農田離子型新興有機污染物的作物吸收、分布和轉化機制研究進展
2024-03-27 01:41:22姜彥岐石傲傲徐凱然任俊明倪卓彪林慶祺仇榮亮華南農業大學資源環境學院廣州廣東5062廣東省農業農村污染治理與環境安全重點實驗室廣州廣東5062嶺南現代農業科學與技術廣東省實驗室廣州廣東5062仲愷農業工程學院資源與環境學院廣東省普通高校農業產地污染綜合防治工程技術研究中心廣州廣東50225中山大學環境科學與工程學院廣州廣東50006
生態與農村環境學報 2024年3期
姜彥岐,黃 健,石傲傲,徐凱然,任俊明,倪卓彪,2,3,阿 丹,林慶祺,2,3①,仇榮亮,2,3,5 (.華南農業大學資源環境學院,廣州 廣東 5062;2.廣東省農業農村污染治理與環境安全重點實驗室,廣州 廣東 5062;3.嶺南現代農業科學與技術廣東省實驗室,廣州 廣東 5062;.仲愷農業工程學院資源與環境學院/ 廣東省普通高校農業產地污染綜合防治工程技術研究中心,廣州 廣東 50225;5.中山大學環境科學與工程學院,廣州 廣東 50006)
隨著社會城市化、工業化發展及農業集約化生產,種類繁多且數量巨大的新興化學品被釋放到環境中。新興化學品大規模的生產使用及長期監管體系不健全,導致我國面臨著嚴峻的新污染物污染問題。《中華人民共和國國民經濟和社會發展第十四個五年規劃和2035年遠景目標綱要》中強調要深入打好污染防治攻堅戰,建立健全環境治理體系,尤其要重視新污染物治理。新污染物是指由人類活動造成的、目前已明確存在但尚未納入管理或者現有法律法規管理措施不足以有效防控其風險、在生產建設或者其他活動中產生、在生態環境中廣泛賦存的所有污染物[1]。目前,國際上關注的新興有機污染物種類主要包括環境內分泌干擾物(environmental endocrine disruptors,EEDs)、全氟及多氟烷基化合物(per- and polyfluoroalkyl substances,PFASs)、藥物與個人護理品(pharmaceuticals and personal care products,PPCPs)及抗生素等[2]。大量報道已證實,新興有機污染物(emerging organic pollutants,EOPs)會對環境和人體健康產生不可忽視的威脅。例如,某些PPCPs會導致溪流中水生生物神經系統紊亂,晝夜節律失調[3],明顯抑制斑馬魚胚胎神經-肌肉生理結構,甚至會導致不可逆發育側彎或胚胎死亡[4]。接觸PFASs會導致一系列人體健康問題,包括肝腎受損、免疫毒性、內分泌干擾和致癌影響[5-8]。某些短鏈PFASs可以對人體干細胞(HL7702)產生很強的細胞毒性,還可能會導致生殖和發育異常[7]。同時,部分EOPs環境持久性較強,擴散分布廣泛,具有長期、潛在和隱蔽的環境危害[1]。