落葉松八齒小蠹伴生長喙殼真菌致病性研究
2024-04-14 12:51:36周勤政王慧敏
林業科學研究 2024年2期
劉 亞,周勤政,王慧敏,王 正,呂 全*
(1. 國家林業和草原局森林保護學重點實驗室 中國林業科學研究院森林生態環境與自然保護研究所,北京 100091;2. 中國農業大學植物保護學院,北京 100193;3. 山東農業大學植物保護學院,山東 泰安 271018)
落葉松八齒小蠹(Ips subelongatusMotschulsky)為鞘翅目、小蠹亞科、齒小蠹屬昆蟲,在東亞廣泛分布,是中國北方3 種落葉松(興安落葉松、華北落葉松和長白落葉松)的次期性嚴重為害的蛀干害蟲[1],在種群暴發期可為害健康樹木。作為影響本土針葉樹并對非本土針葉樹有威脅的主要害蟲,該害蟲于2003 年被中國列入“全國林業危險性有害生物名單”(國家林業局,2003年),于2005 年被歐洲地中海植物保護組織列入A2 預警名單(EPPO,2005)。此外,小蠹蟲攜帶的多種病原微生物,特別是長喙殼真菌,在小蠹蟲為害過程中會加重對樹木的危害[2]。
長喙殼真菌是一類包括蛇口殼目(Ophiostomatales)真菌及相似類群真菌的總稱,其中一些種類是引起林木病害的重要病原菌,如引起榆樹荷蘭病的Ophiostoma ulmi和Ophiostoma novo-ulmi、引起月桂枯萎病的Raffaelea lauricola及引起針葉樹黑根病的Leptographium wageneri[2-4]。已有研究證實,齒小蠹伴生真菌群落中的先鋒種Endoconidiophoraspp.,如Endoconidiophora fujiensis,對日本地區的落葉松毒力較強,可引發寄主壞死,或誘導寄主落葉松產生強烈的抗性反應[5]。Endoconidiophora polonica高密度接種在挪威云杉上,能導致其死亡[6]。
小蠹蟲和長喙殼真菌在長期的協同進化過程中形成了復雜的伴生關系,二者之間的聯系在小蠹蟲定殖寄主過程中起著重要作用[7-8]。研究表明,長喙殼真菌可以合成小蠹蟲生長發育所必需的甾醇[9]、解毒寄主樹木的防御物質[10-12]、釋放化學信息素吸引或排斥同種小蠹蟲[13-14]等。然而,一些寄主萜類化合物對小蠹蟲及其伴生真菌具有毒性,如針葉樹在受到小蠹蟲攻擊時會增加單萜類化合物的合成,以抵抗小蠹蟲的入侵,當萜類物質濃度高于小蠹蟲的生理耐受閾值時,抑制小蠹蟲的攻擊[15];離體試驗也表明,在培養基中添加α-pinene 和limonene 等單萜類物質,可抑制長喙殼真菌的生長[14, 16]。
