404 Not Found
404 Not Found
玉米葉片硝酸鹽積累性狀的全基因組關聯分析
鄔奇 孫揚名 桑俊偉 周玲 趙涵


摘要:為鑒定篩選調控玉米氮素吸收代謝相關位點和基因,以遺傳來源廣泛的350份玉米自交系為材料,利用玉米55K SNP芯片,對玉米葉片硝酸鹽積累性狀進行基于Q+K混合線性模型的全基因組關聯分析。結果表明,350份玉米自交系構成的自然群體具有廣泛的遺傳多樣性,3次重復測量的葉片硝酸鹽含量均表現出良好的正態分布特性。葉片硝酸鹽的全基因組關聯分析顯示,共有27個穩定的SNP位點與玉米葉片硝酸鹽積累顯著相關,這27個位點共分布在玉米6條染色體上,其中16個集中在4號染色體上。對9個極顯著關聯[-lgP≥5.5]位點基因進行氮響應相對表達分析,有8個基因表現出顯著氮響應特性,其中3個基因響應達到極顯著水平,分別較對照提高3.54、2.74、2.02倍,對應已注釋的基因分別為NAC79、GA20ox7和PREP2,表明它們可能在氮素吸收代謝的調控中發揮作用。相關研究結果可作為玉米硝酸鹽吸收代謝研究的重要候選基因進行功能驗證,對開發氮高效分子標記及育種利用具有重要意義。
關鍵詞:玉米;硝酸鹽;全基因組關聯分析;SNP;候選基因
中圖分類號:S513.01? 文獻標志碼:A
文章編號:1002-1302(2024)03-0047-06
氮素作為植物生長的必需營養元素,對作物產量和品質性狀形成起重要作用。目前,全球農業生產的氮肥年使用量已經超過1.2億t,但氮肥利用率很低,氮素損失率達70%[1-2]。氮肥的大量施用,在支撐作物增產的同時,也引起了土壤酸化、水體富營養化、溫室效應加劇等一系列環境問題[3-4]。因此,篩選和培育氮高效型作物種質是協同解決作物產量、資源效率和環境安全的關鍵。
玉米是全球栽培面積最大的糧食作物[5],其產量對氮素的依賴非常顯著。開發和培育氮高效玉米種質資源和新品種,對減少氮肥消耗及降低氮肥引發的環境問題具有重要意義。為解析玉米氮素吸收和代謝相關性狀的遺傳機制,前人做了大量的研究。ZmDof1是最早被發現與玉米氮素吸收調控相關的轉錄因子。在低氮環境下,ZmDof1在水稻中的表達可以提高水稻對碳氮的吸收和同化[6]。Ge等利用泛轉錄組學(pan-transcriptome)手段,對503個不同玉米自交系氮響應轉錄組數據進行分析,構建了氮響應基因表達調控網絡,篩選出處于關鍵節點的ZmNLP5轉錄因子,該調控因子在玉米氮饑餓狀態下的氮吸收和同化過程中起重要作用[7]。MADS-box轉錄因子在水稻和擬南芥中被證明與氮素的吸收和同化密切相關。在玉米中,第一個被發現的MADS-box轉錄因子Zmm28被證明與氮素利用密切相關,過表達Zmm28可顯著提高玉米的產量[8]。