值得注意的是,部分離子型新興有機污染物(ionic emerging organic pollutants,IEOPs)含有一個或多個可電離官能團(如—COOH、—SO3H、—NH2和—OH等),在不同環境條件(如pH的改變)下其官能團可以發生質子化或去質子化,因而改變其存在形態[9]。根據IEOPs所帶電荷的電性,可分為陽離子型、陰離子型和兩性離子型。一般來說,IEOPs揮發性較弱,極性強,具有較好的水環境遷移能力,因此其環境行為比傳統非離子型有機污染物更復雜。
前人研究多關注地表水體及再生水中IEOPs污染狀況,近年來土壤環境,尤其是對農田土壤-作物系統的研究逐漸受到重視[10]。再生水灌溉、污泥農用以及農用化學品的使用是IEOPs進入農田的主要途徑。據報道,農田土壤及作物器官中全氟辛烷磺酸鹽(perfluorooctane sulfonate,PFOS)和全氟辛酸(perfluorooctanoic acid,PFOA)等典型PFASs及其新興替代物,抗生素(四環素類、磺胺類、喹諾酮類)及多種藥物成分等PPCPs可被廣泛檢出(表1[11-22])。進入農田環境中的IEOPs經過吸附-解吸及淋濾等過程后,部分被作物吸收,并隨食物鏈傳遞對人體產生潛在威脅[23-25]。研究人員對我國常熟和太倉地區氟化物工業園區周邊農田14種蔬菜和6種水果進行調查,總PFASs檢出濃度分別為0.3~11.5 μg·kg-1和0.1~6.0 μg·kg-1,其中,該區域番茄中PFASs含量遠高于歐洲食品安全局推薦的攝入限值[17]。因此,深入了解作物對IEOPs的吸收累積過程及機制,對明確IEOPs在農田-作物系統中的歸趨具有重要意義。該文將基于該領域相關的最新研究進展,以檢出率較高、環境風險較大的PFASs〔除氟調醇(fluorotelomer alcohols,FTOHs)外〕、PPCPs等IEOPs為代表,系統總結農田中IEOPs的作物吸收轉運、累積分布以及體內轉化過程的機理及影響因素,并對該領域的未來研究方向進行展望。
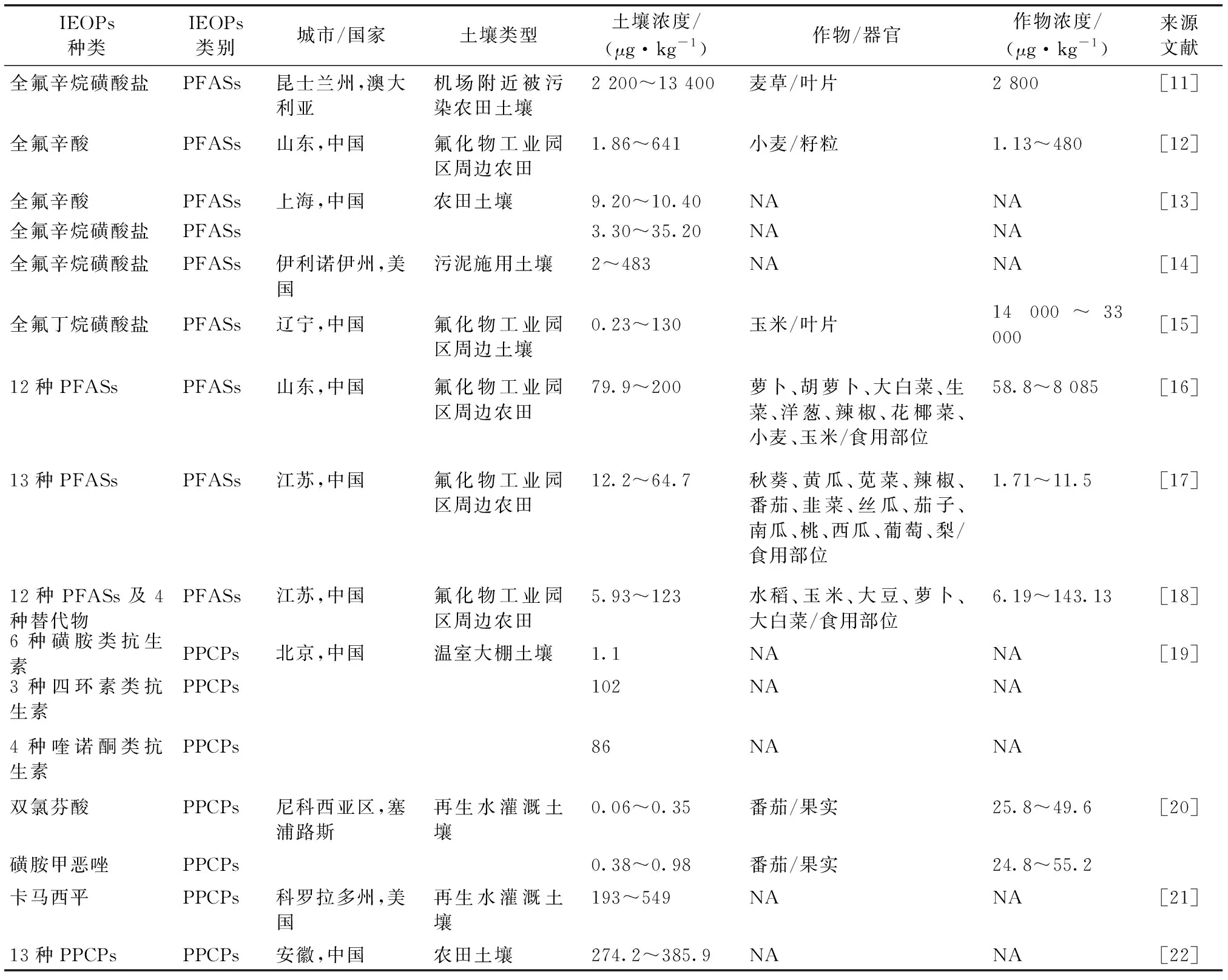
表1 部分國家或地區農田土壤及作物中離子型新興有機污染物(IEOPs)濃度分布[11-22]Table 1 Concentrations of ionic emerging organic pollutants in farmland soils and crops of some countries and regions
1 離子型新興有機污染物的作物吸收、分布與轉化機理
1.1 作物根系對離子型新興有機污染物的吸收過程及機理
農田土壤中IEOPs主要通過根系被作物吸收,并可轉移至地上部和食用部位。PAN等[26]設置兩種濃度抗生素加污土培試驗,驗證了生菜、蘿卜和番茄的抗生素吸收現象,測得作物組織中濃度最高的是恩諾沙星(2.4~10.3 μg·kg-1)和氯霉素(0.76~1.85 μg·kg-1)。研究者通過市政污泥添加試驗,證實胡蘿卜、蘿卜和馬鈴薯塊莖均可以吸收PFASs,推測胡蘿卜的食用部位是吸收養分、水分的肉質根組織,具有較高的PFASs吸收潛力[27-28]。作物根系吸收IEOPs后,可以將其進一步轉移至莖葉等營養器官,并累積于繁殖器官中。
