諾卡氏菌口服疫苗的制備與免疫效果研究
2024-03-06 10:49:38吳雅婷武尊高浩峰何學欣程浩森邵蓬
水產學雜志 2024年1期
吳雅婷,武尊,高浩峰,何學欣,程浩森,邵蓬
(天津農學院水產學院,天津市水產生態及養殖重點實驗室,天津 300384)
珍珠龍膽石斑魚(褐點石斑魚Epinephelus fuscoguttatus ♀× 鞍帶石斑魚E.lanceolatus)是一種肉質細嫩、生長速度快、抗病力強,具有很大潛在市場價值的石斑魚雜交種。研究表明,諾卡氏菌對石斑魚(Epinephelus sp.)、紅笛鯛(Lutjanus sanguineus)等主要海水養殖魚類同樣具有致病性。隨著海水種苗頻繁的流通和檢驗措施的缺乏,諾卡氏菌有蔓延和傳染其他魚類的趨勢,發病率也日益增加[6]。
目前多用抗生素防治諾卡氏菌病,但水產品質量和環境污染問題日益突出,且不同地區、不同來源和不同宿主的菌株對某些抗生素的敏感性往往差異較大,為諾卡氏菌病的治療帶來了較大的困難。諾卡氏菌病疫苗的研究大多集中在福爾馬林滅活疫苗、弱毒疫苗等。這些傳統疫苗在養殖生產中均未得到大規模的推廣和應用,且尚未見到口服疫苗防治的研究。從動物福利、方便程度、節省成本等角度考慮,口服接種都是水產動物比較理想的免疫接種方式。口服免疫一般采用拌飼口服,將疫苗口腔灌入,經胃、腸進入魚類后腸部位被吸收,引發免疫應答以研究疫苗的免疫性能。相同免疫效果下,口服免疫是水產養殖最為理想的免疫接種方式。本實驗采用海藻酸鈉天然高分子聚合物作為諾卡氏菌全菌滅活疫苗的包被載體制作口灌疫苗,分析評價了該疫苗的安全性和保護效果,意為我國諾卡氏菌病害的防控和疫苗研究提供參考。
1 材料與方法
1.1 材料
1.2 滅活疫苗的制備
1.3 口服疫苗的制備
參照薛淑霞等[7]的專利,結合諾卡氏菌的形態特征及生長特性等,選用海藻酸鈉作為口服疫苗的載體,制備NK1001 滅活疫苗微球,口灌珍珠龍膽石斑魚。操作步驟如下:
(1)將滅活的濃度為1×108CFU/mL 諾卡氏菌菌液,加入到等體積、質量濃度為3%的海藻酸鈉水溶液中,磁力攪拌器中600 r/min,10 min;
(2)將上述溶液按體積比1∶2 加入花生油,得到水油混合物,加入水油混合物體積1%的span80,1 200 r/min 攪拌15 min;
(3)加入等體積的質量濃度4%CaCl2水溶液混合,1 200 r/min 攪拌15 min,800 r/min 繼續攪拌1 h,形成油包水型乳液;
(4)3 000 r/min 離心4 min 去除上層油相和水相,留沉淀,重復兩次;
(5)加入35%酒精清洗油相,加入離心管2/3 左右后混勻,12 000 r/min,離心10 min,重復三次;
(6)按照去除的水相和油相的總體積一倍的比例加入超純水重懸,得懸濁液。
1.4 珍珠龍膽石斑魚的免疫及樣品采集
將健康珍珠龍膽石斑魚隨機分為兩組:實驗組雜交石斑魚口服疫苗微球,對照組雜交石斑魚口服生理鹽水微球,每組80 尾。考慮到免疫耐受度和口服疫苗的總量,將投喂周期定為21 d,每次口灌200 mL,灌三停四為一周,后兩次投喂可認為是加強免疫和二次加強免疫(圖1)。