Liu等鑒定了1個局部氮響應的MADS-box轉錄因子ZmTMM1,該轉錄因子主要在根中表達,調控不同氮素水平下的根系構型并影響根對硝酸鹽的吸收[9]。Du等通過基因共表達網絡分析玉米和高粱氮響應保守模塊,鑒定了玉米和高粱進化保守的氮調控轉錄因子ZmNIGT1,該轉錄因子是一個負調控因子,在優化氮素營養代謝中發揮重要作用[10]。上述氮素吸收代謝調控因子的發掘一般基于組學和反向遺傳研究,利用正向遺傳學手段挖掘更多玉米中尚未被發現的調控因子亟待進一步探索。
全基因組關聯分析(GWAS)具有效率高、通量大、適用性強、遺傳多樣性覆蓋度高等優點[11]。目前,基于GWAS分析已鑒定到多個水稻氮高效基因位點,如OsTCP19、OsNAC42、OsNPF61、OsNLP4、OsNiR等[12]。上述基因位點的鑒定為氮高效利用分子調控網絡的構建奠定了基礎,其中多個基因已在現代栽培稻氮高效遺傳改良育種中獲得成功應用。到目前為止,通過GWAS在玉米中開展氮高效基因或調控因子的鑒定和挖掘鮮有報道。
本研究使用來自溫帶、熱帶以及亞熱帶且具有豐富遺傳多樣性的350 份玉米自交系組成的關聯群體為試驗材料,對其葉片中的硝酸鹽積累進行表型評價并進行GWAS分析,旨在調查玉米硝酸鹽積累情況及鑒定可能影響硝酸鹽含量的位點及候選基因,研究結果可為玉米氮素吸收代謝、相關調控網絡的構建及育種應用提供理論基礎和科學依據。
1 材料與方法
1.1 試驗材料
試驗材料為350份來源廣泛的優良玉米自交系組成的自然群體。群體材料遺傳背景豐富,形態差異顯著,包含我國溫帶5個雜種優勢群的代表種質及其衍生系、熱帶和亞熱帶種質及來源國外的種質材料。
1.2 試驗設計
350份自交系于2022年種植在江蘇省農業科學院人工智能溫室。為控制統一的氮素濃度,采用水培法培育玉米苗,水培溶液為含2 mol/L KNO3的改良Hoagland營養液。水培容器為50 cm×80 cm×20 cm中轉箱,采用泡沫板懸浮法培養,每盆培養不同玉米自交系幼苗40株。每隔10 d更換1次營養液以保證相對充足的營養環境。試驗共重復3次,分別于2022年4月、5月和9月開展。待玉米長到4葉1心時收獲植物地上部所有葉片,烘干研磨至粉末用于硝酸鹽含量的測定。
1.3 玉米葉片硝酸鹽含量的測定
試驗于玉米生長至4葉1心時取樣,直接剪取植株全部葉片,快速放入烘箱于105? ℃殺青30 min后轉為80 ℃烘干至恒質量,使用粉碎儀將樣品磨成粉末,玉米植株硝酸鹽含量的測定參考王學奎等的方法[13],采用水楊酸-硫酸法測定,具體方法如下:準確稱取0.1 g烘干的葉片樣品于10 mL離心管中,加入5 mL超純水充分混勻,水浴鍋100 ℃水浴30 min,期間每隔10 min充分搖晃均勻,待冷卻至室溫后離心,離心機于室溫15 000 r/min離心10 min,離心結束后吸取100 μL上清于新的10 mL離心管中,加入質量分數為5%的水楊酸-濃硫酸溶液 400 μL,混勻靜置反應20 min,再向離心管中加入9.5 mL質量分數為8%的NaOH溶液,反應冷卻至室溫后,使用紫外-可見光分光光度計在410 nm處測吸光度,根據硝酸鹽標準曲線計算樣品硝酸鹽含量。