一般認為,污染物從根表徑向遷移至維管組織主要有質外體和共質體兩種途徑(圖1)。質外體途徑指污染物通過細胞壁、細胞間隙等非胞質空間移動;而共質體途徑則指污染物借助胞間連絲完成胞質間的傳遞。研究表明,疏水性IEOPs主要通過質外體途徑向維管組織移動,而親水性IEOPs則利用共質體途徑發生遷移[29]。然而,由于木質素及栓質沉積在初生細胞壁和胞間層形成凱氏帶,通過質外體空間遷移的水和溶質需要轉為共質體方式[30],即必須至少跨越一層磷脂雙分子層,才可以遷移至維管組織繼而向上轉運[31]。
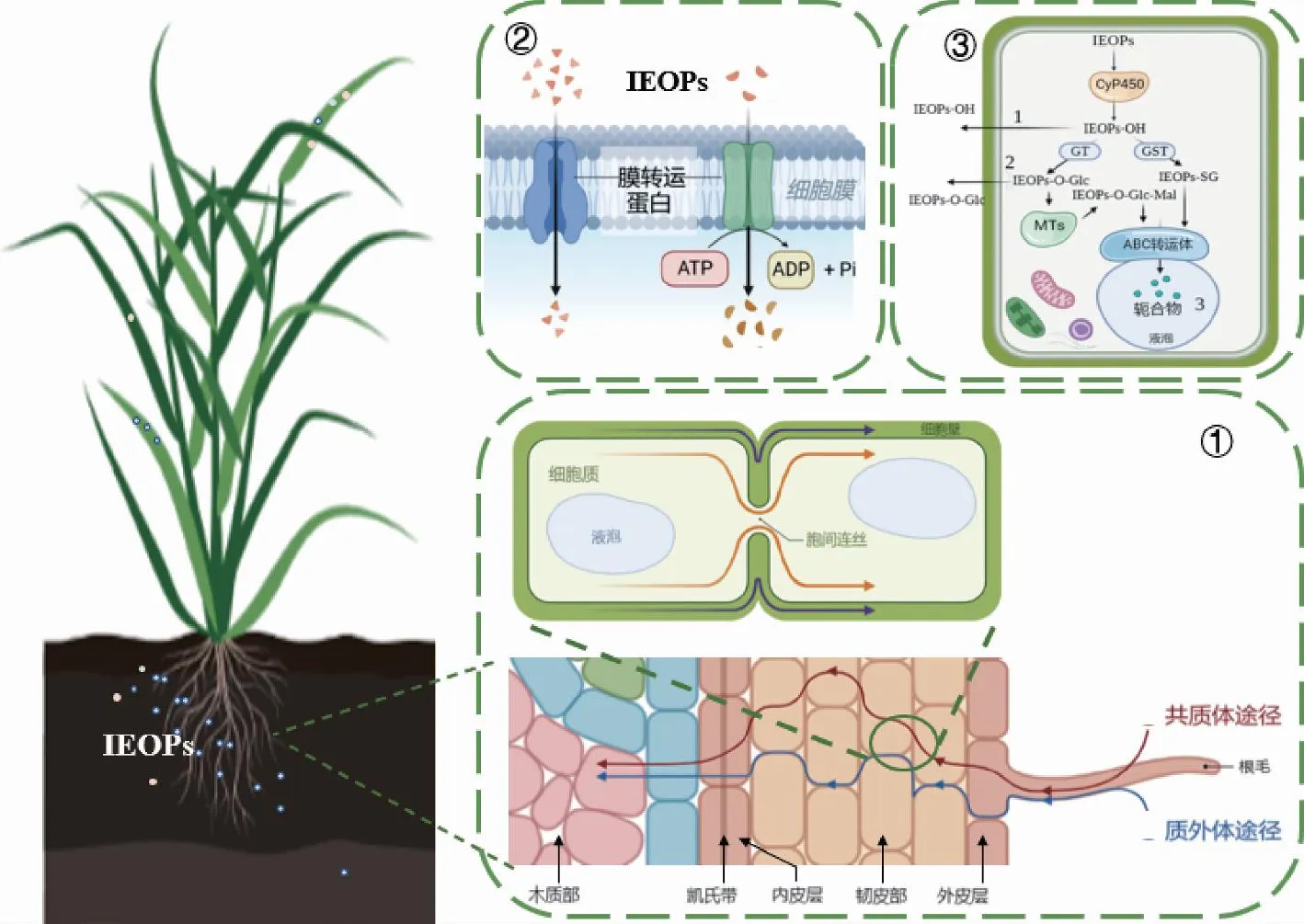
①吸收過程;②跨膜轉運過程;③轉化過程。IEOPs·OH指IEOPs發生親水基團修飾(如加羥基);IEOPs-SG、IEOPs-O-Glc和IEOPs-O-Glc-Mal指IEOPs與細胞內源物質結合(如氨基酸、多肽等)。CyP450為細胞色素P450酶;MTs為丙二酰基轉移酶;GST為谷胱甘肽巰基轉移酶。圖1 作物吸收轉運和轉化離子型新興有機污染物(IEOPs)的過程機制Fig.1 Absorption, transport, and transformation of ionic emerging organic pollutants (IEOPs) by plants
作物吸收轉運IEOPs與化合物的理化性質(疏水性、相對分子質量、分子結構等)和作物生理(組成成分、轉運蛋白、調控基因等)密切相關。傳統觀點認為有機污染物的疏水性(即辛醇-水分配系數,Kow)是影響作物吸收最重要的因素之一。由于辛醇-水分配體系與作物體內脂細胞膜-胞質溶膠體系對有機物的分配模型相似,故而研究人員普遍利用Kow來代替脂質-水分配系數(Klip)以表征有機污染物在脂相和水相之間的分配行為[32]。有研究[33]指出,作物根部有機污染物的富集系數(root concentration factor,RCF,即污染物在根中與土壤或溶液中濃度之比)與污染物的Kow呈線性關系。然而,由于部分IEOPs存在離子形態,導致它們的吸收行為與非離子型有所不同。IEOPs的電離能力也影響作物的吸收積累狀況,例如,利用懸浮培養細胞試驗驗證了弱酸性除草劑苯達松的體內積累[34]。帶電分子可能降低作物吸收潛力,這是因為電離可能降低它們滲透細胞膜的能力。因此,研究人員提出使用pH依賴型辛醇-水分配系數(logDow,即根據電離基團對Kow依賴性分布的影響經校正后的參數)來評估離子化的IEOPs在作物中的吸收情況[31]。該系數與化合物的酸堿解離常數(pKa)及介質環境pH值有密切關系。