研究明確小蠹蟲傳播的真菌是否會引起植物病害以及植物是否會對小蠹蟲及其伴生真菌產生抗性,是關于小蠹蟲傳播病菌的2 個關鍵科學問題。本研究將2010—2018 年采集分離的12 種落葉松八齒小蠹伴生長喙殼真菌代表性菌株接種到5 年生落葉松苗上[17],觀察不同真菌或菌株之間的致病力是否存在差異,并分析寄主防御相關代謝物質的變化,以探究落葉松八齒小蠹及其伴生長喙殼真菌復合物對落葉松的潛在風險。
1 材料與方法
1.1 試驗材料
1.1.1 供試菌株 本試驗選用的12 種長喙殼真菌為由2010—2018 年采集的落葉松八齒小蠹蟲體和坑道中分離純化得到的菌株[17],分別為E.fujiensis、E.laricicola、Ophiostoma hongxingense、Ophiostoma peniculi、Ophiostoma pseudobicolor、Ophiostoma subelongati、Ophiostoma genhense、Ophiostoma lotiforme、Ophiostoma rufum、Ophiostoma minus、Ophiostoma multisynnematum和Ophiostoma xinganense,每種真菌各選用2 個菌株用于致病力試驗(表1)。

表1 供試菌株信息 Table 1 The information of tested strains
1.1.2 寄主植物 選擇移栽于中國林科院內的5 年生長白落葉松(Larix olgensisA. Henry)作為試驗材料。這些落葉松高2.0~3.0 m,平均2.49 m,地徑2.05~4.77 cm,平均3.05 cm。
1.2 試驗方法
1.2.1 接種 菌株在2% MEA 培養基上25 ℃黑暗條件下培養7 d,用經滅菌的直徑6 mm 軟木塞打孔器在新培養菌落邊緣打取菌苔。于2019 年7 月9 日隨機選擇60 棵落葉松,在離地30 cm 處的樹干上,用經滅菌的直徑6 mm 的軟木塞打孔器打孔至木質部,用無菌牙簽將菌苔接入接種孔,然后蓋上樹皮,用封口膜纏繞,并用膠帶固定。以2% MEA 培養基作為接種對照。每棵樹上設1 個接種點,每個處理各設5 次重復。
1.2.2 接種發病調查 接種60 d 后,撕開接種點的膠帶和封口膜,用壁紙刀刮去接種點周圍病斑區域的外樹皮,測量病斑的長和寬,并記錄相關數據。
