、補體依賴的細胞毒性作用的可結晶片段(Fc 段)。然而隨著抗體工程化技術的發展和對抗體藥物的深入研究,近年來衍生出了多種非天然的新型抗體結構模式,包括抗體-融合蛋白[43]、雙特異性抗體[44]和抗體偶聯藥物[45]等。臨床實踐證明,應用抗體治療CP 感染具有一定的療效,無論是鼠源性單抗還是全人源單鏈抗體,均針對CP 分泌的外毒素,協同抗生素對CP 繁殖的抑制,產生更加有效的保護作用[46]。
3.1 抗α毒素疫苗和抗體
目前,許多抗CPα 毒素的疫苗被認為是CP 感染有效的預防劑。Hoang 等[47]等報道,表達α 毒素的重組枯草芽孢桿菌是針對壞死性腸炎的候選口服疫苗。Uppalapati 等[48]研制出二價重組蛋白,這可能是針對CP 診斷和治療領域的一種新型干預措施。研究發現,缺乏任何酶和毒素活性的膜結合C端結構域可能是α 毒素疫苗的主要免疫原[49]。最近,Shreya 等[50]報道了用二價嵌合蛋白(由α 毒素和CPE 的C 端結構域組成)免疫小鼠可預防A 型CP感染。
除疫苗外,針對α 毒素的抗體因具有高效的中和效應被廣泛關注。Logan等[51]和Singh等[52]通過將骨髓瘤細胞與用戊二醛滅活或甲醛滅活的α毒素免疫的小鼠脾細胞相融合,制備了抗α 毒素的鼠單克隆抗體。該單克隆抗體與α毒素反應強烈并特異性結合,可中和α毒素的磷脂酶C活性、溶血活性和致死作用。趙寶華等[53]利用實時熒光定量PCR 和分子克隆技術制備單鏈抗體scFv-1A8,其由抗α 毒素雜交瘤細胞系的重鏈和輕鏈的可變區組成,并用柔性的短接頭肽連接。該單鏈抗體在體外能中和α 毒素的磷脂酶C 活性,在小鼠模型中對致死量α毒素攻擊具有保護作用。scFv 的優點是其相對分子質量較低,滲透性較強[54]。Wang 等[55]等從天然合成的噬菌體展示文庫中制備了抗α毒素的人源單鏈抗體,并以小鼠為模型檢測了其預防和治療效果。為證明scFv 中和毒素的潛力,將α 毒素與scFv 預孵育,隨后測試其卵磷脂酶和溶血活性以及iv 給予小鼠后的致死作用。經分析發現,該單鏈抗體可抑制卵磷脂酶和溶血活性。另外,用2 倍LD50的α 毒素攻擊小鼠,發現在30 min 內用scFv 處理小鼠的存活率可達80%。上述結果均表明,制備的抗A 型CPα 毒素人源單鏈抗體可用于預防和治療α毒素相關疾病。但由于存在人抗鼠免疫反應[56]、單鏈抗體結合力低和易被腎清除等缺點,治療效果被限制。
王冬冬等[57]在抗A 型CPα 毒素單鏈抗體的基礎上,利用基因工程技術構建了雙價單鏈抗體。通過體外檢測該抗體抑制α毒素水解卵磷脂的活性和溶血活性以及小鼠攻毒保護實驗,初步研究該雙價單鏈抗體的生物學活性。結果表明,構建的雙價單鏈抗體scFv2-5 和scFv2-15 均正確,并且與α 毒素具有特異結合活性,scFv2-15 和scFv2-5 與抗原的結合活性明顯高于scFv,且scFv2-15 中和毒素的能力較scFv2-5 和scFv 具有明顯優勢。邱玥等[58]構建了抗A 型CPα 毒素單鏈抗體,是利用全人源噬菌體抗體庫篩選獲得的抗α 毒素的全人源scFv 抗體。他們構建5種不同組分的重組表達質粒后轉入表達菌,獲得5 個單分子抗體的表達菌。利用EILSA 檢測該5 種單分子抗體蛋白的抗原免疫結合活性。結果顯示,scFv-CH2-CH3(scFv-Fc)的結合活性最高。隨后進行攻毒保護實驗,發現該抗體對α毒素感染具有很好的治療作用,且半衰期較長。
此外,自20 世紀90 年代納米抗體被發現以來,其已經成為動物和人類疾病預防和治療的新型小分子抗體藥物[59-60]。納米抗體是衍生自駱駝科動物(單峰駝、駱駝和美洲駝)及鯊魚等血清中天然存在的重鏈(缺乏輕鏈和重鏈1結構域)抗體[61]。與傳統抗體相比,納米抗體具有多個突出的理化性質,如相對分子質量小、穩定性高、靶向性強和水溶性好等,且可在細菌中進行規模化生產,生產成本大大降低[62-63]。盡管具備上述優勢,但納米抗體和常規單抗藥物存在同樣的問題,即口服遞送仍不成功,且費用昂貴。研究發現,微生物載體為納米抗體的口服遞送提供了機會。