WU等[35]比較了8種蔬菜中19種PPCPs累積狀況,堿性和中性PPCPs吸收量高于酸性PPCPs。然而,僅僅通過疏水性大小(如Kow、Dow)不足以評估IEOPs遷移轉運能力[25,36-37]。LIPINSKI等[38]基于IEOPs分子特性,如相對分子質量(relative molecular mass,RMM)、logKow、氫鍵供體數(H-bond donors,即分子中帶有的—OH和—NH總數)及氫鍵受體數(H-bond acceptors,即分子中帶有的N和O原子總數),提出作物吸收有機污染物的“規則5”(Rule of 5),指當一種有機物RMM >500,logKow>5,氫鍵受體數>10,或氫鍵供體數>5時,作物吸收該有機物的能力較弱。KUMAR等[39]基于“規則5”進一步細分提出“規則3”(Rule of 3)和“規則3~5”(Rule of 3~5)。“規則3”是指當有機物RMM <300,logKow<3,氫鍵受體數<6或氫鍵供體數<3時,作物吸收該物質的能力較強。而“規則3~5”則指當有機物RMM為300~500,logKow為3~5,氫鍵受體數為6~10,氫鍵供體數為3~5時,作物可以中等程度地吸收該類物質。該原則為優化作物吸收IEOPs的機理模型提供了理論基礎,然而作物吸收IEOPs的實際狀況并非僅取決于化合物性質。
根系脂質和蛋白質含量等作物組成成分,在用于評估作物吸收IEOPs時也具有重要作用[40],且該生理因素具有明顯的種間差異。WEN等[41]通過比較7種土培作物(紫花苜蓿、生菜、玉米、綠豆、蘿卜、黑麥草、大豆)脂質和蛋白質含量對PFOA和PFOS吸收量的影響,發現根系對兩種污染物的RCF與根系蛋白質含量呈顯著正相關,與脂質含量呈顯著負相關,這可能是由于脂質與污染物競爭蛋白質的吸附位點所致。但最近的研究卻發現8種植物根系脂質含量與PFOA和PFOS累積量呈正相關[42]。考慮到IEOPs種類繁多,可能具有親水、親脂或親蛋白等多種特性,因此,它們與作物的不同組成成分之間的關系仍有待深入挖掘。
早期對于IEOPs作物吸收過程的研究,主要針對化合物的物理化學性質差異展開[43],然而,對于吸收過程中的跨膜機制探討仍較少。LIMMER等[44]發現應用于哺乳動物系統的藥物跨膜遷移體系可以應用于作物吸收轉運IEOPs的過程。近年來,不少研究已證實某些PFASs、氨基糖苷類抗生素、二甲雙胍等IEOPs的跨膜吸收過程涉及蛋白質載體介導[28,45-52]。目前,學界多利用能量代謝抑制劑和水/離子通道阻斷劑等來研究相關機制。KONG等[53]添加能量代謝抑制劑和水通道阻斷劑后,發現苜蓿吸收土霉素是需要耗能的轉運蛋白介導過程,且可能與細胞膜上的水通道蛋白無關。MENG等[54]使用能量代謝抑制劑證實菜心對環丙沙星的吸收是主動過程。