1.2.3 病原菌的再次分離 測量病斑大小后,每個處理隨機選擇3 個重復,在接種植株病健交界處取約1 cm2樣品放至15 mL 無菌離心管并帶回實驗室,于-4 ℃冰箱保存。次日,樣品表面經2%次氯酸鈉消毒1 min,然后用無菌水沖洗3 次,經無菌濾紙吸去表面水分后取約1 mm2組織碎片放置在2% MEA 培養基上進行培養,根據菌落形態學和核酸β-微管蛋白(BT)基因片段序列比較鑒定是否為原始接種的長喙殼菌株。
1.2.4 單萜類物質測定 對MJG55 (E.fujiensis)和TRL63(E.laricicola)菌株和對照接種1、7、14、30 和60 d 后的5 棵落葉松進行單萜類物質測定,以評估寄主防御反應強弱。用聚乙烯薄膜(48 cm × 60 cm,雷諾公司,美國)包裹接種點周圍樹干,用特氟龍管將含有100 mg Porapak-Q 吸附劑(默克公司,德國)的玻璃管連接在小型真空泵上形成閉環動態頂空采樣系統,以500 mL·min-1的流速持續1 h 收集寄主揮發性物質。將含有揮發物的玻璃管兩端用錫紙包裹并纏繞封口膜,放入裝有干冰的保溫箱并帶回實驗室。用2 mL 色譜級正己烷洗脫寄主揮發物,用溫和的氮氣將樣品濃縮至50 μL,置于-20 ℃冰箱保存直至氣相色譜檢測。氣相色譜分析方法同Liu 等人描述[18]。將樣品放置在配有火焰離子檢測器和自動進樣器的氣相色譜儀(GC-FID,安捷倫,美國)上,使用口徑0.25 mm × 0.2 μm,30 m 長的HP-5 色譜柱進行分析,每次進樣1 μL。
1.3 數據處理
根據Rajtar 等[19]的方法,計算接種點病斑面積(長 × 寬)。單萜類化合物釋放速率根據處理組和健康組的差值(Δ)進行分析。使用IBM SPSS Statistics19 針對不同菌株產生的病斑、MJG55 和TRL63 菌株處理的落葉松在同一時間產生的單萜類化合物釋放速率、MJG55 或TRL63 菌株處理在不同時間產生的單萜類化合物釋放速率,進行單因素方差分析(LSD test,α=0.05)。
2 結果與分析
2.1 接種不同菌株后落葉松發病情況
接種兩個月后的落葉松沒有表現出明顯的病害癥狀和枯萎現象。剝開外層樹皮,不同長喙殼菌株接種落葉松后產生的褐色至黑色壞死病斑如圖1,病斑面積大小見圖2。比較發現,E.laricicola的2 個菌株(TRL63 和KBLK9)產生的病斑最大,面積分別為3.51 ± 0.18 cm2和2.89 ± 0.19 cm2。單因素方差分析結果顯示E.laricicola產生的病斑顯著大于其他處理(圖2)。