過氧自由基氧化對花生球蛋白結構及聚集行為的影響
2024-04-12 09:38:46汪海燕李玟君李玉蝶宋青云龐子皓肖一郎汪超李瑋
食品研究與開發 2024年7期
汪海燕,李玟君,李玉蝶,宋青云,龐子皓,肖一郎,汪超,李瑋
(湖北工業大學 生物工程與食品學院,湖北 武漢 430000)
花生蛋白由90%的鹽溶性蛋白和10%的水溶性蛋白組成,而鹽溶蛋白中含有73% 的花生球蛋白[1]。花生中的球蛋白具有良好的熱穩定性、復雜的空間構象以及維持空間構象所需要的疏水鍵、二硫鍵和離子鍵等功能性官能團,可作為一種優良原料廣泛應用于食品領域[2]。花生球蛋白是花生蛋白中最重要的組分,直接影響花生蛋白的結構和理化性質。因此花生球蛋白更適于花生蛋白的結構和理化性質改性機理的深入研究。
花生在日常貯藏和加工過程中,容易發生脂質氧化反應,導致產品保質期縮短并產生異味,影響食用價值。蛋白質經氧化后羰基化合物含量增加,蛋白的分子間作用力被破壞,高級結構改變并形成交聯聚集,因此通過研究蛋白質的理化性質、光譜特征、反應位點等可反映氧化對蛋白質的作用規律[3]。已有研究人員使用了各種活性氧或脂質過氧化的副產物來模擬蛋白質氧化,發現蛋白質的氧化會影響食物蛋白質的結構和功能特性,例如肉蛋白[4]和大豆蛋白[5]。另外李志豪[6]研究2,2′-鹽酸脒基丙烷對瓜籽蛋白的影響發現,在低氧化度(0~1 mmol/L)時,籽瓜種仁分離蛋白質產生可溶性聚集體。隨著蛋白質氧化程度的增加,共價鍵交聯可能導致這些可溶性聚集體進一步聚集,產生不溶性聚集體,導致籽瓜種仁分離蛋白質溶解度降低。
目前,對于花生球蛋白的改性研究主要集中在物理研究方面,如熱處理和高壓處理[7],而對花生球蛋白氧化的研究較少。2,2′-鹽酸脒基丙烷[2,2′-azobis(2-amidinopropane),AAPH]有氧熱分解產生的過氧自由基是脂質過氧化反應中最主要的自由基中間體[8]。因此,本研究采用AAPH 在有氧條件下熱分解產生的過氧自由基代表花生蛋白氧化酸敗中產生的脂質自由基,研究脂質自由基氧化對花生球蛋白結構的影響,以期更深入地了解花生蛋白的氧化機理,為合理開發花生蛋白提供參考。
1 材料與方法
1.1 材料與試劑