WEN等[50]通過水通道和陰離子通道阻斷劑以及能量代謝抑制劑試驗,發現PFOS的吸收主要是水通道和陰離子通道等蛋白載體參與的被動過程,而PFOA則是陰離子通道參與并依賴能量代謝的主動過程。YU等[55]指出,蛋白介導的蒸騰作用以及質膜上的通道蛋白在PFOS低累積作物吸收轉運PFOS過程中起到重要作用。該研究者進一步使用PyMOL分子圖形系統,構建了擬南芥陰離子通道蛋白SLAC1和菠菜水通道蛋白SoPIP2;1空間結構,并根據PFOS和PFOA分子大小證實了這些污染物具有通過陰離子/水通道的理論可能[56-59]。目前關注的IEOPs多為陰離子型,而對陽離子型IEOPs的吸收轉運機理研究相對較少。CUI等[52]通過抑制有機陽離子轉運體(organic cation transporters,OCTs)活性后,發現寬葉香蒲根系對陽離子藥物成分二甲雙胍的吸收量降低超過70%。
最新的一些研究陸續發現調控IEOPs跨膜吸收的相關基因。YU等[60]利用PFOS高、低累積型生菜品種研究PFOS吸收累積差異的內在機制,在發現PFOS吸收是一個載體介導的被動過程的基礎上,進一步分析了負責調控水通道的PIPs(PIP1-1和PIP2-2)、快/慢速陰離子通道的ALMTs(ALMT10和ALMT13)和SLAHs等基因的表達情況,首次證實水通道與快速陰離子通道基因參與了調控PFOS的吸收過程。FAN等[61]通過擬南芥PFOA暴露試驗比較分析根部及地上部的吸收轉運特征,發現擬南芥地上部及根部ABC轉運蛋白(ATP-binding cassette transporters)、藥物跨膜轉運蛋白和替代活性跨膜轉運蛋白等12個轉運蛋白相關基因均發生顯著上調,表明擬南芥可能通過調控這些轉運蛋白參與PFOA吸收。然而,針對其他類型IEOPs的作物吸收調控基因及轉運蛋白研究仍有待進一步深入。
1.2 離子型新興有機污染物的作物累積分布機理
作物根系吸收IEOPs后,可以將其轉移至地上部及可食用部分,沿食物鏈對人體健康產生威脅。BLAINE等[28]通過溫室土培試驗驗證了多種作物的PFASs遷移累積現象,研究發現芹菜莖部中總PFASs濃度可達39.3~817.3 μg·kg-1,生菜葉片中檢出全氟丁酸、全氟戊酸濃度高達266和236 μg·kg-1,番茄果實中全氟丁酸、全氟戊酸濃度分別達56和211 μg·kg-1[46]。LIU等[12]對大型氟化工園區周邊農田作物PFASs含量進行檢測的結果表明,玉米籽粒中檢出濃度為58.8 μg·kg-1,小麥籽粒中檢出濃度高達480 μg·kg-1。該研究中,除了傳統長碳鏈全氟化合物PFOA被檢出外,多種新興全氟化合物替代物(如短鏈全氟羧酸)正逐漸成為作物籽粒中PFASs主要富集類型。
