微小核糖核酸-155 對肝癌細胞增殖、侵襲遷移和凋亡的影響
2024-03-13 19:24:56秦煥蓉吳祥鍇江哲宇林麗云王黎洲
介入放射學雜志 2024年1期
秦煥蓉,吳祥鍇,江哲宇,張 赟,林麗云,王黎洲,周 石
在肝細胞癌(hepatocellular carcinoma,HCC)進展過程中有多種基因改變,其中微小核糖核酸(miRNA)參與細胞分化、代謝和發育等各種生物過程,在腫瘤組織中異常表達,與腫瘤發生密切相關[1-5]。研究發現,乳腺癌患者miR-155 水平升高有利于抗腫瘤免疫特征且與更好的預后相關[6];而宮頸癌患者miR-155 水平升高與不良預后相關[7]。miR-155 在肝癌中表達上調,通過PI3K-AKT 通路抑制磷酸酶和緊張素同源物(PTEN)進而促進肝癌的進展[8]。然而miR-155 是否存在其他下游靶點影響肝癌的進展仍不清楚。本研究通過構建miR-155 沉默穩轉株以探討miR-155 對肝癌細胞增殖、侵襲、遷移和凋亡影響的機制。
1 材料與方法
1.1 細胞培養
用含10%FBS,1%青、 鏈霉素的完全培養基培養Huh7 人肝癌細胞(中國科學院細胞庫),當細胞融合度達到85%以上時進行傳代培養,3 次傳代后待細胞狀態穩定后進行檢測。
1.2 穩轉株構建
取對數增長期的Huh7 肝癌細胞1×105個接種于6 孔板,待細胞生長至30%~40%,根據慢病毒說明書中推薦的復感染指數=5,換算最佳接種病毒滴度進行轉染操作,分別轉染shNC、sh-miR-155慢病毒,轉染16 h 后進行細胞換液。待細胞長至85%左右將細胞傳代至T25 培養瓶,3~4 d 基因表達穩定后,將細胞用胰酶消化離心傳代,細胞貼壁后用含2 μg/mL 嘌呤霉素的完全培養基進行穩轉細胞株的篩選,反復藥篩。待無細胞死亡時,實時熒光定量聚合酶鏈反應(RT-qPCR)檢測病毒轉染效果,轉染成功后傳代凍存10 管穩轉株,以備后續實驗使用,在此過程中用含1 μg/mL 嘌呤霉素的完全培養基維持。陰性對照組為轉染空載病毒細胞(shNC),轉染抑制基因病毒(sh-miR-155)為沉默組。
1.3 RT-qPCR