花生蛋白粉:上海源葉生物科技有限公司;AAPH:上海麥克林生化科技有限公司;三羥甲基氨基甲烷[tris(hydroxymethyl)aminomethane,Tris]、乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)、5,5′-二硫代雙(2-硝基苯甲酸)[5,5′-dithio bis-(2-nitrobenzoic acid),DTNB]、甘氨酸:廣州賽國生物科技有限公司;硫酸銨、無水磷酸二氫鈉、十二水合磷酸氫二鈉:國藥集團化學試劑有限公司。所用試劑均為分析純。
1.2 儀器與設備
FreeZone12L 真空冷凍干燥機:美國LABCONCO公司;F-7000 熒光分光光度計:日本日立公司;M200 PRO 酶標儀:瑞士Tecan 公司;Zetasizer Nano-ZS 型納米粒度和Zeta 電位分析儀:英國Malvern 公司;JSM-6390LV 掃描電子顯微鏡:日本電子JEOL 公司;Vertex 70 傅里葉紅外光譜儀:德國Bruker 公司。
1.3 試驗方法
1.3.1 花生球蛋白的提取
參考陳鵬梟等[9]的方法并加以修改,稱取適量花生蛋白粉溶于0.5 mol/L NaCl、pH7.9 的磷酸鹽緩沖液中,在室溫下攪拌2 h 后,于4 ℃環境下,以4 000 r/min離心30 min,取沉淀用上述磷酸鹽緩沖液按1∶5(g/mL)復溶,然后透析48 h,凍干備用。
1.3.2 過氧自由基氧化花生球蛋白的制備
參考楊曦等[10]的方法并略加修改,稱取1 g 花生球蛋白溶于100 mL 0.01 mol/L 磷酸鹽緩沖液中(pH7.4),分別加入濃度為0.5、1.0、5.0、10.0、15.0 mmol/L 的AAPH,37 ℃避光反應24 h 后,冰浴降溫至4 ℃,于4 ℃透析72 h,凍干后為氧化花生球蛋白,以不加AAPH 的花生球蛋白作為空白對照。
1.3.3 花生球蛋白濁度的測定
稱取10 mg 樣品溶于10 mL 0.01 mol/L 磷酸鹽緩沖液中(pH7.0),在40 ℃的水浴鍋中水浴30 min 后取出,冷卻后在600 nm 處測定吸光度,以吸光度代表濁度。
1.3.4 游離巰基含量的測定
參考范婷等[11]的方法并略加修改,稱取9 mg 的樣品溶于3 mL 的Tris-甘氨酸緩沖液(pH8.0)中,取2.5 mL上述溶液并在其中加入20 μL DTNB 試劑,混合均勻避光反應10 min,在412 nm 處測上清液吸光度。游離巰基含量(X,μmol/g)的計算公式如下。