分別在第一次免疫后,第二次免疫前(1 d);第二次免疫后,第三次免疫前(2 d);第三次免疫后(3 d),免疫第一輪結束后(7 d)、免疫第二輪結束后(14 d)、免疫第三輪結束后(21 d)、免疫結束后一星期(28 d),按照7 個時間點,每次分別從免疫組和對照組隨機選取9 尾石斑魚分別取出頭腎、肝臟、脾臟組織,于-80 ℃保存。
1.5 免疫保護率測定
在口服免疫4 周后進行攻毒實驗,每尾腹腔注射100 μL諾卡氏菌液,攻毒濃度為1×108CFU/mL。記錄數據,計算相對免疫保護率(RPS)。
1.6 石斑魚肝臟相關免疫指標檢測
從-80 ℃冰箱取出待檢測肝臟組織,分別測定肝臟的堿性磷酸酶(AKP)、酸性磷酸酶(ACP)、超氧化物歧化酶(SOD)、溶菌酶(LSZ)、過氧化氫酶(CAT)活性,及蛋白濃度(考馬斯亮藍法)。相關試劑配制、操作方法及計算公式參考南京建成生物工程研究所試劑盒內的說明書。
1.7 珍珠龍膽石斑魚免疫基因表達變化測定
采用實時熒光定量PCR 方法檢測免疫基因在對照組和免疫組不同時間的表達變化。將分別提取各實驗組魚頭腎、脾臟、肝臟組織中的RNA,反轉錄得到相應的cDNA,以cDNA 為模板,利用免疫相關基因的引物[8-12],進行PCR 反應。按照表1 在冰上制備熒光定量反應體系,免疫相關基因引物見表2。依據最佳反應條件,以β-actin 為內參進行熒光定量PCR,反應程序設定:95 ℃30 s;(95 ℃,5 s;60 ℃,30 s)40 個循環;95 ℃,15 s;60 ℃,1 min,95 ℃,15 s。

表1 熒光定量反應體系Tab.1 Fluorescence quantitative reaction system

表2 免疫相關基因引物Tab.2 Primers of immune related genes
1.8 數據分析
數據采用2-△△CT法分析免疫基因相對表達量的變化,運用SPSS 23.0 軟件統計學分析實驗數據,應用GraphPad Prism 8.4.3 制圖,用*表示數據差異顯著性(P<0.05)。
2 結果與分析
2.1 微球疫苗的粒徑、包封率及人工胃液腸液情況
微球成球性較高,基本呈圓形,粒徑相對均一,粒徑普遍小于50 μm,平均為(33.85±7.53)μm,外部游離菌體少(圖2)。
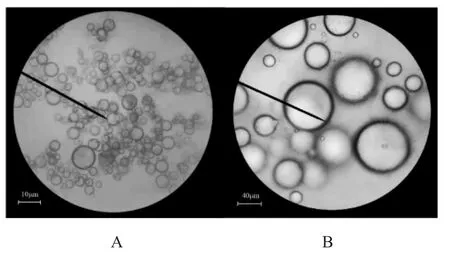
圖2 海藻酸鈉微球形態Fig.2 Form of sodium alginate microspheres
2.2 免疫保護實驗結果
免疫周期結束7 d 后,對免疫組和對照組的攻毒試驗結果表明,對照組在14 d 內全部死亡,實驗組RPS 為67%。
2.3 口服疫苗對石斑魚非特異性免疫指標的影響
石斑魚在口服微球疫苗后,ACP 活力有兩次顯著升高(圖3):口服疫苗第3 d 和第7 d;對照組在第2 d 顯著下降(P<0.05),在3 d 和7 d 極顯著升高(P<0.01),在第21 d 和28 d 顯著下降(P<0.01),其余時間點均無顯著變化;CAT 活力整體呈先升高再下降趨勢,在免疫第后1 d、2 d、3 d 和7 d 極顯著高于對照組(P<0.01),在第28 d 顯著低于對照組(P<0.05),在第14 d 和28 d 無顯著變化;LZM活力整體呈下降趨勢,第1 d 和7 d 顯著性升高(P<0.05)在第2 d、21 d 和28 d 極顯著下降(P<0.01),其余時間點無顯著變化。SOD 活力整體變化趨勢為先降低后升高再降低的趨勢,免疫后的第1~28 d 均顯著低于對照組。