E.fujiensis的2 個菌株(MJG55 和MJG29)、O.hongxingense的2 個菌株(HXS70 和HXS72)、O.peniculi的GH77 菌株和O.xinganense的CFCC52680 菌株產生的病斑顯著大于對照處理(1.28 ± 0.15 cm2)。其余菌株接種產生的病斑與對照無差異,且接種點中間未呈現壞死狀態(圖2)。

圖1 落葉松接種長喙殼真菌后產生的病斑Fig. 1 The lesions produced on larch after inoculation with ophiostomatoid fungi
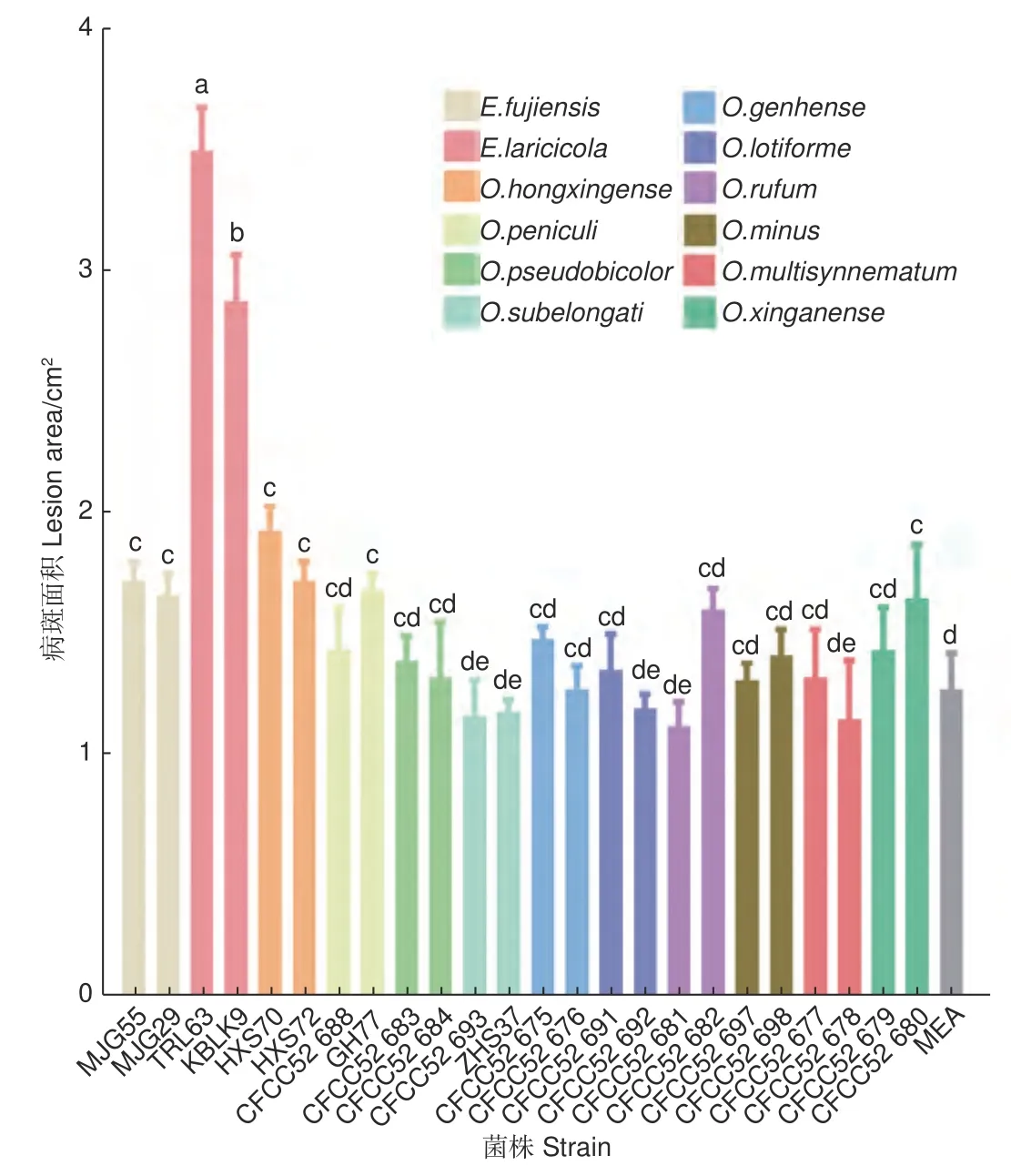
圖2 落葉松接種長喙殼真菌后產生的病斑大小Fig. 2 Size of lesions produced on larch inoculated with ophiostomatoid fungi
2.2 菌株接種后病菌再分離
對人工接種產生病斑的病菌再分離和鑒定,得到每個菌株處理的3 個隨機病變樣本平均再分離率為83.3%,證實了落葉松接種點處產生的壞死病斑是由原始接種的長喙殼真菌引起。
2.3 萜類測定結果
為了研究落葉松對長喙殼真菌的抗性反應,針對接種了E.fujiensis(MJG55)和E.laricicola(TRL63)菌株的落葉松,在接種后的1、7、14、30 和60 d 分別進行了單萜類物質測定。共檢測到(-)-α-pinene、camphene、β-pinene、myrcene、3-carene 和(-)-limonene 6 種單萜物質,結果如圖3 所示。總體而言,TRL63 處理的單萜釋放速率最高,MJG55 處理次之,這兩種處理與對照均存在顯著差異。不同處理后單萜類物質釋放速率隨時間的變化規律略有不同,TRL63 處理中各萜類物質呈現先升高后降低的變化趨勢;MJG55 和MEA 處理的myrcene 也呈現先升高后降低的變化趨勢,(-)-α-pinene、camphene 和3-carene 呈現下降的趨勢。
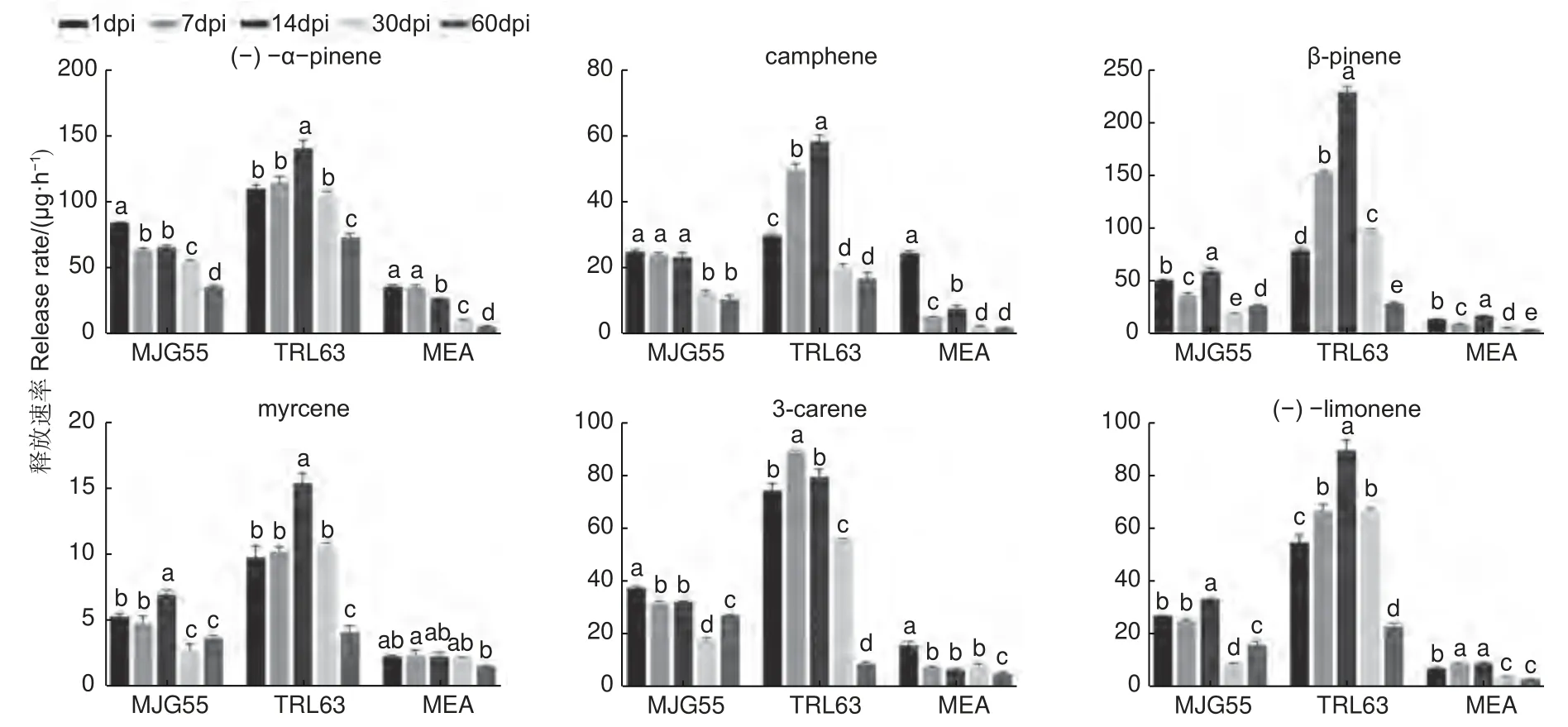
圖3 不同處理下落葉松的單萜釋放速率Fig. 3 Release rates of six monoterpenes by larch in different treatments
3 討論
