長時(shí)間血清饑餓脅迫對豬骨骼肌衛(wèi)星細(xì)胞代謝和自噬的影響
2024-03-18 07:47:28范博鈞吳雅婕楊躍飛鞠輝明
中國獸醫(yī)雜志 2024年2期
高 娟,范博鈞,吳雅婕,王 怡,楊躍飛,鞠輝明
(1. 揚(yáng)州大學(xué)廣陵學(xué)院,江蘇 揚(yáng)州 225000;2.揚(yáng)州大學(xué)獸醫(yī)學(xué)院 江蘇高校動(dòng)物重要疫病與人獸共患病防控協(xié)同創(chuàng)新中心,江蘇 揚(yáng)州 225009)
骨骼肌衛(wèi)星細(xì)胞(Skeletal muscle satellite cells,SMSCs)最早由Mauro于1961年發(fā)現(xiàn),是分布于成年個(gè)體肌肉組織內(nèi),肌細(xì)胞基底膜與肌膜之間,未分化的、處于靜止?fàn)顟B(tài)的肌前體細(xì)胞[1]。當(dāng)肌肉受到損傷或者發(fā)生肌肉退化疾病而引起應(yīng)激時(shí),SMSCs可以通過肌源性分化而形成肌纖維[2]。細(xì)胞自噬通過形成自噬體,將蛋白質(zhì)和糖原顆粒等生物大分子或線粒體等細(xì)胞器回收至溶酶體,最終將其降解為氨基酸和單糖等小分子[3]。細(xì)胞自噬實(shí)現(xiàn)了能量物質(zhì)的循環(huán)再利用,是高度保守的過程[4]。研究表明,骨骼肌在饑餓狀態(tài)、廢用性萎縮、缺氧和鍛煉時(shí),其自噬通量會(huì)增加[5-7]。多個(gè)因子和信號(hào)通路已被證明有助于自噬通量的調(diào)節(jié)。其中,活性氧(Reactive oxygen species,ROS)參與自噬通量的控制,且直接與骨骼肌的再生和修復(fù)密切相關(guān)[8]。一定量的ROS對于調(diào)節(jié)細(xì)胞生長和維持細(xì)胞的各種生物學(xué)功能是必不可少的,但一些退行性疾病、骨骼肌疾病、糖尿病和衰老等會(huì)導(dǎo)致ROS平衡被破壞,進(jìn)而影響骨骼肌代謝[9]。目前,雖然有證據(jù)表明肌肉纖維恢復(fù)過程與SMSCs自噬密切相關(guān)[5,10],但自噬影響肌肉再生和修復(fù)的具體途徑和精確機(jī)制仍有待系統(tǒng)研究。本試驗(yàn)通過長時(shí)間控制SMSCs培養(yǎng)體系中的血清濃度,檢測不同程度血清饑餓脅迫對細(xì)胞代謝和自噬的影響,以期為后續(xù)探索營養(yǎng)脅迫影響豬骨骼肌發(fā)育和肌肉生成的差異機(jī)制提供理論依據(jù)。
1 材料與方法
1.1 主要試劑 DMEM高糖培養(yǎng)基,購自Hyclone公司;PBS和青霉素鏈霉素雙抗,購自美國Gibco公司;anti-LC3b和anti-p62抗體,均購自美國Abcam公司;anti-AMPK、anti-mTOR、anti-p-mTOR和anti-Tubulin抗體,均購自美國Proteintech公司;anti-p-AMPK抗體,購自美國GeneTex公司;細(xì)胞凋亡試劑盒、線粒體膜電位試劑盒、活性氧檢測試劑盒和三磷酸腺苷(Adenosine triphosphate,ATP)檢測試劑盒,均購自碧云天生物技術(shù)有限公司。
1.2 主要儀器 CO2恒溫細(xì)胞培養(yǎng)箱(Heracell 150i),購自美國Thermo公司;蛋白電泳儀(PowerPacTM Basic),購自美國Bio-Rad公司;流式細(xì)胞儀(CytoFLEX),購自美國Beckman Coulter有限公司;全自動(dòng)化學(xué)發(fā)光圖像分析系統(tǒng)(Tanon-5200),購自中國天能公司;多功能酶標(biāo)儀(BioTek Synergy),購自美國Biotek Synergy公司。