將Blank、shNC、sh-miR-155 三組細胞分別取2×104個接種于6 孔板,待細胞長至90%左右,采用RNA 快速提取試劑盒(上海奕杉生物科技有限公司)分別提取各組RNA,反轉錄試劑盒(日本TaKaRa)將RNA 反轉錄為cDNA,RT-qPCR 得到各組Ct 值。U6 作為內參,根據公式:①ΔCt=各組Ct1(miR-155)-Ct2(U6);②ΔΔCt=各組ΔCt 值-Blank 組的ΔCt 均值;③miR-155 表達=2-ΔΔCt;④miR-155 相對表達=各組2-ΔΔCt/Blank 組2-ΔΔCt。
1.4 MTT 實驗
將Blank、shNC、sh- miR- 155、sh- miR- 155+Recilisib、shNC+Recilisib 等5 組細胞分別用胰酶消化后用完全培養基重懸,調整細胞數量為1×104個/mL。以每孔100 μL 接種于96 孔板,每組設置三個復孔,待細胞貼壁生長后,每孔加入MTT 溶液(5 mg/mL,即0.5%MTT)10 μL ,繼續培養4 h,加入結晶紫溶液,放入酶標儀A490 nm 和A630 nm 處測量各孔的吸光度值。按照公式計算細胞增殖率=[A experimental group(630 nm-490 nm)/A blank group(630 nm-490 nm)] × 100%。
1.5 Transwell 實驗
①遷移: 取各組對數生長期細胞,將細胞消化、離心后,細胞計數,每室鋪2×104個細胞,向小室下室加入含20%FBS 的高糖培養基,再輕輕向小室上室加入細胞懸液,避免出現氣泡,將培養板置于培養箱48 h 后收樣,上室細胞用棉簽擦拭,接著使用4%多聚甲醛固定30 min,0.1%結晶紫染色20 min,用水清洗后晾干,使用倒置顯微鏡于200 倍視野下拍5 個視野。②侵襲:提前將槍頭和小室放入-20°冰箱預冷,向小室上室加入基質膠使其均勻地鋪在小室,第2 天提前半小時將小室置于無血清的培養基中水化,取各組對數生長期細胞,按每室鋪5×104個細胞計算細胞懸液量,接著向小室下室加入含20%FBS 培養基。然后將小室置于24 孔板內,每孔加入無血清細胞懸液到小室上室中,細胞生長48 h 后,棄去培養基,用棉簽輕輕擦拭干凈上室中的基質膠,下室加入1 mL 多聚甲醛固定約30 min,然后用0.1%結晶紫浸泡20 min,PBS 涮洗3 遍,晾干后將24 孔板置于200 倍顯微鏡下選取5 個視野拍照并計數。
1.6 流式細胞術
將各組細胞消化、離心后,計數并調整細胞為2×105個/mL,培養體系為2 mL 接種于6 孔板,培養箱培養24 h,離心收取細胞沉淀,按照試劑盒說明書依次加入Annexin V-APC/PI 結合液,Annexin V-APC 和PI 染液混勻,輕輕吹打重懸細胞,室溫孵育10~15 min,加入預冷PBS 混勻,上機檢測各組細胞凋亡情況。
