細胞糖酵解對豬流行性腹瀉病毒復制的影響
2024-03-11 03:24:12吳冰季霖徐雅雯唐泰山楊龍圣何召慶范紅結藺輝星
畜牧與獸醫 2024年3期
關鍵詞:檢測
吳冰,季霖,徐雅雯,唐泰山,楊龍圣,何召慶,范紅結,藺輝星*
(1. 南京農業大學動物醫學院,江蘇 南京 210095;2. 南京海關動植物與食品檢測中心,江蘇 南京 210019;3. 兆豐華生物科技(南京)有限公司,江蘇 南京 211102)
豬流行性腹瀉(PED)是一種急性腸道傳染病,主要表現為急性水樣腹瀉、嘔吐和死亡,對新生仔豬的致死率較高[1-3]。其病原豬流行性腹瀉病毒(PEDV)是單股正鏈RNA病毒,屬于套式病毒目冠狀病毒科α冠狀病毒屬[4]。病毒屬胞內寄生物,依靠宿主細胞代謝來完成其復制。病毒感染可誘導宿主細胞代謝途徑發生顯著的變化,主要包括中心碳代謝、脂肪酸合成和谷氨酰胺分解代謝等[5]。盡管病毒誘導的細胞代謝變化在某種程度上存在相似之處,但每種病毒可能需要其獨特的代謝變化才能完成其生命周期[5]。例如,甲型流感病毒H1N1、登革熱病毒(DENV)、諾如病毒(NV)和卡波西肉瘤皰疹病毒(KSHV)主要誘導糖酵解變化[6-9];而丙型肝炎病毒(HCV)和痘苗病毒(VACV)主要激活谷氨酰胺分解代謝變化[8,10]。據報道,某些冠狀病毒如新型冠狀病毒(SARS-CoV-2)可通過增強宿主細胞的糖酵解來促進自身復制[11-12],但PEDV感染對細胞代謝的影響則報道較少。本研究擬檢測PEDV感染對細胞糖酵解的影響,并驗證糖酵解變化對PEDV復制的影響,研究結果將有助于闡明PEDV的感染機制,并為冠狀病毒病的綜合防控提供參考。
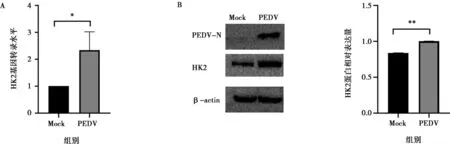
A. HK2基因轉錄水平;B. HK2蛋白表達水平;Mock為未感染IPEC-J2細胞樣品;* P<0.05,** P<0.01,下同。
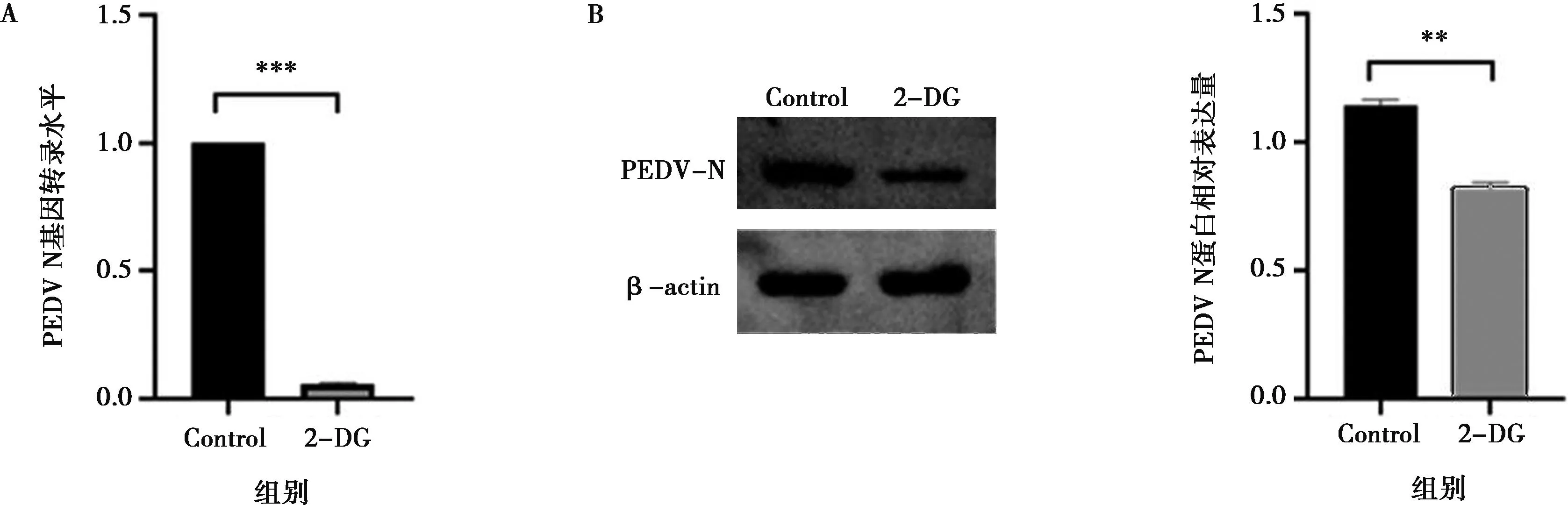
A.PEDV N基因轉錄水平;B.PEDV N蛋白表達水平;Control為PEDV感染IPEC-J2細胞36 h后收取的細胞樣品;***P<0.001。
1 材料與方法
1.1 材料