例如,硅乳桿菌、乳球菌、沙門氏菌和大腸桿菌被工程化后可將靶分子直接遞送至疾病部位,可實現更經濟、更有效的疾病治療。使用載體遞送不僅保護靶分子免受胃腸道環境的影響,還可使其有效性最大化并使脫靶效應最小化[64]。此外,微生物載體還具有制備簡單、成本低等優勢。調查發現,重組乳桿菌已廣泛用作活載體以遞送治療性和預防性分子[65-66]。乳桿菌對CP 有較強拮抗活性,可抑制毒素產生,減少促炎細胞因子,改善腸道病變和免疫應答,糾正微生物生態失調,恢復與亞臨床壞死性腸炎相關的性能缺陷[67-68]。目前,乳桿菌已經用作納米抗體的有效遞送、表達系統,用于不同胃腸道感染性疾病的治療。Dharanesh 等[69]使用重組乳桿菌作為活載體用于原位遞送抗CP NetB 和α 毒素的納米抗體,這些納米抗體中和NetB 和α 毒素協同重組乳桿菌的腸道調節活性,可大大降低家禽壞死性腸炎的發生率。實踐證明,來自載體活菌和納米抗體的雙重作用是解決復雜疾病如壞死性腸炎的突破口,這種治療策略對其他類似復雜疾病的治療潛力也值得探討。
3.2 抗腸毒素疫苗和抗體
由于在小腸內CPE 可誘導大量的上皮細胞脫落和纖毛縮短,各種組織病理損傷可造成相關細胞水分和電介質轉移的改變,進而引起腹瀉癥狀,并且孢子形成的CP 耐熱,煮熟食物幾乎不能降低其致病性[70]。因此,目前與CPE相關的疾病既難預防也無有效治療手段[71]。膜蛋白的緊密連接蛋白家族是已知的CPE 受體,還可控制緊密連接的結構和功能,為細胞間分子輸運制造障礙。CPE 通過緊密連接蛋白與上皮細胞結合,使緊密連接蛋白屏障功能失效,進而誘導細胞毒性,破壞腸道穩態[72]。Hanna 等[73]使用合成抗原結合片段(sFab)文庫發現了2個結合緊密連接蛋白4和CPE C端復合物的合成抗原結合片段,為CPE 相關的胃腸道疾病治療提供了框架和策略。然而,鑒于CPE 感染不常見,目前針對該毒素的疫苗或抗體的研究報道很少,還有待研究。
3.3 抗ε毒素疫苗和抗體
由于ε 毒素致病機制復雜,對人類和動物多個器官均具有較強的毒性作用,目前仍然無針對該毒素的完善的預防或治療策略。Garcia 等[74]報道,用本氏煙草產生的中和單克隆抗體(c4D7)可預防和治療CP ε 毒素中毒。Jiang 等[75]報道,ETX-Y71A可作為CP 的無毒突變體,在小鼠和綿羊中誘導保護性免疫。但上述方案尚未得到臨床驗證。
3.4 抗體-抗生素偶聯藥物策略
除上述抗CP治療性抗體(表1)外,為提高對耐藥細菌的治療效果,借助抗體偶聯藥物的設計原理,研究人員還提出了一種用于治療細菌感染的抗體-抗生素偶聯藥物策略,其主要結構包括可特異性靶向識別細菌的抗體以及用于殺死細菌的抗生素有效載荷[76]。研究發現,數千個抗體-抗生素偶聯藥物可與單個細菌結合,并可被吞噬至細菌體內釋放出足夠濃度的游離抗生素[77],以此提高藥物的有效性(降低藥物劑量和不良反應),從而降低耐藥性。抗體-抗生素偶聯藥物策略通過將抗體和抗生素的藥理學屬性結合到單個藥物中,為感染性疾病的防治提供全新的治療手段。盡管目前還未開發出針對CP 感染的抗體-抗生素偶聯藥物,但這些非臨床研究案例將大大促進治療CP 感染的抗體-抗生素偶聯藥物的發展,在一定程度上解決CP 耐藥問題。

表1 抗產氣莢膜梭菌治療性抗體
4 結語
CP 感染是自然界現存最具危害性的革蘭陽性細菌之一。因此,針對CP 做好科學有效的預防策略,不僅能夠提高動物生產數量和質量,也能消除其給人類健康帶來的威脅。面對CP 抗生素治療后耐藥性激增,預防性疫苗、治療性抗體以及抗體-抗生素偶聯藥物可在一定程度上阻止其耐藥進程。目前,針對預防和治療CP 感染的疫苗和抗體逐漸成為大家關注和研究的重點,無論是單鏈抗體、雙價單鏈抗體,其結合活性、中和活性以及保護活性均有待提高。更重要的是,對具有重大臨床應用價值的候選藥物,需開展進一步的非臨床和臨床研究,以滿足臨床上對CP 感染治療的需求。相信隨著生物科技的迅猛發展,在應對感染性疾病方面不再局限于傳統的抗生素療法,越來越多的新型生物療法將不斷涌現,為人類對抗“頑固性耐藥菌”感染提供更多有效的措施。