1.7 雙熒光素酶實驗
為了檢測PTPN21 與miR-155 結合,取對數生長期293T 細胞,將其隨機分為轉染miR-155模擬物組(mimicNC 組)和轉染miR-155 抑制劑組(miR-155-3p mimic 組),細胞計數,計算細胞密度為5×104個/mL 所需細胞懸液量,分別接種至48 孔細胞培養板,每個濃度梯度3 個復孔。制備轉染復合物,轉染后48 h,收集細胞檢測各組熒光素酶活性,取樣品20 μL,加入100 μL 熒光素酶檢測試劑測得相對光單位1(RLU1),完成后,加入100 μL 海腎熒光素酶檢測試劑測得相對光單位2(RLU2)。目的基因的激活程度=RLU1/RLU2。
1.8 蛋白質印跡實驗
胰酶消化細胞獲得細胞沉淀,向Western 及IP裂解液中加入蛋白酶抑制劑和磷酸酶抑制劑(Western 及IP 裂解液∶酶抑制劑=100∶1)。在冰上充分裂解30 min,每5 min 震蕩1 次,超聲破碎后通過高速離心機(12 000 r,15 min)獲得蛋白上清液,并使用BCA 試劑盒(索萊寶)配合酶標儀在562 nm 波長條件下測量細胞的蛋白濃度。在60 V(30 min)和110 V (1 h)下,采用一步法聚丙烯酰胺凝膠快速制備試劑盒(10%)(上海雅酶生物醫藥科技有限公司)凝膠電泳分離蛋白。電泳后,在4℃冷室中用濕電轉移法將蛋白轉移到聚偏二氟乙烯(PVDF)膜(德國Amersham 公司)上。然后,將PVDF 膜用5%脫脂牛奶密封,室溫孵育2 h,一抗在4℃孵育過夜。次日用含0.1%Tween-20 的Tris 緩沖鹽溶液(TBST)洗滌3 次,加入羊抗兔IgG 二抗,室溫孵育2 h,再次用TBST 洗滌3 次,將膜用ECL 曝光液浸泡在暗盒里孵育30 s左右放入凝膠成像系統中,最后再將條帶導入Image J 軟件進行各組蛋白表達水平灰度分析。
1.9 統計學分析
采用SPSS 26.0 統計軟件分析實驗數據。正態分布的計量資料以表示,多組資料比較采用單因素方差分析,組間兩兩比較用LSD-t檢驗,P<0.05為差異有統計學意義。折線圖及柱狀圖通過GraphPad 8.0 軟件制作。
2 結果
2.1 shNC 和sh-miR-155 轉染Huh7 人肝癌細胞