1.3 細(xì)胞來源 原代姜曲海豬SMSCs 由本實(shí)驗(yàn)室分離鑒定并保存。
1.4 細(xì)胞分組和處理 SMSCs培養(yǎng)于含20%胎牛血清的DMEM培養(yǎng)基中,隨機(jī)分為5個(gè)組:培養(yǎng)基中含20%血清的細(xì)胞為正常對照組(20%S組),培養(yǎng)基中分別含15%、10%、5%和不含血清的細(xì)胞分別為15%S、10%S、5%S和0%S組。各組細(xì)胞在不同血清濃度條件下培養(yǎng)96 h,進(jìn)行細(xì)胞代謝相關(guān)指標(biāo)和自噬相關(guān)蛋白檢測。每組3個(gè)樣本重復(fù),進(jìn)行3次試驗(yàn)重復(fù)。
1.5 流式細(xì)胞術(shù)檢測細(xì)胞凋亡率、線粒體膜電位和ROS水平 收集各組細(xì)胞,制備細(xì)胞懸液,800 r/min離心5 min,棄上清。按照試劑盒說明書操作,使用流式細(xì)胞術(shù)檢測各組細(xì)胞的凋亡率、線粒體膜電位和ROS水平。
1.6 細(xì)胞內(nèi)ATP水平測定 收集各組細(xì)胞,制備細(xì)胞懸液,800 r/min離心5 min,棄上清。按照ATP檢測試劑盒操作說明書操作,測定各組細(xì)胞中ATP水平。
1.7 Western blot檢測細(xì)胞自噬相關(guān)蛋白的表達(dá) 收集各組細(xì)胞,提取細(xì)胞總蛋白。測定各組細(xì)胞總蛋白濃度,通過聚丙烯酰胺凝膠電泳、轉(zhuǎn)印、封閉后加入一抗,包括LC3b、p62、AMPK、p-AMPK、mTOR、p-mTOR和Tubulin,4 ℃過夜振蕩孵育。洗膜后根據(jù)一抗選擇相應(yīng)的二抗,使用全自動(dòng)化學(xué)發(fā)光圖像分析系統(tǒng)進(jìn)行顯影。用Image J和Analysis軟件測定雜交條帶的灰度值。目標(biāo)蛋白條帶灰度值與內(nèi)參條帶灰度值的比值為各目標(biāo)蛋白的相對表達(dá)量[11]。
1.8 統(tǒng)計(jì)學(xué)分析 試驗(yàn)數(shù)據(jù)均用“平均值±標(biāo)準(zhǔn)差(Mean±SD)”表示,應(yīng)用GraphPad Prism 7軟件對數(shù)據(jù)進(jìn)行單因素方差分析,組間差異采用t檢驗(yàn),以P>0.05為差異不顯著,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結(jié)果
2.1 流式細(xì)胞術(shù)檢測細(xì)胞凋亡率 結(jié)果如圖1所示,20%S、15%S、10%S、5%S和0%S組SMSCs細(xì)胞凋亡率分別為(6.33±0.19)%、(7.58±0.7)%、(8.14±0.53)%、(8.51±0.3)%和(11.46±0.32)%;與20%S組相比,0%S和5%S組細(xì)胞凋亡率極顯著上調(diào)(P<0.01),10%S組細(xì)胞凋亡率顯著上調(diào)(P<0.05),15%S組細(xì)胞凋亡率差異不顯著(P>0.05)。
2.2 流式細(xì)胞術(shù)檢測細(xì)胞中線粒體膜電位 JC-1探針紅、綠熒光比率代表線粒體膜電位值,結(jié)果如圖2所示,20%S、15%S、10%S、5%S和0%S組細(xì)胞線粒體膜電位值分別為(12.72±0.55)%、(8.01±0.38)%、(6.77±0.16)%、(3.57±0.22)%和(2.81±0.04)%;與20%S組相比,0%S、5%S、10%S和15%S組細(xì)胞線粒體膜電位均極顯著下降(P<0.01)。

圖2 各組細(xì)胞線粒體膜電位檢測和分析