X=73.53×A412nm×D/C
式中:A412nm為吸光度;D為稀釋倍數,1.01;C為蛋白濃度,mg/mL。
1.3.5 粒徑的測定
參考楊晨[12]的方法并略加修改,稱取10 mg 樣品溶于10 mL 0.01 mol/L 磷酸鹽緩沖液中(pH7.0),攪拌均勻后4 000 r/min 離心20 min,取上清液,然后將上清液通過0.45 μm 濾膜除去不溶性的雜質后進行粒徑的測定。
1.3.6 Zeta 電位的測定
參考李學鵬等[13]的方法并略加修改,稱取10 mg樣品溶于10 mL 0.01 mol/L 磷酸鹽緩沖液中(pH7.0),攪拌均勻后4 000 r/min 離心20 min 取上清液,然后將上清液通過0.45 μm 濾膜除去不溶性的雜質后進行Zeta 電位的測定,測定溫度為25 ℃,溫度平衡時間為2 min。
1.3.7 內源熒光的測定
參考朱增芳[14]的方法并略加修改,稱取10 mg 樣品溶于10 mL 0.01 mol/L 磷酸鹽緩沖液中(pH7.0),使用熒光分光光度計測定蛋白質的固有發射熒光光譜,在290 nm 處激發,掃描波段為300~400 nm,狹縫寬為5 μm,靈敏度為1,記錄光譜的最大發射波長以進行分析。
1.3.8 傅里葉紅外光譜的測定
參考沈明娟等[15]的方法略作修改,稱取1 mg 凍干氧化花生球蛋白和100 mg 干燥的KBr 粉末在研缽中混勻并充分研磨,稱取適量進行壓片,在4 000~400 cm-1內測定紅外吸收光譜,吸光分辨率2 cm-1,掃描次數32 次,利用OMNIC 軟件對圖譜進行分析,并利用Peakfit Version 4.12 軟件對圖譜中酰胺Ⅰ區(1 700~1 600 cm-1)進行解析,根據二級結構分區計算各區域內子峰面積占總峰面積之比,確定蛋白質4 種二級結構的相對含量。
1.3.9 掃描電子顯微鏡測定
參考王莉[16]的方法略作修改,采用掃描電子顯微鏡,取凍干后少量花生球蛋白樣品,將樣品均勻涂抹在帶導電膠的干凈樣品臺上,隨后樣品經噴金處理30 min,在20 kV 電壓下用掃描電子顯微鏡觀察其微觀形貌,放大倍數為800 倍。
1.4 數據處理與分析