真菌致病性是指真菌侵染活體植物組織后在其體內存活和發育并引起病害的能力[20-21]。致病性用毒力來量化,毒力是通過測量寄主可視化癥狀的發展來確定的,包括病斑長度、邊材干燥和藍變面積、樹脂反應以及樹木的死亡率[22-23]。病斑指寄主植物感染點周圍韌皮部組織產生的變暗、凸起或輕微凹陷以及壞死等現象,用于評估真菌對寄主樹的影響[24]。長喙殼真菌的致病性一般是通過在成年樹或幼苗上人工接種真菌來評估[6,25-26]。盡管在成年樹上接種更接近小蠹蟲高密度攻擊后的自然接種,但幼苗接種是控制實驗環境條件的另一選擇[22,24]。此外,在接種量相當的情況下,幼苗接種更適合于確定小蠹蟲伴生長喙殼真菌的毒力[22]。因此,本研究選用5 年生的落葉松苗木驗證12 種落葉松八齒小蠹伴生長喙殼真菌的致病性。結果表明,5 種真菌對長白落葉松表現出不同程度的致病力,E.laricicola致病力最強,E.fujiensis和O.hongxingense次之,O.peniculi和O.xinganense較弱(圖2),表明這幾種真菌對落葉松存在潛在危害風險。其余7 種Ophiostoma真菌在接種點處形成的韌皮部變色與對照無差異,未表現出明顯致病性。
本研究從亞洲落葉松八齒小蠹蟲體上分離到E.laricicola真菌,與之前關于E.laricicola和歐洲落葉松八齒小蠹(Ips cembrae)特異性伴生的認識存在差異[27],這可能是小蠹蟲和真菌在擴散中形成了新的伴生關系。Redfern 等人最早發現I.cembrae與E.laricicola存在伴生關系,且由這種伴生關系導致的復合侵染使落葉松發生頂梢枯死[28]。E.laricicola能在歐洲赤松韌皮部形成壞死病斑,誘導木質素和樹脂的積累,同時減少歐洲赤松細胞壁游離原花青素、低分子量碳水化合物和非木質素成分的含量,表明其能抑制寄主傷口反應早期階段的木質化,從而延緩寄主對于損傷的自我修復[29]。本研究中,E.laricicola接種在落葉松韌皮部形成的壞死病斑最大,其誘導寄主產生的萜類物質呈現先升后降的趨勢,表明該菌具有較強致病力。因此,E.laricicola與亞洲落葉松八齒小蠹形成的伴生關系可能更有利于小蠹蟲的入侵,對落葉松造成較強的破壞力。
