PagKNAT2/6a 與PagKNAT2/6b 在木質部發育中存在功能保守性
2024-04-14 12:51:22李海陽趙立子趙巖秋
林業科學研究 2024年2期
關鍵詞:研究
李海陽,趙立子*,趙巖秋
(1. 魯東大學農林工程研究院,山東 煙臺, 264025;2. 魯東大學農學院,山東 煙臺, 264025)
木材是木本植物次生木質部長年累積的產物,次生木質部發育是一個動態過程,包括維管形成層分裂、木質部與韌皮部細胞分化、次生細胞壁形成等各個階段[1]。其中,木質部細胞的分化是木材形成的關鍵。木本植物中楊樹(Populus)憑借其基因組小、適應性強、生長速度快、易于無性繁殖等特點,已被公認是林木分子生物學研究的模式物種[2]。近年來,隨著對木質部發育相關研究的逐步深入,已有大量研究表明楊樹木質部發育受多種遺傳和內源因子調控,如激素、轉錄因子及多肽信號等[3]。其中,轉錄因子作為調控木質部發育的關鍵因子存在,已有大量研究報道轉錄因子參與調控木質部發育,如:XND1(XYLEM NAC DOMAIN 1)負調控木質部導管分化[4];油菜素內酯對木質部分化起到正向調控作用,且有利于促進應力木的形成[5];生長調節因子PagGRF12a被顯性抑制可有效增加木質部寬度,促進木質部發育[6]。綜上可見,木質部發育受復雜的轉錄因子調控,從而影響木材形成過程。
KNOX 家族是一類重要的同源異型盒基因,廣泛存在并參與生物的發育進程,并通過調控植物分生組織影響不同組織、器官的發育[7]。擬南芥(Arabidopsis thaliana)中,I 類KNOX 包括4 個 成 員 分 別 是 :STM(SHOOT MERISTEMLESS)、KNAT1(KNOTED1-LIKE FROM ARABIDOPSIS THALIANA1)、KNAT2和KNAT6。已有研究表明KNAT2與KNAT6分別在莖頂端分生組織的底部與邊緣處表達[8];與此同時,本研究團隊前期對‘84K’楊(Populus alba×Populus glandulosa)中KNAT2和KNAT6的同源基因PagKNAT2/6a與PagKNAT2/6b的功能展開研究,已發現PagKNAT2/6b通過直接激活XND1a的表達抑制木質部分化及次生細胞壁的合成[9],但PagKNAT2/6a作為與PagKNAT2/6b進化關系最近的成員,其在楊樹木質部發育中的調控功能未知。