所有試驗均重復3 次以上,試驗結果采用Origin Pro 2021 軟件分析作圖,SPSS 25.0 軟件進行顯著性分析。P<0.05 表示差異顯著。
2 結果與分析
2.1 過氧自由基氧化對花生球蛋白濁度的影響
濁度是指溶液對光線通過時所產生的阻礙程度,包括懸浮物對光的散射和溶質分子對光的吸收,可以反映氧化過程中蛋白質的聚集狀態。樣品在600 nm下的吸光度表示其濁度,其結果如圖1 所示。
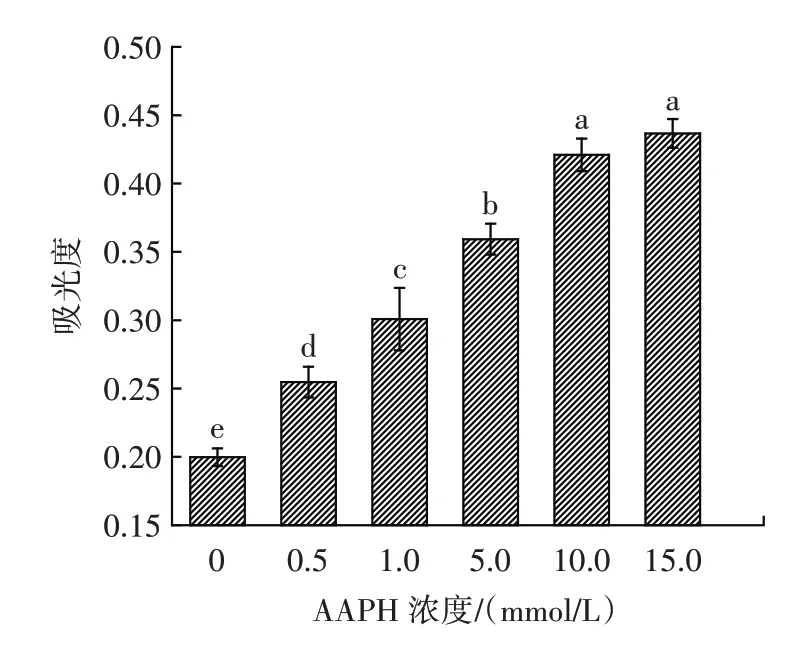
圖1 不同濃度的AAPH 對花生球蛋白濁度的影響Fig.1 Effect of different concentrations of AAPH on the turbidity of arachin
由圖1 可知,花生球蛋白隨濃度的增加其濁度也顯著增加(P<0.05),表示其濁度的吸光度由0.20±0.01增加至0.44±0.01,15.0 mmol/L 氧化時導致濁度上升程度最高,上升了1.2 倍。氧化后的花生球蛋白濁度明顯上升,說明羥自由基氧化使蛋白質之間發生交聯反應,導致蛋白溶液光學密度增加[15]。這表明氧化使蛋白質分子結構發生了變化,形成了小分子聚集體,且隨氧化程度的上升,小分子聚集體逐漸增多,濁度上升,這也與Jiang 等[17]的結果一致。魏娜[18]的研究結果也表明,在同一溫度下,鴨肉肌原纖維蛋白的濁度大小與羥自由基濃度成正比,氧化作用會影響鴨肉肌原纖維蛋白不可溶聚集體的生成,導致其濁度的增加。
2.2 過氧自由基氧化對花生球蛋白游離巰基的影響
巰基以自由基形式存在或形成二硫鍵,在蛋白質的二級、三級結構中發揮重要作用。游離巰基含量可以用來表征花生球蛋白的氧化程度,不同濃度的AAPH對花生球蛋白的游離巰基的影響如圖2 所示。
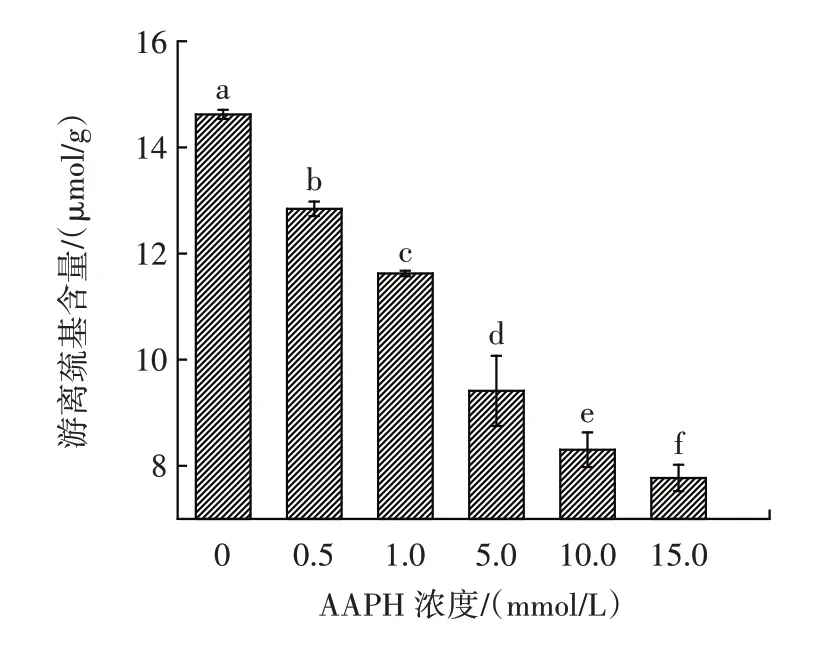
圖2 不同濃度的AAPH 對花生球蛋白游離巰基的影響Fig.2 Effect of different concentrations of AAPH on free sulfhydryl group of arachin