RT-qPCR 檢驗轉染效果,sh-miR-155 組中miR-155 的表達水平顯著低于Blank 組及shNC 組(P<0.000 1),miR-155 在Blank 組及shNC 組表達水平差異無統計學意義(P>0.05),見圖1。
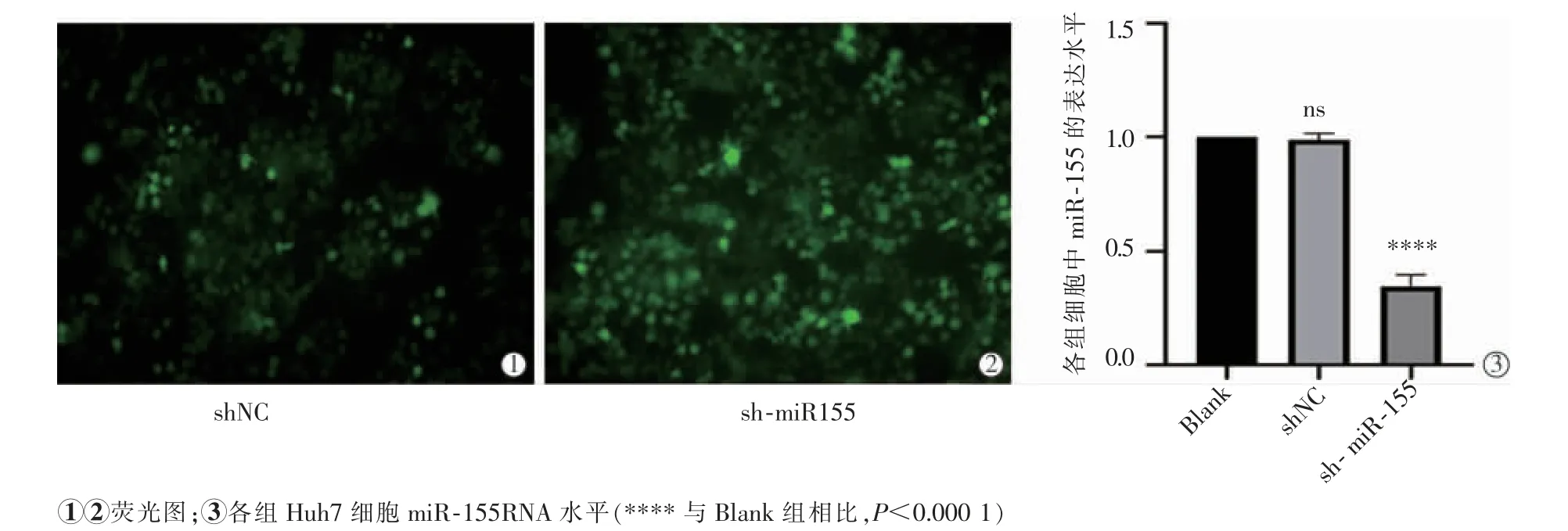
圖1 轉染shNC、sh-miR-155 后熒光圖及其轉染效率
2.2 miR-155 靶向調控PTPN21 的表達
根據microRNA 靶基因預測在線數據庫(http://www.targetscan.org/vert_70/)(http://mirwalk.umm.uniheidelberg.de/)和(http://mirdb.org/miRDB/)預測PTPN21 基因3' 非翻譯區(PTPN21-3'-UTR)上存在與miR-155 的潛在靶向結合點。雙熒光素酶實驗結果顯示,miR-155-3p 與PTPN21 有2 個結合位點,2 個位點分別設計PTPN21- 3'UTR 突變序列(PTPN21-3'UTR MUT-1)和(PTPN21-3'UTR MUT-2)。miR-155-3p 可以和PTPN21-3'UTR 野生型(PTPN21-3'UTR WT)結合使雙熒光素酶活性增強,MUT-1 位點突變后雙熒光素酶的活性也增強,說明該位點不是結合位點; 而MUT-2 位點突變后雙熒光素酶活性沒有變化,說明該位點是結合位點。通過雙熒光素酶實驗得出:miR-155-3p 可與PTPN21 結合,且結合位點是MUT-2 對應的序列,見圖2。

圖2 雙熒光素酶實驗檢測miR-155 與PTPN21 的關系
2.3 沉默miR-155 抑制肝癌細胞的增殖
MTT 結果顯示,sh-miR-155 組中Huh7 細胞在2、3、4、5 d 時A 值均低于Blank 組及shNC 組(P<0.000 1),而Blank 組與shNC 組差異無統計學意義(P>0.05);sh-miR-155 組在2、3、4、5 d 時A 值低于sh-miR-155+Recilisib 組及shNC+Recilisib 組(P=0.005 2,P<0.000 1),而sh-miR-155+Recilisib 在2、3、4、5 d 時A 值低于shNC+Recilisib 組(P<0.000 1),見圖3。