本研究為探究PagKNAT2/6a在木質部發育中的功能,以PagKNAT2/6a為研究對象,通過‘84K ’ 楊樹遺傳轉化技術獲得異常表達PagKNAT2/6a轉基因株系,借助GUS 染色實驗和實時熒光定量(qRT-PCR ) 比較分析PagKNAT2/6a與PagKNAT2/6b組織表達特異性,同時借助莖部解剖學分析其異常表達PagKNAT2/6a轉基因株系莖中木質部分化差異,通過比較分析PagKNAT2/6a與PagKNAT2/6b在楊樹木質部中的表達定位及木質部分化中的功能差異發現該基因在木質部發育中可能存在的功能保守性。本研究對PagKNAT2/6a與PagKNAT2/6b在楊樹木質部發育中功能保守性的解析,以期為KNOX 類基因在木本植物木質部發育上的保守性和分化性提供理論支持,并為KNOX 基因在定向分子設計育種工作開展中提供重要的基因資源。
1 材料與方法
1.1 氨基酸序列比對
通過NCBI中的BLAST(https://blast.ncbi.nlm.nih.gov)檢索‘84K’楊樹(Populus alba×Populus gllandulosa)中PagKNAT2/6a與PagKNAT2/6b基因序列,然后利用MEGA 軟件中Clustal Co 程序進行多序列比對。
1.2 GUS 染色
取生長3~4 周的PPagKNAT2/6ba::GUS 組培苗整株浸泡于固定液(90%丙酮)中,4 ℃固定過夜;固定后的材料用配置好的GUS 染色緩沖液(0.2 mol·L-1磷酸二氫鈉,0.2 mol·L-1磷酸氫二鈉,2 mmol·L-1鐵氰化鉀,2 mmol·L-1亞鐵氰化鉀,現用現配)在冰上洗滌3 次,然后將材料轉移至GUS 染色液( 1 mmol·L-1x-Gluc 先溶于DMF,后溶于染色緩沖液,并加0.2%的TritonX-100,現配現用)中,抽真空20~30 min,后置于37 ℃搖床70 r·min-1,染色10 h。最后用75%乙醇進行脫色,取脫色后的莖或葉片,將其固定于5%~6%瓊脂中,待凝固后使用振動式切片機切片,后使用顯微鏡對GUS 信號的定位進行觀察。GUS 染色實驗包含2 個轉基因株系,每個轉基因株系設置單株重復。
1.3 RNA 提取及qRT-PCR 分析下游基因表達量
采用華越洋植物RNA 提取試劑盒,分別提取‘84K’楊,過表達株系(OE6,OE18)的相同節間總RNA,然后以RNA 為模板按照 HiScript II Q RT SuperMix for Qpcr(+ gDNA wiper)試劑盒說明書合成cDNA,并利用 ChamQ Universal SYBR qPCR Master Mix 試劑盒進行qRT-PCR(反應體系:SYBR Premix Ex Taq Tm 5 μL,引物各0.5 μL,cDNA 1 μL,ddH2O 3 μL),以Actin作為內參基因,每個樣本設4 個生物學重復。