IEOPs的理化性質(相對分子質量和logKow)影響著其在作物根、莖和葉中的遷移與累積分布。研究者利用水稻對3種農藥的吸收轉運及亞細胞分布規律進行研究,結果表明新煙堿類(logKow為0.66~0.8)主要集中在葉片中,三唑類(logKow為3.72~4.4)主要集中在根部[62]。IEOPs的相對分子質量和logKow對其在作物根、莖和葉中的富集程度及莖葉轉移過程具有重要影響。同時,IEOPs的分子結構(如鏈長)也被證實是影響其在植物組織中分布的關鍵因素。PFASs的碳鏈越長,其logKow越大,水培作物根系富集PFASs的能力越強,但往地上部的轉運則越弱[63]。KRIPPNER等[64]為探明C4~C10的10種PFASs在玉米中累積分布規律,通過試驗發現玉米地上部傾向累積C4~C7的短碳鏈PFASs,而碳鏈較長的PFASs則較多截留在根部。總體來說,與小麥相比,玉米秸稈部位傾向于富集短碳鏈PFASs,而對于C8的PFASs,小麥秸稈累積能力則更強。XU等[65]的研究同樣驗證了此結論,發現長鏈PFASs(≥C8)在生菜根部積累量更大,而短鏈PFASs(≤C7)則優先向地上部遷移。
污染物亞細胞分布是不同作物品種累積和轉運有機污染物的重要因素,作物可以通過自身的組織結構影響污染物在其體內轉運過程,利用吸附、阻隔等方式,削減外源污染物對作物生理生化進程的影響。細胞壁是阻止IEOPs進入作物體內的重要屏障,參與了作物的初步解毒過程。果膠、纖維素、半纖維素和木質素等成分組成網狀結構通過直接吸附限制IEOPs跨膜運輸,減少或阻止IEOPs對原生質體的毒害[66]。然而,并非所有外源化合物都能被細胞壁阻隔。WANG等[67]通過研究4種新煙堿農藥在白菜體內亞細胞分布規律發現,低親脂性農藥噻蟲嗪更容易累積在作物細胞可溶性組分中,中等親脂性農藥螺旋霉素則主要分布在細胞壁和可溶性成分中,而高度親脂性農藥阿維菌素則主要分布于細胞壁和細胞器中。IEOPs的亞細胞分布規律在屬間、種間存在差異。在生菜吸收累積PFOA的研究中,低累積生菜品種主要將PFOA富集在細胞壁及葉綠體中,而高累積品種則更多富集在細胞膜及液泡等脂膜中[60]。
分子成像是指在組織、細胞及亞細胞水平實現對污染物分子空間分布的原位成像手段,可以更為詳備地了解有機污染物及其代謝產物的空間分布特征及時空動態變化。其中,質譜成像技術(mass spectrometry imaging,MSI)通過產生任意指定質荷比化合物的二維離子密度圖,實現對生物組織切片中化合物組成、相對豐度及空間分布情況的原位表征[68]。最近,WANG等[69]利用解吸電噴霧電離質譜(desorption electrospray ionization mass spectrometry,DESI-MS)和透射電子顯微鏡結合X射線能譜(transmission electron microscopy equipped with energy-dispersive spectroscopy,TEM-EDS)觀察了兩種典型PFASs在不同濕地植物根組織和細胞中的分布,結果顯示PFOS和PFOA在美人蕉、再力花、風車草和蘆葦根的表皮、皮層和維管束中有分布。其中,在美人蕉和風車草皮層中的PFOS和PFOA主要通過質外體途徑,更多更快地將PFOS和PFOA運輸至維管束,并分布于植物皮層細胞的線粒體及高爾基體等多種細胞器中。質譜成像技術可為后續深入了解作物體內IEOPs的空間定位及脅迫代謝效應提供新的技術手段。