由圖2 可知,隨著氧化程度的增加,花生球蛋白的游離巰基含量顯著下降(P<0.05),從(14.62±0.08)μmol/g 下降到(7.77±0.25)μmol/g。當AAPH 濃度為15.0 mmol/L 時,游離巰基含量相比對照組降低了48.62%,表明花生球蛋白經AAPH 氧化后,可能會發生可逆的氧化反應,使巰基氧化為二硫鍵和次磺酸狀態,同時也可能發生不可逆的氧化,導致花生球蛋白游離巰基的下降[3]。楊曦等[19]也發現自由基氧化可導致蛋白質的游離巰基含量降低,自由基氧化可影響巰基和二硫鍵交互反應平衡常數的改變。巰基是維持蛋白質結構的一種重要的作用力,巰基含量的降低意味著蛋白質結構發生改變。
2.3 過氧自由基氧化對花生球蛋白粒徑的影響
不同濃度的AAPH 對花生球蛋白粒徑的影響如圖3 所示。
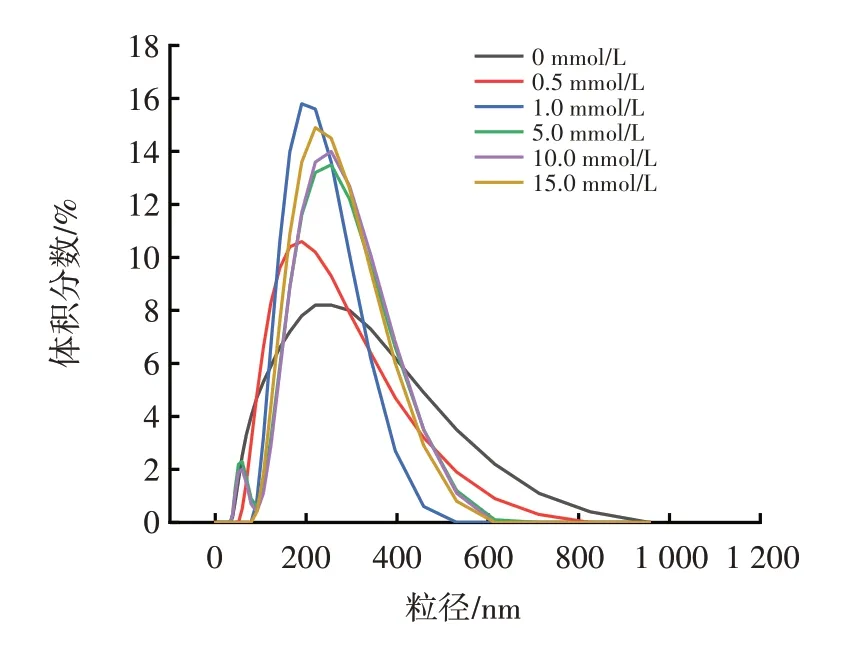
圖3 不同濃度的AAPH 對花生球蛋白粒徑的影響Fig.3 Effect of different concentrations of AAPH on the particle size of arachin
由圖3 可知,不同AAPH 濃度下,花生球蛋白的粒徑總體分布在100~1 000 nm 之間,未氧化的花生球蛋白的粒徑明顯低于氧化后的花生球蛋白的粒徑,且隨著AAPH 濃度的增加氧化花生球蛋白粒徑分布開始向大尺寸方向移動,平均粒徑也從(153.94±3.31)nm增加到(196.80±3.06)nm。這可能是氧化使蛋白分子的疏水基團暴露,在二硫鍵和次級鍵的作用下空間結構發生交聯,生成較大的蛋白質聚集體,表現為蛋白的平均粒徑增大[3]。Ye 等[20]的研究也出現了同樣的結果,AAPH 氧化可導致形成花生蛋白可溶性聚集體。尤翔宇[21]在采用AAPH 氧化米糠蛋白時,氧化米糠蛋白粒徑分布也隨AAPH 濃度的增加呈上升趨勢。本研究中,濁度的試驗結果與粒徑變化的結果一致,進一步說明氧化會使蛋白質分子發生聚集,從而使其粒徑增大。
2.4 過氧自由基氧化對花生球蛋白Zeta 電位的影響
Zeta 電位的絕對值可以表征溶液的穩定性。氧化花生球蛋白的電位變化如圖4 所示。
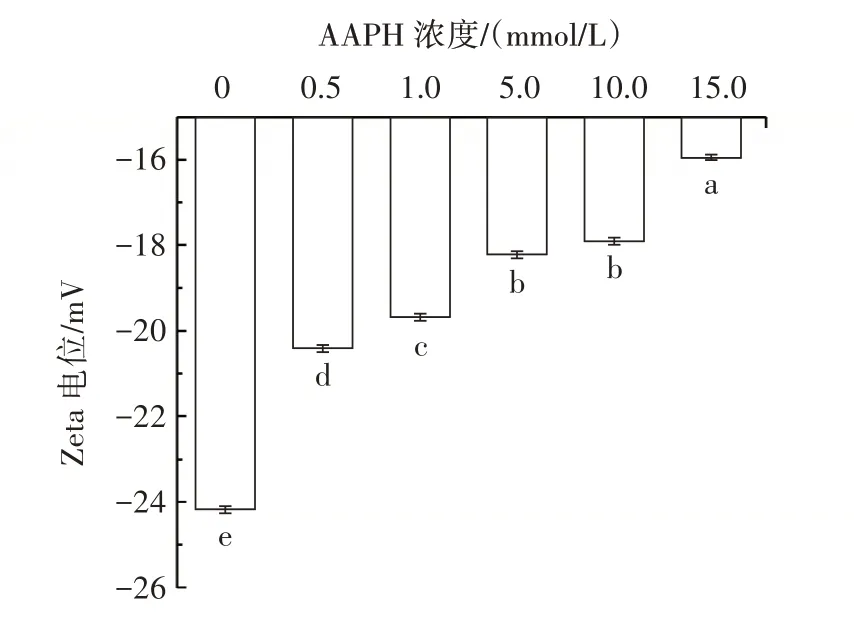
圖4 不同濃度的AAPH 對花生球蛋白Zeta 電位的影響Fig.4 Effect of different concentrations of AAPH on Zeta potential of arachin