最后采用2-△△CT法計算莖植物組織相對表達量。同時,本研究選擇了細胞壁三大素(纖維素,半纖維素,木質素)部分合成相關的基因(CESA4、CESA7A、CESA7B、CESA8A、CESA8B、GUX1a、GUX1b、PARVUS、4CL、C3H、COMT、CAD、LAC4、LAC17)通過qRT-PCR技術進行分析PagKNAT2/6a可能的下游靶基因,所有引物見表1。
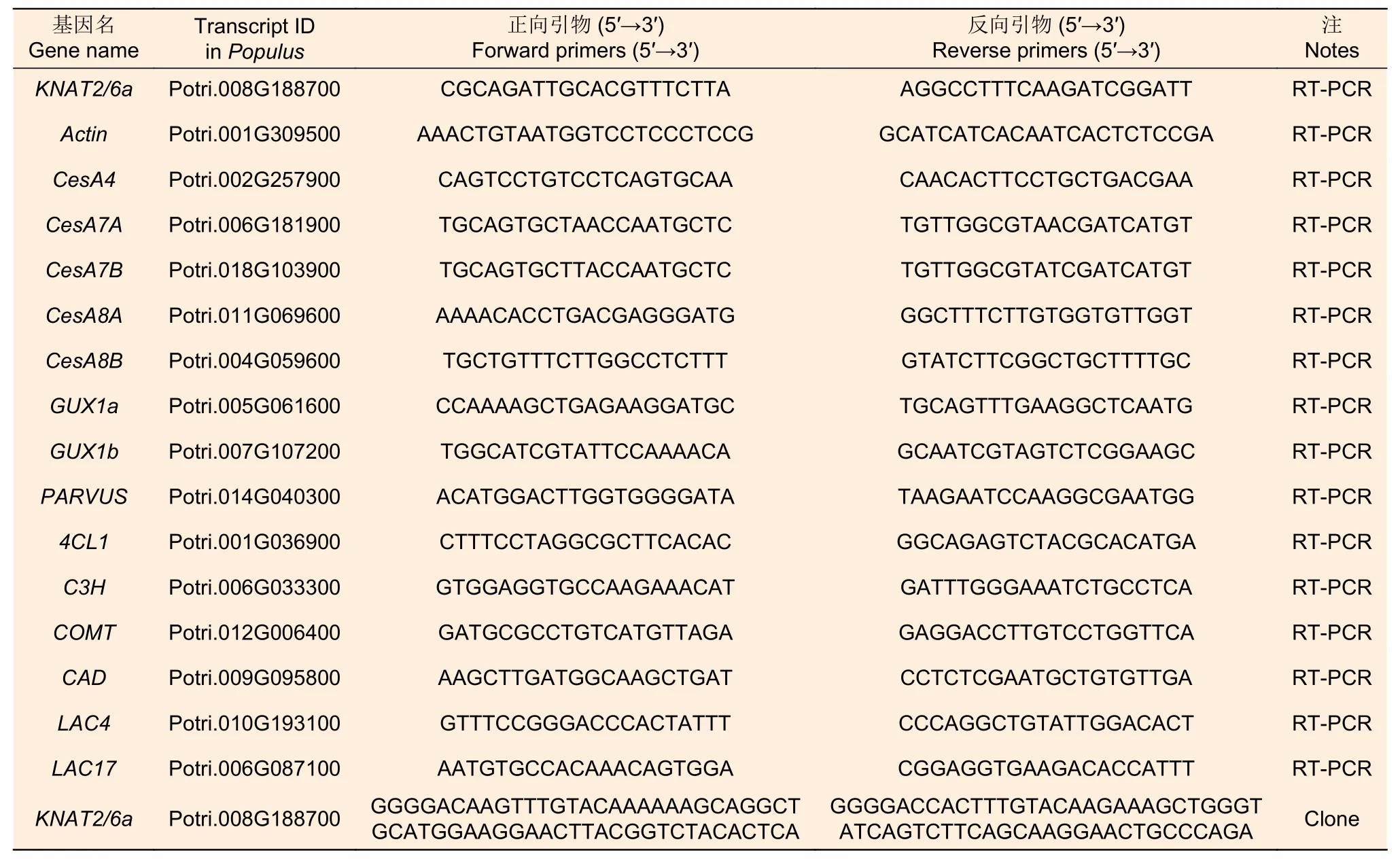
表1 本研究中所用的引物序列 Table 1 The primer sequences used in this study.
1.4 震蕩切片與TBO 染色
將新鮮楊樹莖的節間切成1 cm 長,一端用膠水固定于樣品架,利用震蕩切片機對樣品進行切片,切片厚度一般設置為60 μm,將切片收集至ddH2O 中,待染色觀察。染色時直接將染色液(0.5% 甲苯胺藍)滴到切片上,1 min 左右后用蒸餾水沖洗在熒光共聚焦顯微鏡下觀察并拍照。
1.5 掃描電鏡分析
用雙面刀片迅速取下野生型及PagKNAT2/6a異常表達的轉基因植株新鮮的莖段,后將其置于裝有4%多聚甲醛中固定2 h,后經過脫水,臨界點干燥,將莖段垂直放置于樣品臺上,噴金處理使莖段橫切面均勻噴上金粉,最后用掃描電鏡觀察并拍照。
2 結果與分析
2.1 PagKNAT2/6a 與PagKNAT2/6b 氨基酸序列高度保守
根據PagKNAT2/6a(Pop_A08G045880)與PagKNAT2/6b(Pop_A10G047641)進化關系, 本研究首先分析了PagKNAT2/6a與PagKNAT2/6b氨基酸序列的相似性,如圖1 所示,PagKNAT2/6a與PagKNAT2/6b氨基酸序列高度相似,且都包含HOMEBOX-1 保守的同源盒,表明PagKNAT2/6a與PagKNAT2/6b基因同源性相對較高,序列相對保守,為此推測PagKNAT2/6a與PagKNAT2/6b基因在楊樹發育中可能存在功能保守性。
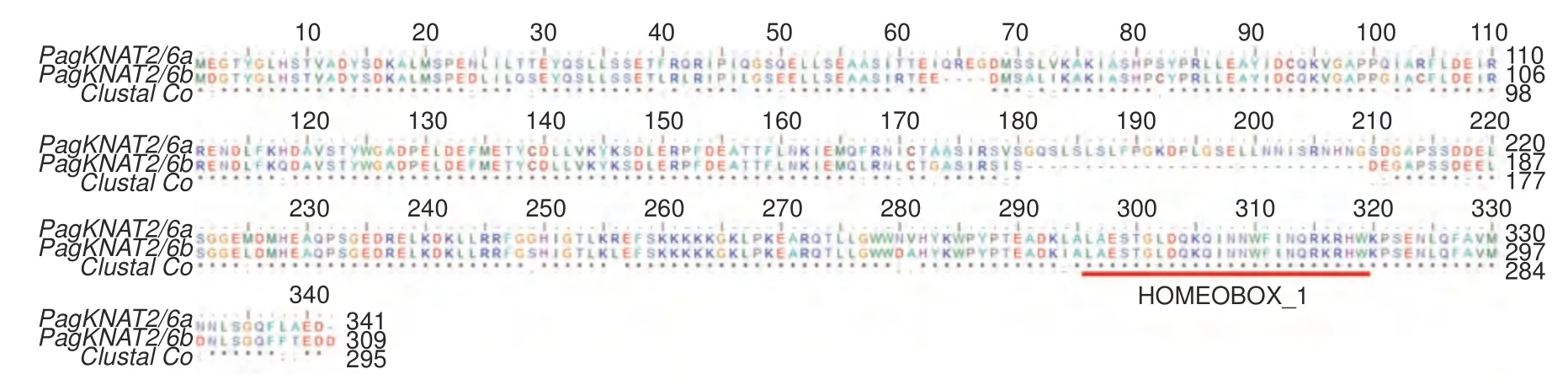
圖1 PagKNAT2/6a 與PagKNAT2/6b 氨基酸序列比對Fig. 1 PagKNAT2/6a and PagKNAT2/6b amino acid sequence alignment.