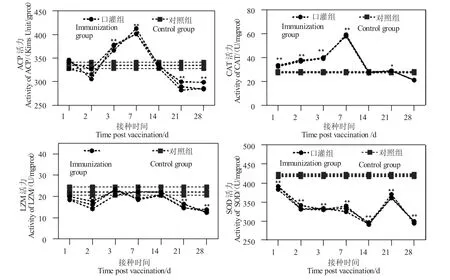
圖3 疫苗免疫后石斑魚肝臟中ACP、CAT、LZM、SOD 活性隨時間的變化Fig.3 The changes in activities of ACP,CAT,LZM and SOD in liver of the hybrid grouper after vaccination with time
2.4 口服疫苗后珍珠龍膽石斑魚部分免疫相關基因的表達
2.4.1 TLR 信號通路相關基因表達
如圖4 所示,口服疫苗免疫后,石斑魚基因TLR2 在頭腎、脾臟、肝臟組織中的表達量均有上調趨勢,在頭腎中,除第7 d 和28 d 無顯著性變化,其余時間均檢測到顯著上調;在脾臟中,除第28 d 無顯著性變化,第14 d 顯著性上調了8 倍;在肝臟中,除第2 d 和21 d 無顯著變化,第1 d、第3 d、第7 d、第14 d 和第28 d 均檢測到顯著性上調。
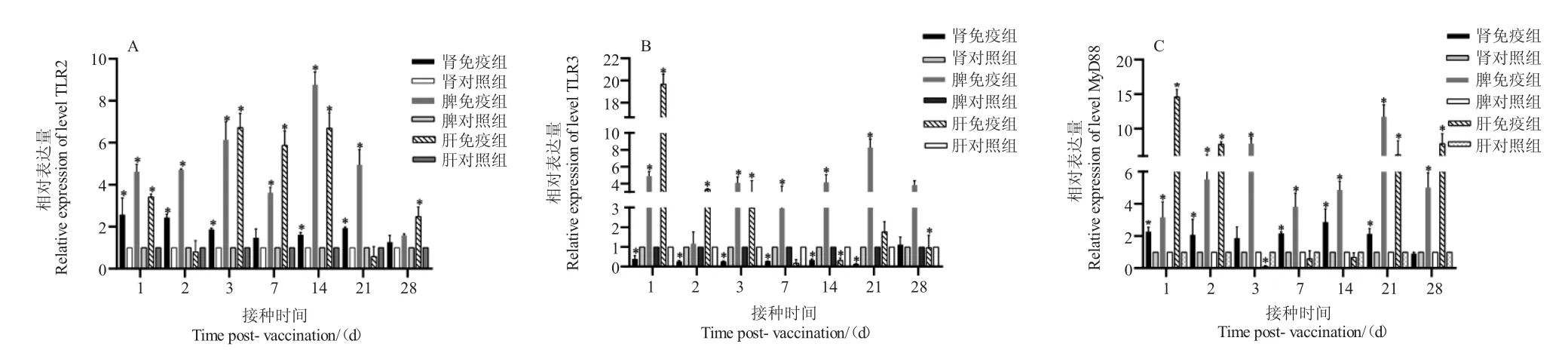
圖4 口服免疫后TLR 通路相關基因在各組織中的表達量Fig.4 The expression level of TLR pathway related genes in tissues after oral vaccination
基因TLR3 在頭腎、脾臟、肝臟中的表達量均有上調趨勢,在頭腎中,除第28 d 無顯著性變化,其余時間點均檢測到顯著性下調;在脾臟中,除第2 d 無顯著性變化,其他時間點與對照組相比均檢測到顯著性上調,第21 d 顯著性上調了8.3 倍;在肝臟中,除第7 d、21 d、第28 d 無顯著性變化,第1 d、第2 d和第3 d 均檢測到顯著性上調,分別上調19.6 倍、3.0 倍和2.9 倍,而14 d 檢測到顯著性下調。
基因MyD88 在頭腎中,除第3 d 和第28 d 無顯著性變化,其余時間均檢測到顯著性上調;在脾臟中,各個時間點與對照組相比均檢測到顯著性變化,第3 d、21 d 分別上調了7.8 倍和11.7 倍;在肝臟中,除7 d、14 d 無顯著性變化,第3 d 顯著性下調,第1 d、2 d、21 d 和28 d 顯著性上調,分別上調了14.5 倍、7.7 倍、6.1 倍和7.8 倍。
2.4.2 細胞因子類相關基因表達
口服疫苗免疫后,基因IL-12p40 在石斑魚頭腎中的表達,除第28 d 無顯著性變化,其余時間均檢測到顯著性上調,第1 d、第2 d 和第3 d 分別上調了9.3 倍、8.0 倍和5.7 倍,整體趨勢呈第一天增長隨后遞減;在脾臟中,除第2 d 無顯著性變化,其他時間與對照組相比均顯著性上調,第7 d 顯著性上調12 倍;在肝臟中,在14 d、28 d 均檢測到顯著性上調,分別上調2.2 倍和7.0 倍,其他時間與對照組無顯著性差異(圖5)。