由圖4 可知,加入0.5 mmol/L AAPH 后,電位絕對值開始降低,當AAPH 濃度為1.0 mmol/L 時,電位絕對值降低幅度明顯沒有AAPH 濃度為0.5 mmol/L 時降低幅度大,說明氧化改變了花生球蛋白的電位絕對值,從而改變了蛋白溶液的穩定性。然而隨著AAPH濃度的增加,電位絕對值又呈現不同程度的下降,在AAPH 濃度為15.0 mmol/L 時,電位絕對值降低幅度最大,從(24.17±0.06)mV 下降到(15.93±0.05)mV,降低了34.09%(P<0.05),說明高濃度的氧化劑會較大程度地影響溶液的穩定性。蛋白質帶電荷主要是因為氨基酸帶電荷,蛋白質在過氧自由基的攻擊下,一些氨基酸被氧化,導致蛋白質電荷絕對值降低,由于所帶電荷絕對值的減少,蛋白質之間靜電相互作用力減小,這也是蛋白質在氧化后溶解度降低的原因之一[22],此結果與本試驗的氧化后花生球蛋白濁度的變化相互印證。這說明AAPH 氧化使得花生球蛋白氨基酸殘基發生變化,從而破壞花生球蛋白溶液的穩定性,改變花生球蛋白的結構。
2.5 過氧自由基氧化對花生球蛋白熒光光譜的影響
色氨酸最大熒光發射波長可以表示色氨酸殘基在蛋白質中的相對位置,通常用作檢測蛋白質構象變化[23]的標記物。色氨酸殘基在290 nm 激發下,可以在300~400 nm 內發出熒光[5]。不同濃度的AAPH 對花生球蛋白熒光掃描圖譜的影響如圖5 所示。
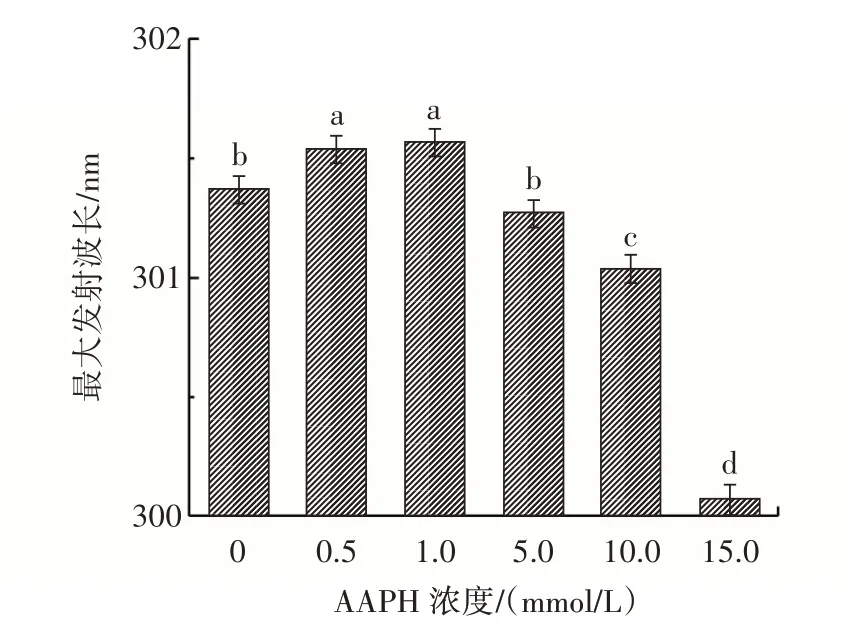
圖5 不同濃度的AAPH 對花生球蛋白熒光掃描圖譜的影響Fig.5 Effect of different concentrations of AAPH on fluorescence scanning spectra of arachin
由圖5 可知,在0~1.0 mmol/L AAPH 之間,最大發射波長從301.4 nm 逐漸增加到301.6 nm,然后在5.0 mmol/L AAPH 時開始下降,在15.0 mmol/L AAPH下,最大發射波長降到300.1 nm。熒光發射向較長波長的位移(紅移)表明發色團(色氨酸殘基)更容易暴露于親水環境中,而較短的波長(藍移)更容易暴露于疏水環境中[23]。這可能是因為在較低的氧化劑濃度時,花生球蛋白暴露出色氨酸殘基,隨著AAPH 濃度的增加,一些暴露的疏水基團參與了疏水作用,形成了不可溶性聚集體,這也與前面的試驗結果(濁度增加、粒徑增大)相一致,說明氧化致使花生球蛋白發生聚集。