2.3 流式細(xì)胞術(shù)檢測細(xì)胞ROS水平 結(jié)果如圖3所示,20%S、15%S、10%S、5%S和0%S組細(xì)胞ROS水平分別為(15.87±0.42)%、(29.96±0.46)%、(31.95±0.54)%、(39.46±1.42)%和(60.68±0.42)%;與20%S組相比,0%S、5%S、10%S和15%S組細(xì)胞ROS水平均極顯著提升(P<0.01)。

圖3 各組細(xì)胞ROS水平檢測和分析
2.4 細(xì)胞內(nèi)ATP水平測定 結(jié)果如圖4所示,20%S、15%S、10%S、5%S和0%S組細(xì)胞中ATP水平分別為(4.12±0.23)、(8.01±0.3)、(9.86±0.36)、(12.64±0.55)和(23.59±2.03) nmol/L;
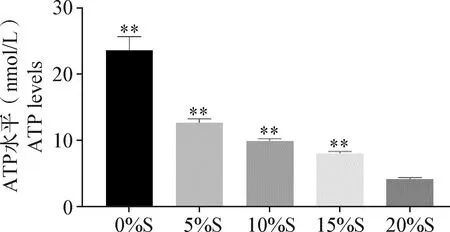
圖4 各組細(xì)胞ATP水平檢測
與20%S組相比,0%S、5%S、10%S和15%S組細(xì)胞中ATP水平均極顯著提升(P<0.01)。
2.5 Western blot檢測細(xì)胞自噬相關(guān)蛋白的表達(dá) 以Tubulin為內(nèi)參,Western blot檢測自噬標(biāo)志蛋白LC3b、p62和AMPK/mTOR信號(hào)通路相關(guān)蛋白的表達(dá)。其中,LC3b-Ⅱ與LC3b-Ⅰ蛋白條帶灰度值的比值代表LC3b蛋白的水平,通過p-mTOR/mTOR和p-AMPK/AMPK的比值變化分析AMPK/mTOR信號(hào)通路是否參與自噬的發(fā)生。結(jié)果如圖5所示,細(xì)胞中LC3b蛋白的相對表達(dá)量隨血清濃度降低而升高,p62蛋白的相對表達(dá)量隨血清濃度降低而降低。與20%S組相比,15%S、10%S、5%S和0%S組LC3b蛋白的相對表達(dá)量均極顯著上調(diào)(P<0.01);10%S、5%S和0%S組p62蛋白的相對表達(dá)量均極顯著降低(P<0.01),15%S組p62蛋白的相對表達(dá)量顯著降低(P<0.05)。p-mTOR/mTOR比值隨血清濃度降低而降低,p-AMPK/AMPK比值隨血清濃度降低而升高。與20%S組相比,10%S、5%S和0%S組p-mTOR/mTOR和p-AMPK/AMPK比值均差異極顯著(P<0.01),15%S組差異顯著(P<0.05)。

圖5 Western blot檢測細(xì)胞自噬及其信號(hào)通路相關(guān)蛋白的表達(dá)
3 討論
SMSCs對動(dòng)物個(gè)體出生后的肌肉發(fā)育、維持和再生具有重要作用,其受到損傷或者某些因素刺激后具有增殖和分化的能力,這直接決定了骨骼肌的生長發(fā)育和產(chǎn)肉性狀。SMSCs在體內(nèi)外的生長代謝需要一定營養(yǎng)能量的支撐,在體外細(xì)胞培養(yǎng)體系中添加一定濃度的血清可滿足細(xì)胞的能量需求,血清中含有細(xì)胞生長所需的生長因子、維生素、核酸衍生物、激素和其他多種未知營養(yǎng)物質(zhì)[12]。為了全面研究不同營養(yǎng)條件下動(dòng)物個(gè)體肌肉發(fā)育的差異,在細(xì)胞水平檢測SMSCs在不同營養(yǎng)條件下代謝水平的變化非常重要。細(xì)胞代謝是細(xì)胞內(nèi)用于維持生命的各種各樣的化學(xué)反應(yīng)的總稱,通過測定細(xì)胞中線粒體膜電位、細(xì)胞凋亡率、ROS和ATP水平可以較好地評(píng)估環(huán)境對細(xì)胞代謝的影響[13-16]。本試驗(yàn)結(jié)果顯示,隨著SMSCs培養(yǎng)體系中血清濃度的降低,細(xì)胞凋亡率升高、線粒體膜電位降低、ROS和ATP水平升高,這種變化趨勢與血清濃度降低的程度呈正相關(guān),說明長時(shí)間(96 h)血清饑餓脅迫能加速細(xì)胞代謝、促進(jìn)細(xì)胞凋亡,對細(xì)胞有毒害作用。本團(tuán)隊(duì)前期研究了短時(shí)間(24 h)血清饑餓脅迫對SMSCs代謝的影響,發(fā)現(xiàn)低程度的血清饑餓(15%S)在加速細(xì)胞代謝的同時(shí),對細(xì)胞凋亡影響不大,而更嚴(yán)重的血清饑餓脅迫(10%S、5%S和0%S)則會(huì)導(dǎo)致細(xì)胞代謝加快的同時(shí),加速細(xì)胞凋亡[17]。