1.3 離子型新興有機污染物的作物體內轉化機制
學界主流觀點認為,有機污染物被作物吸收進入細胞后,可以在不同系列酶的共同作用下進行代謝轉化。轉化過程涉及3個階段。第一階段為親水基團修飾,即通過氧化還原、水解及脫烷基反應獲得親水性官能團(如—COOH、—NH2、—OH等),此階段主要由細胞色素P450酶(cytochrome P450,CYP450s)、過氧化物酶等參與。第二階段為內源分子結合,即修飾后產物在酶的作用下,與作物細胞內源分子(如多肽、氨基酸、多糖等)結合,形成具有二硫鍵、醚鍵等共軛化合物[70]。其中,糖基化共軛產物可以在丙二酰基轉移酶(malonyltransferase,MTS)的介導下與丙二酸反應,生成丙二酰化共軛產物[71]。第三階段為轉化產物區隔化,即共軛產物被轉運至非活性部位,水溶性較強的共軛產物借助ABC轉運蛋白(ATP-binding cassette transporter)運輸至液泡內部,而較難溶的則外排至細胞壁中[32]。通過三相反應,有機污染物的極性及反應活性有所增強,更容易被作物細胞所代謝。一般有機污染物毒性會隨著轉化而逐漸減小,但也存在某些有機污染物的轉化中間產物比母體化合物具有更高毒性的現象[72]。
有機污染物在作物細胞內部的轉化步驟復雜,需要一系列酶的參與。細胞色素P450-含單加氧酶(cytochrome P450-containing monooxygenases)、過氧化物酶(peroxidase)和酚氧化酶(phenoloxidases)是有機污染物代謝研究中受到關注較多的3種酶。作物體內轉化過程可以影響環境有機污染物的持久性或生物積累,特別是那些易受細胞色素P450酶攻擊的污染物[73]。細胞色素P450-含單加氧酶是位于內質網質膜上的混合功能氧化酶系,它們在有機污染物的羥基化過程中起著重要作用。藥物成分布洛芬在作物組織中經過細胞色素P450-含單加氧酶氧化,接著被糖基轉移酶偶聯,最終分離至液泡或細胞壁中。雖然布洛芬的4種中間轉化產物可以被檢出,但在該實驗中未觀察到明顯的作物毒性[74]。研究者在番茄葉片中分別鑒定出卡馬西平的21種相I和10種相Ⅱ轉化產物[75],并提出卡馬西平在作物中的完整代謝轉化途徑,該途徑經過環氧化、水解、羥基化和去氨甲酰基等一系列代謝過程后,與葡萄糖或半胱氨酸軛合。研究表明,作物可以利用谷胱甘肽(glutathione,GSH)與多種PPCPs結合進行解毒。如藥物成分對乙酰氨基酚被作物吸收后,可迅速與GSH結合,在黃瓜根和葉中形成GSH-對乙酰氨基酚軛合產物[76]。然而值得注意的是,一些代謝產物可能表現出與其母體化合物相似或等效的毒性。例如,卡馬西平的代謝產物10,11-環氧卡馬西平具有比母體物質更大的潛在遺傳毒性[77]。類似地,研究者在黑麥草中觀察到FTOHs可以代謝為持久性更高、化學穩定性更強的PFOA[78]。