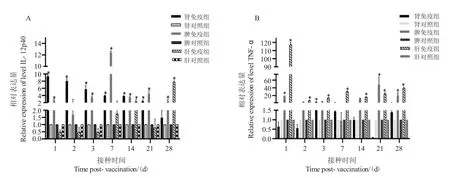
圖5 口服免疫后IL-12p40 和TNF-α 在各組織中的表達量Fig.5 The expression level of IL-12p40 and TNF-α genes in tissues after oral vaccination
口服疫苗免疫后,基因TNF-α 在頭腎中的表達,除第3 d 有顯著性變化,21 d 無變化,其余時間與對照組相比有微量上調,但與對照均無顯著性;在脾臟中,第1 d、21 d 和28 d 顯著性上調,分別為18 倍、48 倍和20 倍;在肝臟中,與對照組相比所有時間顯著性上調,第1 d 顯著性升高117.1 倍,整體趨勢先上升后下降,隨時間變化趨于穩定,在第21 d和28 d 分別顯著上調24.4 倍和38.8 倍。
2.4.3 細胞表面分子相關基因表達
口服疫苗免疫后,基因MHCII 在頭腎中的表達,在第28 d 無顯著性變化,其余時間顯著性下調;在脾臟中,除第3 d、第7 d 和第21 d 無顯著性變化,其余時間顯著上調,其中第1 d 和第14 d 分別上調了19.9 倍和18.0 倍;在肝臟中,除1 d、2 d、7 d無顯著變化,3 d、14 d、21 d 和28 d 分別顯著上調4.2 倍、7.8 倍、7.6 倍和5.5 倍(圖6)。

圖6 口服免疫后雜交石斑魚MHCII 在各組織中的表達量Fig.6 The expression level of MHCII genes in different tissues after oral vaccination
2.4.4 免疫球蛋白IgM 基因表達
口服疫苗免疫后,IgM 基因在頭腎、脾臟中的表達,各時間點均比對照組顯著性上調;在頭腎中,在第2 d 是對照組的18.9 倍;在脾臟中,在第2 d 顯著性上調15 倍;在肝臟中,除3 d、7 d 和14 d 無顯著性變化,其他時間均顯著性上調(圖7)。
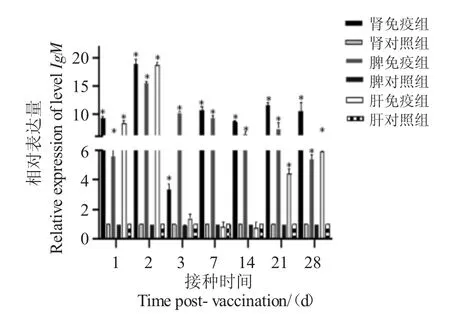
圖7 口服免疫后IgM 基因在各組織在中的表達量Fig.7 The expression level of IgM genes in tissues after oral vaccination
3 討論
非特異性免疫是魚抵擋外來病原菌至關重要的屏障,其中相關酶類的作用是魚類非特異性免疫的重要組成部分。酸性磷酸酶(ACP)是細胞免疫和體液免疫的綜合體現,反映機體對外源微生物侵染的防御力。解俊等[8]將滅活的諾卡氏菌注射入烏鱧(Channa argus)后,其血清中的酸性磷酸酶活性顯著增加。本實驗中,免疫組與對照組相比在免疫后第3 d、7 d 顯著升高,表明口服諾卡氏菌滅活疫苗使石斑魚ACP 活性顯著增強,ACP 活性升高意味著巨噬細胞被激活,提升了動物體內的應激水平,這與解俊等將滅活的諾卡氏菌對烏鱧免疫后的結果一致。