豬小腸上皮細胞IPEC-J2和PEDV流行毒株YC2014[13](GenBank:KU252649.1)均由本實驗室保存。PEDV N蛋白單克隆抗體由本實驗室制備并保存,糖酵解抑制劑2-脫氧-D-葡萄糖(2-DG)購自美國MCE公司,DMEM/F-12培養基及胎牛血清購自美國Gibco公司,RIPA裂解液、蛋白酶抑制劑PMSF購自上海碧云天生物技術有限公司,己糖激酶2(HK2)抗體購自武漢愛博泰克生物科技有限公司,鼠抗β-actin單克隆抗體購自北京全式金生物技術有限公司。
1.2 PEDV感染后細胞糖酵解的變化
1.2.1 病毒的細胞感染
IPEC-J2細胞在12孔板中培養約24 h長成單層細胞,以感染復數(MOI)為1將PEDV YC2014毒株感染細胞,孵育1 h后棄去病毒液并加入維持液。
1.2.2 實時熒光定量PCR(qPCR)
PEDV感染IPEC-J2細胞后36 h將細胞板置于超低溫冰箱,反復凍融3次,收取細胞裂解液。用TRIzol法提取細胞裂解液總RNA,然后用試劑盒將其反轉錄為cDNA。將cDNA稀釋至合適濃度后作為模板,qPCR檢測糖酵解關鍵酶HK2基因的mRNA量,以β-actin為內參基因。qPCR反應條件:95 ℃預變性30 s;95 ℃變性10 s,60 ℃退火延伸30 s,共計40個循環。每個樣本設3個重復。qPCR所用引物見表1。
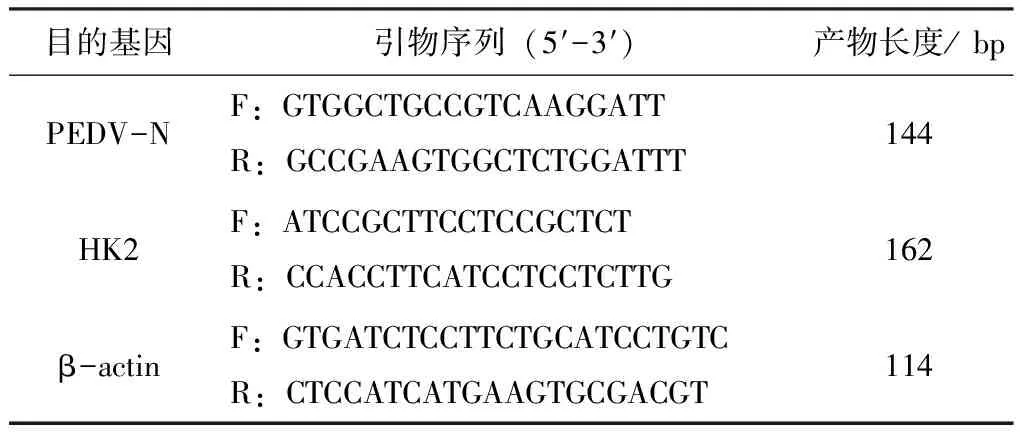
表1 引物信息
1.2.3 Western blot分析
PEDV感染IPEC-J2細胞后36 h,細胞用PBS清洗3次,每孔加入100 μL蛋白裂解液,冰上作用15 min充分裂解細胞。將細胞樣品刮下并收集到1.5 mL離心管中,在4 ℃ 12 000 r/min 離心后吸取上清液,使用BCA法測定蛋白濃度。蛋白經SDS-PAGE后轉印至PVDF膜。轉印后,用5%脫脂乳封閉2 h,PBST清洗3次。將PVDF膜分別置于脫脂乳稀釋的PEDV N抗體、HK2抗體中4 ℃過夜進行孵育。然后PBST清洗3次,將PVDF膜分別置于羊抗鼠IgG-HRP或羊抗兔IgG-HRP中,室溫孵育1 h,PBST清洗3次后加入ECL顯色液,用化學發光成像儀進行拍照。
1.3 糖酵解抑制劑對病毒復制的影響
為了研究糖酵解對PEDV復制的影響,本試驗檢測了糖酵解抑制劑2-DG對PEDV復制的影響。IPEC-J2細胞在12孔板中培養24 h,加入含有10 μmol/L 2-DG的培養基,預處理2 h后棄去培養基,以MOI為1將PEDV YC2014毒株感染細胞,孵育1 h后棄去病毒液,然后加入含有10 μmol/L 2-DG的維持液。在感染后36 h收取樣品,通過qPCR以及Western blot檢測病毒復制的變化。