1.4 群體結構分析
利用Alexander 等開發的Admixture 1.3軟件進行最佳亞群數的計算[14],對每個K值進行10次獨立重復運算,當交叉檢驗誤差(CV Error)最小時,此時對應的K值即為最佳亞群個數,基于此選用Structure 2.3.4 軟件繪制群體結構圖。
1.5 全基因組關聯分析
全基因組關聯分析選用TASSEL 5.2.28 軟件,聯合群體結構(Q矩陣)及親緣關系(K矩陣)采用混合線性模型(MLM)對基因型數據及硝酸鹽含量進行關聯分析,P值的顯著閾值選擇為10-5,結合R程序進行曼哈頓圖及QQ散點圖的可視化。
1.6 候選基因功能注釋
基于關聯分析結果找到于硝酸鹽含量顯著相關的SNP位點,選用SNP eff工具在玉米V3 (以B73自交系為模板)參考基因組中(https://www.maizegdb.org/)根據顯著SNP位點的物理位置找到其所在的基因或結構域,以LD(r2≤0.1)為范圍篩選硝酸鹽含量性狀的候選基因,將玉米V3參考基因組作為候選基因預測的依據并進行候選基因的功能注釋。
1.7 候選基因氮響應表達分析
參照已發表的RNA-seq數據,對定位到的顯著關聯位點進行表達分析和統計[7]。試驗材料為B73自交系,統計數據為10 mmol/L KNO3處理0、30 min后的表達變化。
2 結果與分析
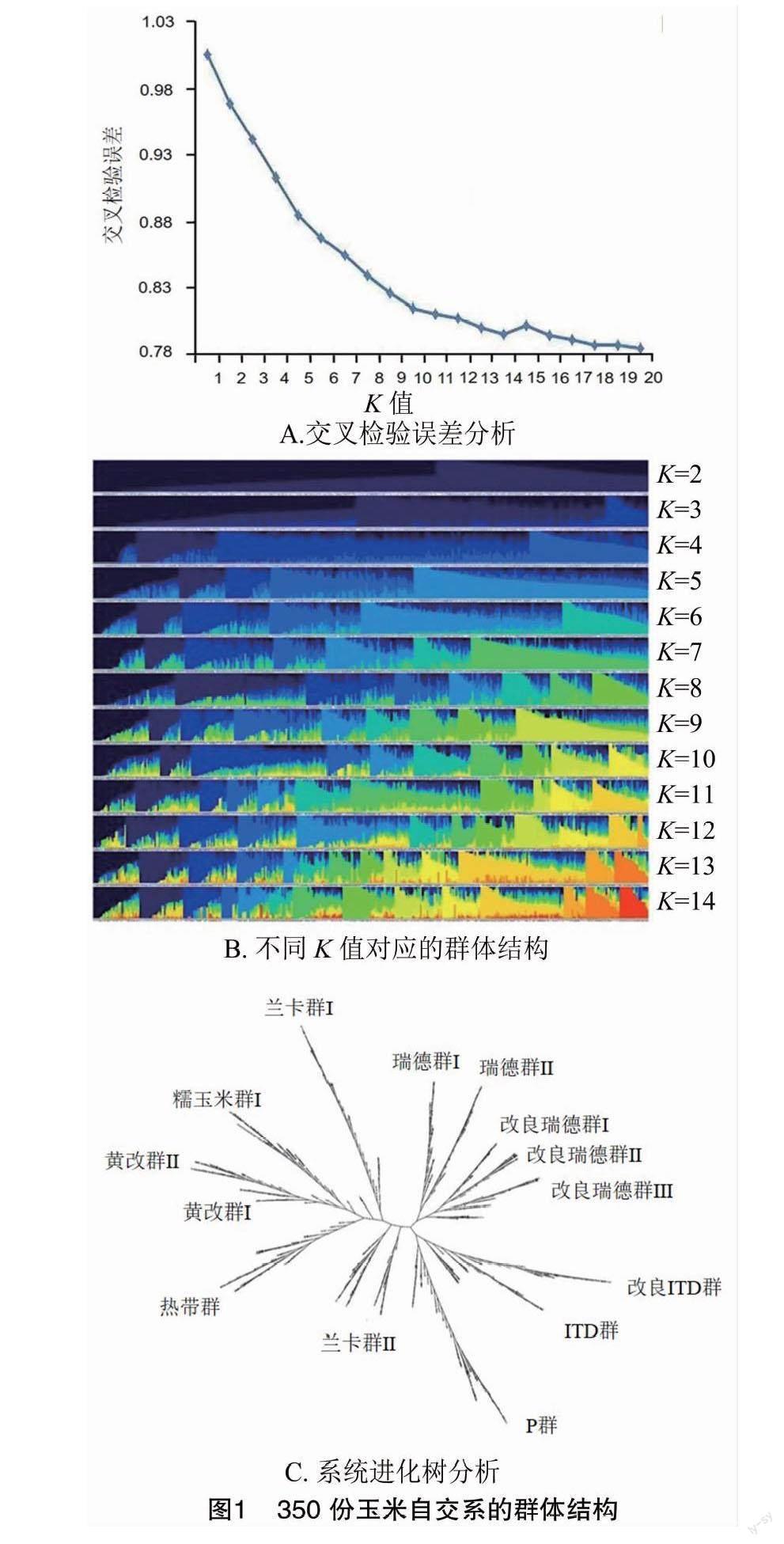
基于Admixture 1.3軟件計算最佳亞群數量,當交叉檢驗誤差(Cross-Validation Error)最小時,其對應的K 值即為最佳亞群數。由圖1-A可見,當K值為14時,交叉檢驗誤差最小。基于此,將350份玉米自交系劃分為14個亞群(圖1-B),分別為黃改群Ⅰ、黃改群Ⅱ、IDT群、改良IDT群、蘭卡群Ⅰ(Mo17)、蘭卡群Ⅱ(OH43)、糯玉米群、P群熱帶群、瑞德群Ⅰ、瑞德群Ⅱ、改良瑞德群Ⅰ、改良瑞德群Ⅱ及改良瑞德群Ⅲ(圖1-C)。
由圖2可見,對350份玉米自交系開展了3次水培試驗,對收集到的玉米葉片開展硝酸鹽含量測定,正態分布分析結果顯示,3組試驗重復中玉米葉片苗期硝酸鹽含量均呈現出中間高兩端低的分布特點,表現出良好的正態分布特征,表明GWAS群里葉片硝酸鹽積累符合數量性狀的遺傳規律,并存在豐富的遺傳變異。