近年來有研究指出,過去被認為難以在作物體內轉化的IEOPs,如全氟烷基次膦酸,可以轉化為結構更穩定的全氟烷基膦酸并儲存在作物液泡中[79]。因此,對于IEOPs的具體代謝過程和涉及的酶促反應仍然有待開展深入研究。
2 結論與展望
農田IEOPs污染狀況日趨嚴重,不容忽視,目前關于作物對IEOPs的吸收累積行為研究尚處于起步階段。作物可通過根系從土壤中吸收IEOPs,并在蒸騰作用下從根部遷移至莖葉甚至果實。IEOPs的吸收遷移過程和累積分布特征與其自身的理化性質(疏水性、相對分子質量、分子結構等)、作物品種特性(組成成分、吸收載體類型、調控基因等)有著重要關系。在作物體內的IEOPs也會在多種酶(細胞色素P450酶、糖基轉移酶等)的介導下,發生3個階段的系列轉化反應,實現污染物的解毒。然而,今后仍存在以下問題有待深入探索:
(1)需要加強作物吸收累積IEOPs機理的研究。以往對于作物吸收IEOPs的研究大多采用線性回歸方程關系將logKow、logDow、RMM等污染物理化性質參數與作物吸收能力聯系起來。然而,這些關系可能僅適用于特定的化合物和作物物種之間。“規則3”和“規則5”等概念模型的提出為科學評估作物吸收IEOPs能力提供了新的標準。然而,探討作物吸收能力也需要考慮作物品種、轉運體性質及相關基因等諸多作物生理因素。深入探究作物吸收IEOPs的潛在機理,尤其是蛋白轉運體及相關基因功能挖掘工作具有重要意義。另外,目前原位觀察IEOPs在作物體內動態吸收遷移過程的工作僅能針對作物單一器官展開,有待建立完整了解IEOPs在根-莖-葉-果中動態遷移過程的技術體系。
(2)需要繼續完善預測作物吸收累積IEOPs的模型構建。當前學界多只通過logKow、相對分子質量以及作物脂質成分等參數構建預測模型,然而部分IEOPs的可離子化特性導致其實際吸收累積程度與模型預測結果相差較大。此外,在實際作物吸收過程中,復雜的土壤-根系界面過程以及IEOPs在作物體內的轉化均會對模型預估結果產生嚴重干預。因此,在預測模型構建過程中,需納入更多的參數指標,同時結合更多基礎數據,以提高模型的有效性及參考價值。
(3)需要關注作物體內IEOPs轉化機制的研究。雖然IEOPs等有機污染物被作物吸收進入細胞后,可以在不同系列酶(細胞色素P450-含單加氧酶、過氧化物酶、酚氧化酶)的共同作用下進行代謝解毒。然而,某些IEOPs的體內轉化產物具有更高的生物毒性,或在特定條件下可以重新轉化為原母體化合物,因此其代謝產物行為需進一步關注。此外,對于某些化學性質穩定、環境持久性較強的IEOPs的體內轉化解毒過程及機制仍有待深入探究。