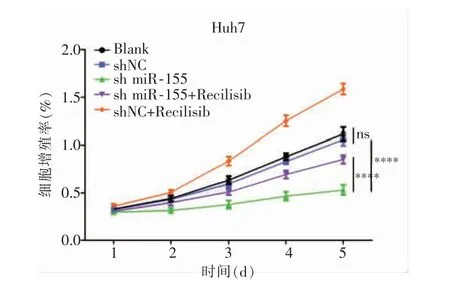
圖3 MTT 檢測各組細胞增殖能力(**** 與Blank 組相比,P<0.000 1)
2.4 沉默miR-155 抑制肝癌細胞的遷移和侵襲
Blank 組與shNC 組遷移及侵襲細胞數差異無統計學意義(P>0.05),在激活PI3K-AKT 信號通路后,與Blank 組相比,shNC+Recilisib 組肝癌細胞的遷移、侵襲能力顯著增加(P<0.000 1)。相反,沉默miR-155 后Huh7 細胞的遷移及侵襲細胞數明顯低于Blank 組及shNC 組(P<0.000 1),而PI3K 激動劑逆轉了這一現象,與sh-miR-155 組相比,sh-miR-155+Recilisib 組肝癌細胞的遷移、 侵襲能力增強(P=0.000 2),見圖4。
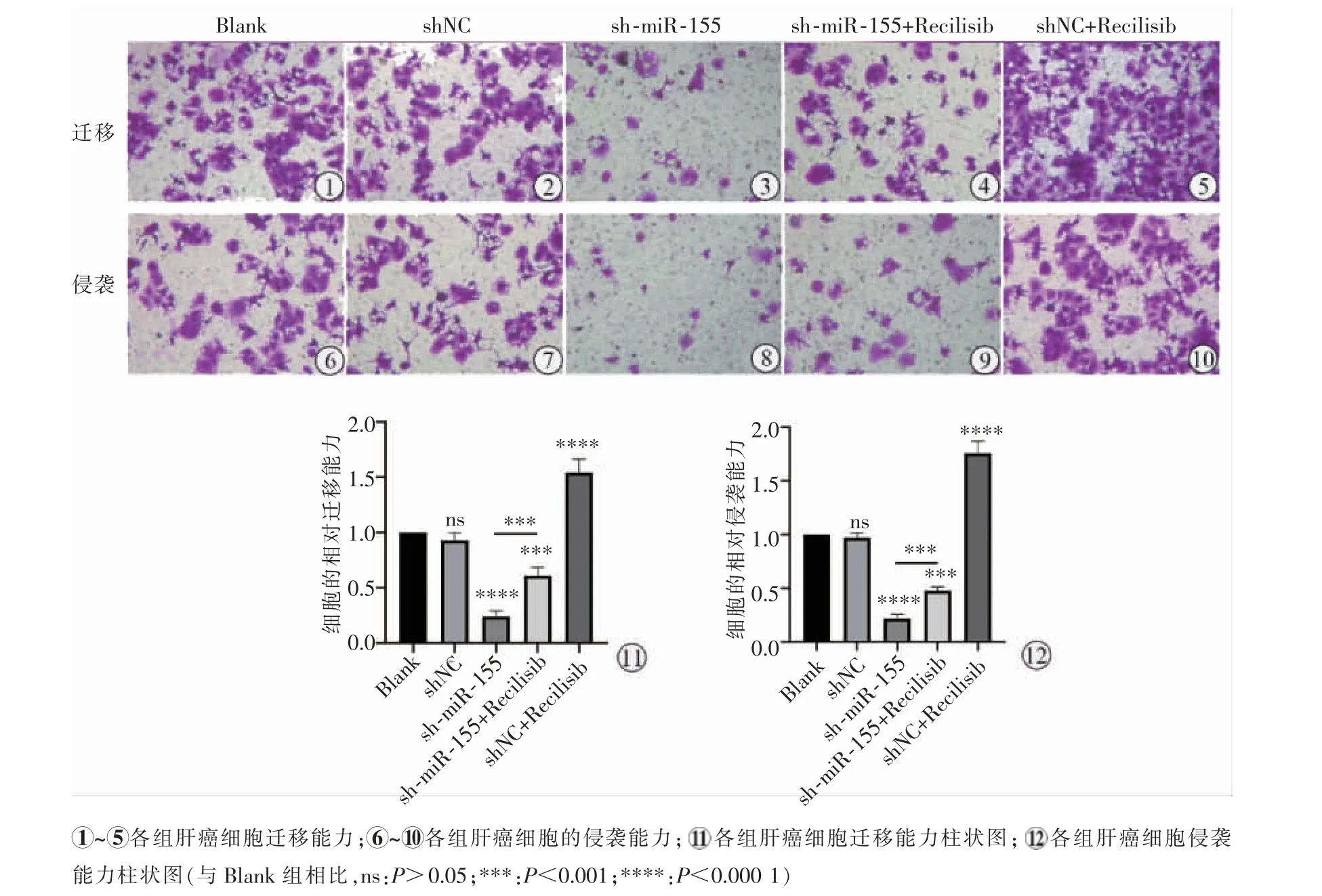
圖4 Transwell 檢測各組細胞侵襲及遷移能力
2.5 敲低miR-155 促進肝癌細胞的凋亡
流式細胞術結果顯示,Blank 組、shNC 和shNC+Recilisib 組之間細胞凋亡率差異無統計學意義(P>0.05)。與Blank 組相比,sh-miR-155 組細胞凋亡率顯著增加(P<0.000 1),并且sh-miR-155 組細胞凋亡率高于sh-miR-155+Recilisib 組(P<0.000 1),見圖5。
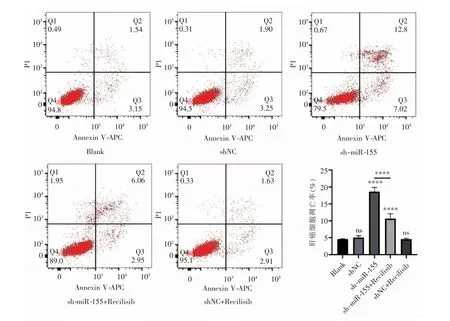
圖5 各組肝癌細胞流式細胞術圖及凋亡分析柱狀圖(與Blank 組相比,ns:P>0.05;****:P<0.000 1)
2.6 miR-155 對PTPN21、PI3K-AKT 信號通路及凋亡相關蛋白的影響
蛋白印跡實驗結果顯示,與Blank 組相比,沉默miR-155 后促凋亡蛋白Caspase-3 和BAX 表達水平顯著增加(P<0.000 1),而抗凋亡蛋白BCL2 的表達量明顯降低(P<0.000 1); 此外,sh-miR-155 組與Blank 組相比PTPN21 表達明顯下調(P<0.000 1),而在PI3K-AKT 信號通路激活時PTPN21 表達較shmiR-155 組高(P<0.000 1),但sh-miR-155+Recilisib組PTPN21 表達仍低于Blank 組(P=0.000 2);與Blank組相比,在沉默miR-155 