基于45 299個高質量SNP標記對3個不同環境350 份玉米自交系苗期硝酸鹽含量性狀進行全基因組關聯分析。由圖3-A、圖3-C、圖3-E的QQ圖可見,每組重復樣品點分布的實際值與理論值表現出較好的擬合特性,反映了各重復組樣品點具有較好的正態分布特征。
以-lgP≥5為標準統計與硝酸鹽含量性狀顯著關聯位點,共鑒定到27個顯著關聯位點。由圖3-B、圖3-D、圖3-F及表1可見,這27個顯著關聯位點分布在6條染色體上。其中在4號染色體上分布最多,共有16個顯著關聯位點。其他位點分布分別為:1號染色體1個,2號染色體2個,5號染色體3個,6號染色體3個,7號染色體2個。對-lgP≥6的極顯著位點進行統計,結果發現4個與硝酸鹽可能存在極顯著關聯的位點,對應的基因分別為GRMZM2G101271、GRMZM2G478034、GRMZM2G010097和GRMZM2G478034(表1)。
為進一步確定鑒定位點及其對應基因是否對氮素存在響應,對-lgP≥5.5的9個關聯位點及對應基因進行相對表達分析。由圖4可見,與對照(Con:1 mmol/L KNO3)相比,玉米幼苗在高氮(HN:10 mmol/L KNO3)處理15 min后,除GRMZM2G478034以外,其他8個基因均表現出不同程度的氮響應特性,且均達到顯著響應水平,其中基因GRMZM2G033884、GRMZM2G0002704、GRMZM2G101271和GRMZM2G153722的氮響應水平最為顯著,分別為對照的3.54、2.74、2.02、2.51倍。
3 討論與結論
群體結構體現了一個群體中不同亞群的分化程度[15],在進化研究、篩選并展開性狀與分子標記的關聯分析中發揮重要作用。本研究基于交叉檢驗誤差(CV Error)來確定最佳亞群數,將350份玉米自交系劃分成14個亞群,各亞群之間基因交流廣泛,重組變異充足。然而,群體結構會引起分布在不同染色體上的位點間的連鎖不平衡增加,這會導致目的性狀與真實情況并不相關的位點關聯,從而使結果產生假陽性[16]。因此,在進行關聯分析時,必須要考慮到群體結構對GWAS分析結果的影響,然后引入其他協變量或算法加以解決[17]。本研究同時考慮親緣關系與群體結構對最終結果的影響,因而采用混合線性模型(MLM)將2個協變量(Q+K或PCA+K)導入統計計算模型中以此來控制假陽性結果。盡管采用雙矩陣進行質控導致了統計方法過于嚴格、有相關性的位點因其不顯著而被篩除,但保證了本研究鑒定的顯著性關聯位點的可靠性。