2.2 PagKNAT2/6a 與PagKNAT2/6b 具有相似的表達模式
為比較分析PagKNAT2/6a與PagKNAT2/6b在楊樹不同組織中的表達特異性,本研究首先借助實時熒光定量PCR 技術,檢測PagKNAT2/6a與PagKNAT2/6b基因在‘84K’楊樹根、根尖、莖、莖尖、幼嫩葉及成熟葉6 個部位中的組織表達變化(圖2A),結果顯示PagKNAT2/6a與PagKNAT2/6b基因均在楊樹莖、莖尖以及幼嫩葉片中表現出較高的表達量。與此同時,本研究克隆了PagKNAT2/6a與PagKNAT2/6b基因啟動子序列,將其融合至含有GUS 報告基因的載體中,通過穩定的楊樹遺傳轉化方法獲得PPagKNAT2/6a::GUS 與PPagKNAT2/6b::GUS轉基因楊樹并進行GUS 染色實驗分析其表達定位(圖2B 與2C)。結果顯示PagKNAT2/6b基因(圖2B)在幼嫩節間與葉片中GUS 信號強于PagKNAT2/6a(圖2C),且在GUS 信號觀察中發現PagKNAT2/6a 與PagKNAT2/6b在幼嫩葉片與幼嫩莖段中的表達均強于成熟葉片與根。

圖2 PagKNAT2/6a 與PagKNAT2/6b 基因組織表達特異性分析Fig. 2 Tissue expression specificity analysis of PagKNAT2/6a and PagKNAT2/6b genes
隨后, 為深入研究PagKNAT2/6a與PagKNAT2/6b基因在莖中的表達特性,通過GUS 染色放大圖發現PagKNAT2/6a 與PagKNAT2/6b均特異在木質部與韌皮部表達(圖3A、C),但利用Popgenie 網站分析發現PagKNAT2/6b表達量在木質部表達量顯著高于PagKNAT2/6a(圖3B、3D)。根據上述結果推測,楊樹PagKNAT2/6a與PagKNAT2/6b基因可能均參與楊樹莖部木質部發育方面的調控。

圖3 PagKNAT2/6a 與PagKNAT2/6b 基因在莖中高表達Fig. 3 PagKNAT2/6a and PagKNAT2/6b genes are highly expressed in stems
2.3 PagKNAT2/6a 異常表達影響木質部分化與次生細胞壁合成
基于PagKNAT2/6a與PagKNAT2/6b在楊樹莖中木質部區域特異性表達,為深入比較分析PagKNAT2/6a與PagKNAT2/6b在木質部發育中的功能,本研究在前期分別創制了PagKNAT2/6a過量表達轉基因楊樹(PagKNAT2/6aOE,OE6與OE18)和PagKNAT2/6a抑制表達轉基因楊樹(PagKNAT2/6aSRDX,DR3 與DR15)轉基因株系(已發表)。與此同時,對生長一個月的不同轉基因株系的第八節間分別進行切片(圖4A)及掃描電鏡分析(圖4B)。結果顯示:PagKNAT2/6aOE 株系的木質部寬度和次生壁沉積厚度顯著窄于野生型(CK),而PagKNAT2/6aSRDX 株系莖中木質部顯著寬于CK。該結果與研究團隊前期發現的PagKNAT2/6b基因對木質部的影響相似,說明PagKNAT2/6a與PagKNAT2/6b在木質部分化調控上的保守性。

圖4 PagKNAT2/6a 異常表達影響莖中木質部分化與木質部細胞壁厚度Fig. 4 Abnormal expression of PagKNAT2/6a affects woody partialization and xylem cell wall thickness in stems
2.4 PagKNAT2/6a 調控壁合成相關轉錄因子