后P-AKT 和P-PI3K 的表達顯著下降(P<0.000 1),且sh-miR-155+Recilisib 組P-AKT 和P-PI3K 表達也較Blank 組低(P=0.017 7,P=0.031 3),而sh-miR-155+Recilisib 組P-AKT和P-PI3K 表達較sh-miR-155 組高(P<0.000 1,P=0.001 4),PI3K 與AKT 在各組肝癌細胞之間的差異無統計學意義(P>0.05),見圖6。
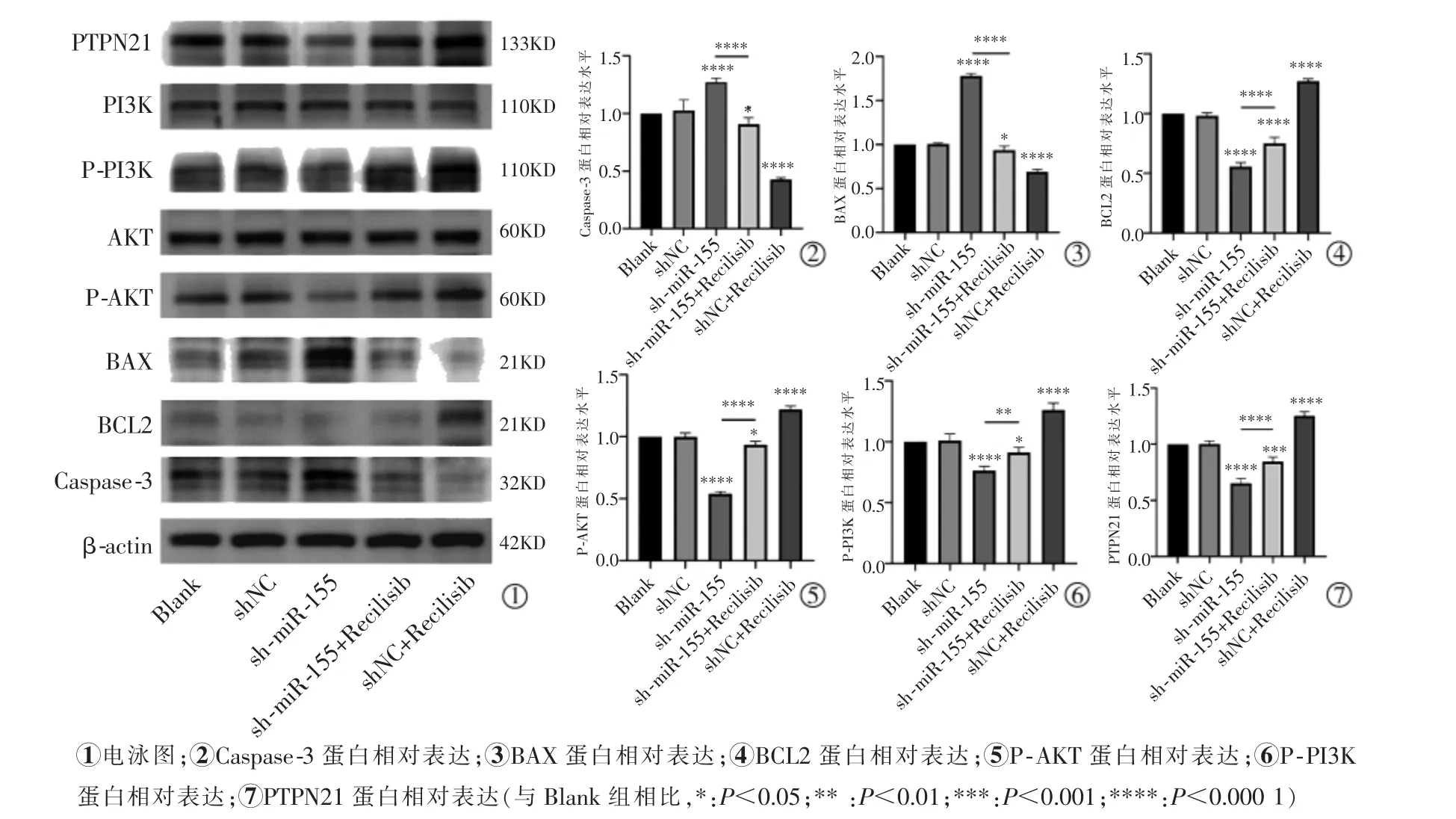
圖6 蛋白質印跡檢測PTPN21、PI3K-AKT 信號通路及凋亡相關蛋白
3 討論
研究表明,miRNA 與腫瘤的發生有密切關系,致癌相關miRNA 的過表達通過沉默腫瘤抑制因子或參與細胞分化中的某一過程,最終影響腫瘤組織血管生成、增殖及侵襲水平導致腫瘤形成[9-11]。miR-155 作為一類癌癥相關miRNA,在各種腫瘤細胞中表達量增加,如胃癌、膀胱癌、結直腸癌、乳腺癌和其他實性惡性腫瘤,并發揮不同的作用[12]。此外,miR-155 在肝癌中高表達,并與HCC 患者不良的臨床病理特征和較低生存率相關[13]。本研究通過構建miR-155 沉默穩轉株,探討下調miR-155 后對肝癌細胞增殖和侵襲遷移影響的機制。
PTPN21 在肺癌[14]、膀胱腫瘤[15]及非霍奇金淋巴瘤[16]中高表達,同時參與腫瘤細胞的生長過程,但其在肝癌中的作用機制尚不明確。本研究發現,PTPN21 基因3'-UTR 上存在與miR-155 相結合的潛在靶點。雙熒光素酶實驗結果顯示,miR-155-3p與PTPN21 有2 個結合位點,2 個位點分別設計突變序列MUT-1 和MUT-2,MUT-1 位點突變后雙熒光素酶的活性增強,而MUT-2 位點突變后雙熒光素酶活性沒有變化,說明MUT-2 是結合位點,并且miR-155 沉默可以抑制PTPN21 的表達,提示PTPN21 是miR-155 的下游靶點。
PI3K-AKT 通路是調控細胞生長、代謝、增殖、存活、轉錄和蛋白質合成的關鍵調控中心[17]。研究表明,miRNA 在調控PI3K-AKT 通路中發揮著至關重要的作用,其通過靶向PI3K-AKT 信號軸,在調節細胞增殖、凋亡和代謝方面具有重要意義[18-19]。