1.4 不同葡萄糖濃度對病毒復制的影響
為探究葡萄糖濃度適度升高是否有利于PEDV的復制,本試驗檢測了正常濃度(5.5 mmol/L)和高濃度(11.1 mmol/L和22.2 mmol/L)葡萄糖條件下PEDV復制的情況。IPEC-J2細胞在12孔板中培養24 h,以MOI為1將PEDV YC2014毒株感染細胞,孵育1 h后棄去病毒液并加入含有正常濃度和高濃度葡萄糖的維持液進行培養。于感染后36 h收取樣品,通過qPCR以及Western blot檢測病毒復制變化。
1.5 統計分析
使用GraphPad Prism 8.0分析所有數據。組間的差異如下所示:P<0.05表示差異顯著,P<0.01和P<0.001表示差異極顯著。
2 結果
2.1 PEDV感染增強IPEC-J2細胞糖酵解
實驗室前期的非靶向代謝組學數據顯示,與對照組IPEC-J2細胞樣品相比,PEDV感染的IPEC-J2細胞中糖酵解產物乳酸的含量顯著增加,并且通過乳酸含量檢測試劑盒驗證了該結果。為了確定PEDV感染后IPEC-J2細胞中糖酵解的變化,收取PEDV感染后36 h的IPEC-J2細胞樣品,通過qPCR檢測了HK2的轉錄水平。結果如圖1A顯示,與對照組IPEC-J2細胞樣品相比,PEDV感染顯著增強了HK2的轉錄(P<0.05)。通過Western blot分析PEDV感染組和對照組的糖酵解限速酶HK2的表達,結果如圖1B所示,與對照組IPEC-J2細胞樣品相比,PEDV感染顯著增加HK2的表達(P<0.01),表明PEDV感染增強了IPEC-J2細胞的糖酵解。
2.2 糖酵解抑制劑對PEDV復制的影響
本研究進一步檢測了使用糖酵解抑制劑來抑制糖酵解是否會對PEDV的復制產生影響。己糖激酶是糖酵解途徑中的關鍵酶,2-DG是一種葡萄糖類似物,可競爭性抑制己糖激酶以抑制糖酵解。使用qPCR檢測對照組與2-DG處理組PEDV N基因的轉錄水平,結果如圖2A所示,與對照組PEDV感染的IPEC-J2細胞相比,2-DG處理顯著抑制了PEDV N基因的轉錄(P<0.001)。使用Western blot檢測對照組和2-DG處理組的PEDV N蛋白表達水平,結果如圖2B所示,與對照組PEDV感染的IPEC-J2細胞相比,2-DG處理顯著降低了PEDV N蛋白的表達水平(P<0.01),即PEDV復制受到了抑制。上述結果表明,糖酵解是PEDV復制過程中所需的關鍵代謝途徑之一。