眾所周知,次生細胞壁是木質部的主要組成部分,其組成成分對木材品質有直接的影響,基于PagKNAT2/6b基因抑制次生壁合成相關基因的表達從而抑制楊樹莖中木質部分化的研究基礎[10];為此,推測PagKNAT2/6a基因可能通過調控次生壁合成基因來影響木質部的發育。為進一步解釋PagKNAT2/6a與PagKNAT2/6b異常表達與次生壁合成相關基因之間的調控關系,本研究通過qRT-PCR 分析了細胞壁三大素(纖維素、半纖維素、木質素)合成相關基因在CK、PagKNAT2/6aOE 植株莖中的表達變化(圖5)。結果分析發現:相比于CK,參與調控次生壁合成的纖維素、木糖(半纖維素中主要組分)、木質素等相關基因(COMT、CAD、LAC4、LAC17、CesA4、CesA7A、CesA7B、CesA8A、CesA8B)在PagKNAT2/6aOE 莖中均表現出下調表達的趨勢,其中,纖維素合成相關的CesA類基因的下調趨勢最顯著(圖5C)。根據以上結果可知,PagKNAT2/6a基因通過調控次生壁合成相關基因的表達影響木質部發育,且PagKNAT2/6a與PagKNAT2/6b在對次生壁合成相關基因的調控上具有保守性。

圖5 PagKNAT2/6a 異常表達影響細胞壁合成相關基因的表達Fig. 5 Aberrant expression of PagKNAT2/6a affects the expression of genes related to cell wall synthesis
3 討論
次生木質部由維管形成層細胞通過平周、垂周分裂不斷增殖,位于內側的細胞不斷分化,伴隨次生壁形成和細胞內容物溶解從而形成成熟的次生木質部[11]。次生木質部發育是木材生產的生物學基礎,其發育調控原理一直是林木分子生物學基礎研究中關注的核心科學問題。已有研究表明,木質部發育受多種因子調控,如細胞比例,次生細胞壁的結構組成,植物激素和關鍵基因等。目前,次生壁合成被認為由三級調控網絡調控,調控木質部細胞分化的VND 類轉錄因子,其可直接調控第二級轉錄因子MYB46、MYB83的表達,后者又作為重要開關,調控細胞壁三大素合成相關酶基因的表達[12]。與此同時,已有研究發現KNOX 基因參與調控木本植物形成層分化,其中楊樹中Ⅰ類KNOX 成員ARK1與ARK2均在形成層區域表達[13-14],而PagKNAT2/6a與PagKNAT2/6b在形成層兩側表達,可見不同KNOX 類成員分別影響形成層的分裂及子細胞的分化過程[15]。
木材來自形成層的活動,調控形成層活動的基因會影響木質部的發育,楊樹中PagKNAT2/6a與PagKNAT2/6b均在形成層兩側區域有較高的表達量,分析PagKNAT2/6b在木質部發育中的功能,發現PagKNAT2/6b過表達會使木質部分化受到抑制[9]。本研究著重比較了PagKNAT2/6a與PagKNAT2/6b在對楊樹莖中木質部的發育調控上的異同,結果發現其在木質部發育中的功能存在一定保守性,但從莖部切片結果可知(圖4A),PagKNAT2/6b對木質部發育的影響大于PagKNAT2/6a。
4 結論
本研究在探究PagKNAT2/6a基因在木質部發育中的功能,同時對楊樹PagKNAT2/6a與PagKNAT2/6b在木質部發育中功能的保守性進行分析,確定了PagKNAT2/6a與PagKNAT2/6b在木質部發育與次生壁合成中的功能保守性,本研究不僅為木材形成調控網絡建立提供有效線索,而且為木本植物分子育種工作提供理論基礎。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