自噬是細(xì)胞通過分解亞細(xì)胞內(nèi)容物以維持營養(yǎng)和能量動(dòng)態(tài)平衡的一種保護(hù)機(jī)制。一定程度的自噬有助于細(xì)胞的修復(fù)和再生,而過度的自噬則會(huì)對細(xì)胞功能產(chǎn)生負(fù)面影響,進(jìn)而導(dǎo)致細(xì)胞凋亡[18]。目前已有證據(jù)表明,自噬可以調(diào)節(jié)SMSCs的分化[19-20],然而不同程度的自噬可能對細(xì)胞的代謝及分化功能產(chǎn)生不同影響,最終可能成為影響動(dòng)物個(gè)體肌肉發(fā)育的重要因素,但相關(guān)工作尚未見報(bào)道。營養(yǎng)饑餓是誘導(dǎo)自噬的最典型的應(yīng)激源,本試驗(yàn)通過控制SMSCs培養(yǎng)基中的血清濃度來模擬營養(yǎng)物質(zhì)缺乏,檢測SMSCs發(fā)生自噬的程度。作為細(xì)胞自噬標(biāo)志蛋白,p62和LC3b常被用于檢測自噬發(fā)生。LC3b是自噬標(biāo)志物,自噬形成時(shí)胞漿型LC3b(即LC3b-I)會(huì)酶解掉一小段多肽,轉(zhuǎn)變?yōu)?自噬體)膜型(即LC3b-II)。p62水平升高通常標(biāo)志著自噬活性受到抑制[21-23]。本試驗(yàn)結(jié)果顯示,長時(shí)間血清饑餓脅迫后,細(xì)胞中p62蛋白相對表達(dá)量降低,而LC3b蛋白表達(dá)趨勢正好與p62相反,說明血清濃度降低能夠誘導(dǎo)細(xì)胞發(fā)生自噬,而且自噬程度與血清饑餓程度呈正比。AMPK/mTOR信號(hào)通路是細(xì)胞自噬發(fā)生的經(jīng)典信號(hào)通路之一,AMPK激活后磷酸化mTOR調(diào)節(jié)相關(guān)蛋白,進(jìn)而降低mTOR的磷酸化,mTOR的生物學(xué)功能被抑制,AMPK的活化和mTOR的抑制兩者互相協(xié)作,共同啟動(dòng)自噬[24-25]。本試驗(yàn)結(jié)果顯示,SMSCs培養(yǎng)體系中血清濃度降低后,p-AMPK/AMPK比值顯著上升的同時(shí),p-mTOR/mTOR比值顯著降低,而且變化趨勢和血清饑餓程度呈正比,表明SMSCs在血清饑餓脅迫后可通過AMPK/mTOR 通路激活細(xì)胞自噬,而且自噬的程度和血清降低的程度呈正相關(guān)。
本試驗(yàn)發(fā)現(xiàn)了長期降低SMSCs培養(yǎng)體系中血清濃度可以加速細(xì)胞代謝和凋亡,并且通過AMPK /mTOR信號(hào)通路激發(fā)細(xì)胞自噬,上述變化的程度和血清減少的程度呈正相關(guān)。本試驗(yàn)結(jié)果有助于后續(xù)研究不同程度饑餓脅迫對動(dòng)物肌肉發(fā)育的影響,探究新的動(dòng)物飼養(yǎng)模式。
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫(yī)科大學(xué)學(xué)報(bào)(2021年2期)2021-03-29 05:31:08
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
人大建設(shè)(2019年12期)2019-05-21 02:55:32
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年4期)2014-02-02 02:44:59