2.3 不同葡萄糖濃度對PEDV復制的影響
由于前期研究發現PEDV感染會導致培養基中葡萄糖消耗增加,本試驗探究了在細胞維持液中適度增加葡萄糖濃度是否可促進PEDV的復制。在病毒感染后使用含有正常濃度(5.5 mmol/L)和高濃度(11.1 mmol/L和22.2 mmol/L)葡萄糖的維持液進行培養,36 h后提取細胞蛋白進行Western blot檢測,結果如圖3所示,高濃度葡萄糖條件下病毒復制顯著增加。
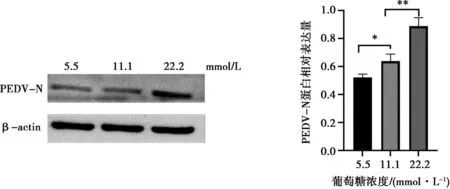
圖3 不同葡萄糖濃度對PEDV復制的影響
3 討論
病毒需要依賴細胞代謝來為病毒復制提供能量和生物大分子。許多病毒可增強細胞糖酵解,如人巨細胞病毒(HCMV)和SARS-CoV-2感染可激活細胞糖酵解,而抑制細胞糖酵解可抑制HCMV和SARS-CoV-2的復制[11,14-16]。糖酵解可促進細胞脂質的生物合成,從而促進病毒的復制[17]。DENV感染可激活糖酵解促進脂質的生物合成[18]。HCMV通過糖酵解為脂肪酸合酶(FAS)的生物合成提供原料[19-20]。本實驗室在前期的代謝組學研究中發現,PEDV感染IPEC-J2細胞后36 h,細胞中乳酸的量顯著升高。由于乳酸是細胞糖酵解的產物,乳酸濃度的升高可反映細胞糖酵解的增強。HK2是糖酵解過程中的第一個限速酶,是糖酵解的關鍵代謝酶之一[21-22]。本研究結果表明,PEDV感染導致HK2表達水平的升高,而抑制細胞糖酵解后,PEDV的復制也受到了抑制。可見,適度抑制細胞糖酵解有利于防控PEDV感染。
病毒感染對細胞糖酵解的調控機制各不相同。腺病毒通過其E4ORF1蛋白與上皮細胞中的轉錄因子Myc結合,激活糖酵解相關基因,使糖酵解相關酶的表達量升高,最終使糖酵解增強[23]。甲病毒可通過其非結構蛋白3(Nsp3)中的YXXM基序激活PI3K/AKT信號通路,從而增強細胞糖酵解[24]。SARS-CoV-2的非結構蛋白6(Nsp6)與宿主細胞中的MGA/MAX復合物相互作用從而介導糖酵解的增強[25]。PEDV感染通過何種途徑來增強宿主細胞的糖酵解有待進一步研究。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