2.6 過氧自由基氧化對花生球蛋白紅外掃描光譜的影響
傅里葉變換紅外光譜表征蛋白質二級結構的主要技術方法之一,酰胺I 帶(1 700~1 600 cm-1),是反映蛋白質二級結構變化最常用的譜帶[24]。紅外光譜中蛋白酰胺Ⅰ帶常被劃分為β-折疊(1 636~1 620 cm-1,1 696~1 677 cm-1)、無規則卷曲(1 648~1 640 cm-1)、α-螺旋(1 658~1 649 cm-1)、β-轉角(1 670~1 659 cm-1)[15]。經AAPH 氧化后,花生球蛋白的二級結構變化如圖6所示。
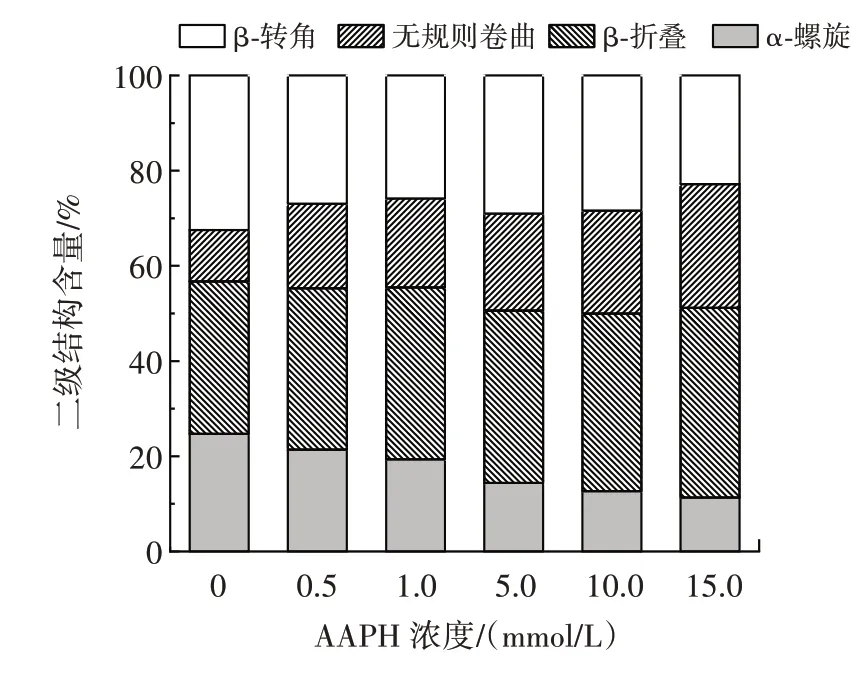
圖6 不同濃度的AAPH 對花生球蛋白二級結構的影響Fig.6 Effect of different concentrations of AAPH on the secondary structure of arachin
由圖6 可知,隨著AAPH 濃度的上升,α-螺旋明顯下降,β-折疊和無規則卷曲明顯上升,說明氧化使花生球蛋白的二級結構發生了較大變化,呈現由α-螺旋向β-折疊和無規卷曲轉變的趨勢。陳曉思等[22]的研究發現,隨著AAPH 濃度的升高,兔肉肌原纖維蛋白的α-螺旋含量逐漸降低,這與本研究結果一致。α-螺旋結構主要依靠蛋白質分子內的氫鍵維持,而自由基攻擊蛋白質的氨基使得蛋白質分子內氫鍵減少,從而導致蛋白質去折疊化[25]。魏娜[18]的研究表明過氧自由基氧化可使鴨肉肌原纖維蛋白的α-螺旋的含量減小,β-折疊和無規則卷曲含量增大,表明過氧自由基能促進氫鍵的斷裂,使鴨肉肌原纖維蛋白朝著不穩定的方向轉變。
為進一步研究氧化后花生球蛋白二級結構的變化,繪制了AAPH 濃度與花生球蛋白二級結構的相關性熱圖,如圖7 所示。
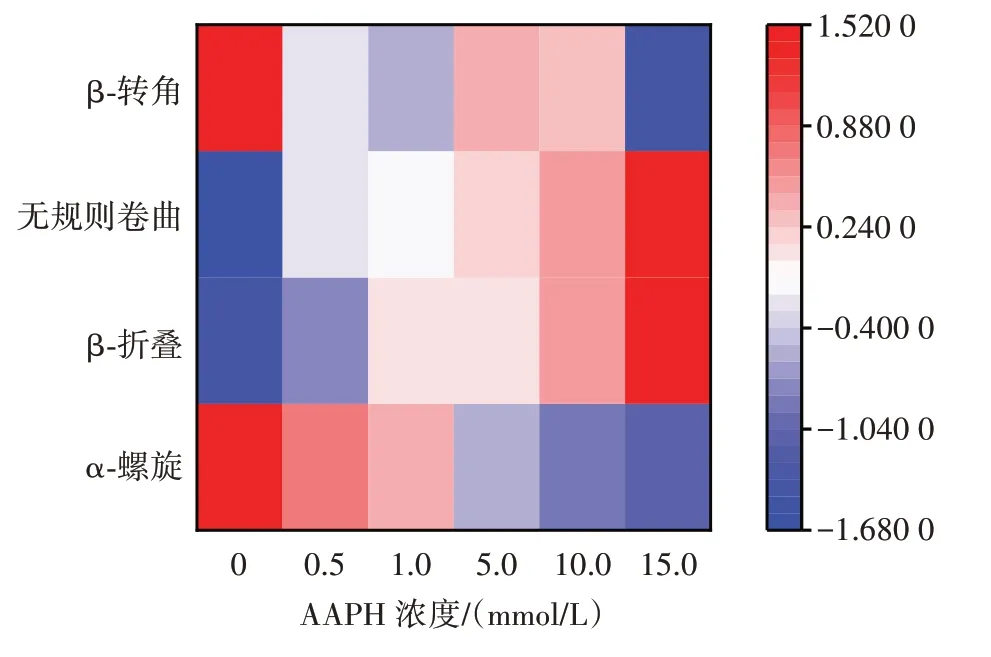
圖7 AAPH 濃度與花生球蛋白二級結構的相關性熱圖Fig.7 Heatmap of correlation between AAPH concentration and the secondary structure of arachin