采用遺傳力高、科學穩定、遺傳變異豐富、簡單經濟的指標來衡量作物的氮效率是選育氮高效種質的關鍵。在全基因組關聯中,選擇合適的氮素吸收表型指標與自然群體基因型進行關聯分析,對成功挖掘氮素吸收利用相關的功能基因或調控因子至關重要。Liu等通過調查分析低、中、高N條件下產量相關農藝性狀,發現分蘗氮響應(tillering response to nitrogen,TRN)是衡量水稻N利用效率的理想指標,并通過GWAS成功定位一個與氮素利用顯著相關的候選基因OsTCP19[18]。 本研究選擇硝酸鹽含量作為GWAS分析的表型指標,原因如下:一是玉米是旱地作物,主要利用的氮素形態是硝酸鹽;二是硝酸鹽通常被用來作為反映作物氮素積累差異的重要指標,在葉片的積累能夠很好地反映植物本身的硝酸鹽的吸收情況。盡管代謝同化等其他過程也會影響硝酸鹽形態在植物體的積累,但不能完全消除不同植物對硝酸鹽吸收的差異性。本研究的結果顯示,350個玉米自交系葉片硝酸鹽含量存在差異,表現出良好的正態分布特性,表明以硝酸鹽作為GWAS分析的表型指標具有較好的科學性和代表性。
本研究通過3次試驗共定位到了27個顯著位點,在關聯的相關基因組共發現了3個轉錄因子,分別是MYB49(GRMZM2G032655)、GNAT18 (GRMZM2G055141)、NAC79(GRMZM2G033884)。目前為止,關于MYB和GNAT家族轉錄因子在植物氮響應及調控中的研究尚未見報道。在水稻中,NAC家族轉錄因子OsNAC42是重要的氮高效轉錄因子,它與NPF6.1的信號級聯在水稻氮利用及產量提高中發揮重要作用[19]。在玉米中,OsNAC7是調控玉米持綠的關鍵調控因子,在調節葉綠素降解和蛋白質周轉中發揮重要作用,可能與玉米氮素代謝密切相關[20]。本研究中,NAC79的表達受氮素誘導,高氮誘導后其表達提高了3.54倍(圖4),推測可能是玉米氮素吸收或代謝調節的重要轉錄因子,相關表型性狀和機理仍需進一步研究。除此以外,本研究還發現一個重要的功能基因ZmGA20ox7(GRMZM2G0002704)可能與氮素吸收代謝有關。已有報道表明,玉米GA20ox5突變體在低氮條件下葉綠素含量顯著高于野生型,對氮素的敏感性顯著降低[21]。ZmGA20ox7作為同源基因,在氮誘導后其表達上升了2.74倍,表明其可能在調控氮響應及代謝過程中發揮重要功能。
綜上所述,本研究中獲得的顯著關聯 SNP 位點及基因在調控氮素吸收代謝過程中可能具有重要作用。然而,對鑒定的基因進行功能驗證及育種利用需要進一步探索,相關深入研究將為通過分子輔助育種提升玉米的氮素利用效應有重要參考價值。
參考文獻:
[1]Fowler D,Steadman C E,Stevenson D,et al. Effects of global change during the 21st century on the nitrogen cycle[J]. Atmospheric Chemistry and Physics,2015,15(24):13849-13893.