以前有研究表明,在人工接種條件下E.fujiensis能使30 年生的健康日本落葉松死亡[5]。該菌對長白落葉松也表現出一定致病力,并能引起長白落葉松的防御反應[30]。本研究發現,E.fujiensis菌株接種后落葉松韌皮部形成的壞死病斑顯著大于對照處理,但與E.laricicola相比病斑較小,在5 個調查時間段內大多數萜類物質的變化也較小。這與致病力更強的真菌誘導宿主防御萜類物質發生較大變化的結論一致[16,18,31],說明E.fujiensis菌株對長白落葉松的致病力較弱。本文研究結果與Yamaoka[5]和周秀華等[30]的研究結果不同,可能與樹木的個體發育有關。有研究發現,與成年樹相比,幼樹具有更強的化學防御能力,在抵抗病原菌侵染時產生更強烈的抗性反應[32-33]。因此,本研究在5 年生落葉松上設定的單一接種量有限,可能不足以造成嚴重影響,以后將考慮增加接種量。
長喙殼真菌入侵寄主后,寄主體內防御性萜類物質釋放增加,這些萜類化合物影響真菌的生長,同時真菌也具有解毒能力,甚至利用寄主產生的防御性化合物。Fang 等人研究發現,α-pinene、limonene 和carene 等8 種單萜類物質會抑制E.fujiensis的生長[16]。Cale 等人發現,在額外添加α-pinene 的培養基上山松大小蠹伴生菌Grosmannia clavigera生長也受抑制[31];而limonene 的存在會促進G.clavigera和Ophiostoma ips真菌的生長。此外,不同濃度的萜類對真菌生長的影響也存在差異,如低濃度的myrcene 能促進O.ips的生長,而高濃度myrcene 對O.ips生長發育無影響[30]。Liu 等人認為長喙殼真菌能夠利用寄主揮發物中的氮能來促進真菌的生長發育并合成小蠹蟲生長發育所必需的麥角甾醇[34]。小蠹蟲和長喙殼真菌復合侵染對寄主造成的危害遠高于小蠹蟲自身鉆蛀樹木造成的危害[35],因此,研究小蠹蟲和伴生真菌的內在聯系以及伴生真菌對寄主防御的影響,對于防治小蠹蟲暴發危害具有重要理論指導意義。
4 結論
本研究通過人工接種試驗發現,小蠹蟲伴生長喙殼真菌的不同菌株在落葉松接種點處產生的病斑大小存在差異,有5 種長喙殼真菌對長白落葉松表現出不同程度的致病力,E.laricicola菌株致病力最強,E.fujiensis和O.hongxingense次之,O.peniculi和O.xinganense較弱。Endoconidiophora真菌能夠誘導寄主產生防御反應,釋放的萜類物質增加,且在E.laricicola接種14 d 左右達到峰值。長喙殼真菌被小蠹蟲攜帶至新的寄主植物上,在小蠹蟲坑道周圍產生的壞死病斑與寄主植物的死亡存在密切關系。本研究通過人工接種長喙殼真菌后測量病斑大小來評估真菌致病力,對于優化小蠹蟲綜合治理策略提供理論依據與技術支撐。