熱圖中顏色越接近紅色,表示相對值越高,越接近藍色,表示相對值越低。從圖7 可看出,AAPH 濃度與β-折疊和無規則卷曲的變化有較強的正相關,與α-螺旋有較強的負相關。
2.7 過氧自由基氧化對花生球蛋白微觀形態的影響
花生球蛋白過氧自由基氧化的微觀形態如圖8所示。

圖8 不同濃度的AAPH 氧化后花生球蛋白的掃描電子顯微鏡圖Fig.8 Scanning electron microscope of arachin oxidized by different concentrations of AAPH
由圖8 可知,經對比發現,經0.5 mmol/L AAPH氧化后,花生球蛋白明顯聚集起來;當AAPH 濃度為1.0 mmol/L 時,花生球蛋白分子聚集變得更加緊實;5.0 mmol/L 時,可看出蛋白分子內部結構逐漸變得松散;當AAPH 濃度為15.0 mmol/L 時,相比AAPH 濃度為10.0 mmol/L 花生球蛋白表面孔隙明顯增大,且呈現明顯的“蜂窩”狀結構,這與張雪春等[3]的研究結果相似。可能是AAPH 氧化使花生球蛋白分子結構變化,內部松散結構伸展,巰基和疏水基團暴露,并發生相互作用,形成聚集體,這也與花生球蛋白經氧化游離巰基含量逐漸降低,濁度逐漸上升,粒徑增大的結果相對應。周非白[26]研究的豬肉肌原纖維蛋白氧化后的微觀形態也呈現同樣的結果,表明氧化使得蛋白質結構發生了變化。
3 結論
本研究通過AAPH 氧化花生球蛋白,分析其對花生球蛋白結構和聚集行為的影響。結果表明,經AAPH氧化后花生球蛋白溶液濁度顯著增加(P<0.05),這是由氧化后形成蛋白聚集體所造成,這與氧化后花生球蛋白的粒徑顯著增加,形貌孔徑顯著增大的現象相一致。同時AAPH 對花生球蛋白的氧化使其游離巰基含量和電位絕對值下降,降低了花生球蛋白的穩定性,經氧化后花生球蛋白的二級結構顯示由α-螺旋向β-折疊和無規則卷曲轉化的趨勢。氧化后花生球蛋白熒光最大發射波長的藍移與紅移,與花生球蛋白中色氨酸及其殘基在低濃度氧化劑下結構穩定性降低、高濃度氧化劑下參與聚集體有關。由此可見,花生球蛋白經過AAPH 氧化后會發生聚集且其結構穩定性降低,可對花生球蛋白的改性機理研究提供依據。