本研究發現下調miR-155 后,各組PI3K、AKT 總蛋白變化不明顯,而磷酸化蛋白P-PI3K、P-AKT 表達量明顯下降,提示miR-155 的表達與PI3K-AKT 通路呈正相關。為了進一步驗證miR-155 對肝癌細胞生長影響是通過PI3K-AKT 通路實現的,本實驗在shNC組及沉默miR-155 的同時應用PI3K-AKT 通路激動劑,觀察PI3K-AKT 通路相關蛋白的變化情況,結果顯示,與shNC+Recilisib 組相比,miR-155 可抑制P-PI3K 及P-AKT 的表達,提示miR-155 可通過調控PI3K-AKT 通路影響HCC 生長。MTT 實驗發現,沉默miR-155 可抑制肝癌細胞的增殖。此外,流式細胞術結果顯示,與Blank 組及shNC 組相比,shmiR-155 組細胞凋亡率顯著增加,Blank 組和shNC組細胞凋亡量無顯著變化,差異無統計學意義。細胞凋亡常與多個信號傳導通路密切相關,其中BCL-2及Caspase 家族蛋白扮演著重要角色,當細胞進入凋亡途徑時,抑凋亡蛋白BCL-2 表達水平下調,而促凋亡蛋白BAX 表達上調,其下游Caspase 家族蛋白接受凋亡信號,進而激活Caspase-3 活化,執行凋亡程序[20]。本研究結果顯示,sh-miR-155 組與Blank組及shNC 組相比,抑凋亡蛋白BCL-2 表達水平顯著降低,促凋亡蛋白Caspase-3 及BAX 表達水平顯著提高,加速了肝癌細胞凋亡進程。
有研究表明,沉默PTPN21 通過調控PI3K-AKT信號通路抑制膠質瘤的進展[21]。本研究中,Transwell結果顯示,沉默miR-155 明顯抑制肝癌細胞侵襲及遷移。以上實驗表明,miR-155 通過靶向PTPN21 調控PI3K-AKT 信號通路,進而影響肝癌細胞生長,提示miR-155 在肝癌發生發展過程中發揮促癌基因的作用。
綜上所述,沉默Huh7 細胞中miR-155 通過靶向PTPN21 調控PI3K-AKT 信號通路抑制肝癌細胞生長。因此,miR-155 可能是未來HCC 患者的輔助診斷指標之一,也為臨床治療HCC 患者提供了新的治療靶點。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
海峽科技與產業(2016年3期)2016-05-17 04:32:12
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