[2]Kanter D R,Bartolini F,Kugelberg S,et al. Nitrogen pollution policy beyond the farm[J]. Nature Food,2020,1(1):27-32.
[3]Zhang X,Davidson E A,Mauzerall D L,et al. Managing nitrogen for sustainable development[J]. Nature,2015,528(7580):51-59.
[4]申建波,白 洋,韋 中,等. 根際生命共同體:協調資源、環境和糧食安全的學術思路與交叉創新[J]. 土壤學報,2021,58(4):805-813.
[5]Hochholdinger F,Yu P,Marcon C.Genetic control of root system development in maize[J]. Trends in Plant Science,2018,23(1):79-88.
[6]Kurai T,Wakayama M,Abiko T,et al. Introduction of the ZmDof1 gene into rice enhances carbon and nitrogen assimilation under low-nitrogen conditions[J]. Plant Biotechnology Journal,2011,9(8):826-837.
[7]Ge M,Wang Y C,Liu Y H,et al. The NIN-like protein 5 (ZmNLP5) transcription factor is involved in modulating the nitrogen response in maize[J]. The Plant Journal,2020,102(2):353-368.
[8]Wu J R,Lawit S J,Weers B,et al. Overexpression of zmm28 increases maize grain yield in the field[J]. Proceedings of the National Academy of Sciences of the United States of America,2019,116(47):23850-23858.
[9]Liu Y,Jia Z T,Li X L,et al. Involvement of a truncated MADS-box transcription factor ZmTMM1 in root nitrate foraging[J]. Journal of Experimental Botany,2020,71(15):4547-4561.
[10]Du H Y,Ning L H,He B,et al. Cross-species root transcriptional network analysis highlights conserved modules in response to nitrate between maize and Sorghum[J]. International Journal of Molecular Sciences,2020,21(4):1445.
[11]楊 彬,周嘉潤,沈玉婷,等. 水稻種子活力性狀全基因組關聯分析研究進展[J]. 江蘇農業科學,2023,51(3):16-21.
[12]Hu B,Wang W,Chen J J,et al. Genetic improvement toward nitrogen-use efficiency in rice:lessons and perspectives[J]. Molecular Plant,2023,16(1):64-74.
[13]王學奎,黃見良. 植物生理生化實驗原理與技術[M]. 3版.北京:高等教育出版社,2015:55-57.
[14]Alexander D H,Novembre J,Lange K. Fast model-based estimation of ancestry in unrelated individuals[J]. Genome Research,2009,19(9):1655-1664.
[15]陳 斐,魏臻武,李偉民,等. 基于SSR標記的苜蓿種質資源遺傳多樣性與群體結構分析[J]. 草地學報,2013,21(4):759-768.
[16]吳永升. 玉米谷氨酰胺合成酶基因Gln1-3、Gln1-4氮利用效率關聯性分析[D]. 北京:中國農業科學院,2009:17-18.
[17]Song M S,Hao W,Storey J D.Testing for genetic associations in arbitrarily structured populations[J]. Nature Genetics,2015,47(5):550-554.
[18]Liu Y Q,Wang H R,Jiang Z M,et al. Genomic basis of geographical adaptation to soil nitrogen in rice[J]. Nature,2021,590(7847):600-605.
[19]Tang W J,Ye J,Yao X M,et al. Genome-wide associated study identifies NAC42-activated nitrate transporter conferring high nitrogen use efficiency in rice[J]. Nature Communications,2019,10:5279.
[20]Zhang J,Fengler K A,van Hemert J L,et al. Identification and characterization of a novel stay-green QTL that increases yield in maize[J]. Plant Biotechnology Journal,2019,17(12):2272-2285.
[21]張肖逢. 玉米ga20ox5矮化突變體和轉ZmGRF1基因玉米的鑒定及氮素利用率研究[D]. 北京:中國農業科學院,2021:62-68.