SOD 催化超氧化物自由基成為過氧化氫,再在CAT 的作用下將過氧化氫還原為水,是保護酶系統的重要組成部分。它可以增強巨噬細胞的吞噬活性,增強機體的免疫機能。CAT 在減少活性氧自由基和維持生物體內的細胞穩態方面同樣也起著至關重要的作用[14,15]。因此,常將兩種酶一起測定,其含量及活性高低間接反映養殖對象的健康狀態及生理狀況。李赫[16]等研究表明,在飼料中添加免疫增強劑,發現CAT 變化不顯著而SOD 活性顯著高于對照組。而在本試驗中,CAT 活力免疫組與對照組相比第1 d、2 d、3 d、7 d、21 d 含量顯著性升高,說明石斑魚受免疫后,增強了魚體的免疫水平,機體防御病害的能力得到提高,但SOD 活性在相應時期被抑制。這與李赫測得的血清中兩種酶活性的變化趨勢相同,分析可能是因為某些細胞受損會使胞內酶釋放入血,所以血清中的酶活性會升高。
溶菌酶主要由單核細胞和粒細胞產生,不僅能水解革蘭氏陽性細菌細胞壁中的粘肽,破壞和消除侵入體內的異物,還可改善和增強巨噬細胞的消化功能,增強機體抵抗力[17]。魚類不同組織中的溶菌酶(LZM)不僅在表達量上有差異,活性也有差異。LZM 廣泛分布在整個身體中。大馬哈魚(Oncorhynchus keta)血清、分泌物、黏膜和富含白細胞的組織(主要是腎和腸)中均能檢測到溶菌酶。何晟毓等[18]對大口黑鱸(Micropterus salmoides)免疫諾卡氏菌滅活疫苗,第7 d 其溶菌酶活力達到峰值,表明諾卡氏菌滅活疫苗可明顯增強大口黑鱸溶菌酶活力。但在本實驗中免疫組與對照組相比無顯著差異,分析原因是肝臟中LZM 的含量少,影響較小。
免疫基因的表達被作為評價疫苗效果的指標之一。Toll 樣受體(TLR)是無脊椎動物和脊椎動物中存在的單個非催化跨膜模式識別受體PRR[19]。TLR2 可以識別并對特定的革蘭氏陽性菌均能產生強烈反應,現已有大量研究證實該結論。將革蘭氏陽性枝桿菌(Mycobacterium)感染斑馬魚(Danio rerio)后,TLR2 的表達量有顯著的變化[20];烏鱧感染諾卡氏菌后,脾臟和頭腎中兩個TLR 受體基因表達均上調,且在感染后的96 h、144 h 呈持續上調;Byadgi 研究結果與其相似,諾卡氏菌感染大口黑鱸后TLR2 通路被激活[21]。
MyD88 是最先被發現的TLR 通路中的下游分子與TLR2 均為TLR 通路中的關鍵因子,不斷經過信號的傳導,最終針對不同的病原產生不同的免疫應答,兩者的表達量上和變化趨勢較為相似。本研究中TLR2 在腎臟、脾臟和肝臟中的表達均有顯著性升高;MyD88 轉錄量在各組織中都明顯上升,在腎臟中第14 d 轉錄量達到最高峰,脾臟中最高值出現在第21 d,肝臟中最高值出現在第1 d,表明MyD88 與TLR2 在不同組織及不同免疫時間表達不同。
TLR3 在魚類免疫系統調節中向T 細胞提供外源抗原,可以識別入侵病原體和激活先天免疫[22]。Tan 等[23]研究證明0.2%~1%益生元的三七提取物能夠顯著提高珍珠龍膽石斑魚肝臟內的TGF-β 和TLR3 的表達量。吳越等研究表明,甘露寡糖可以顯著提高TLR3 以及IL-8 在石斑魚腸道內的表達;菊粉和甘露寡糖可以顯著提高石斑魚頭腎中TLR3 的表達[24]。本研究結果顯示,TLR3 在頭腎、肝臟、脾臟中表達上調,說明該口服疫苗對免疫基因的上調中起重要作用,可能增強石斑魚的先天免疫力。
IL-12p40 可能在引發免疫應答中起到重要作用。張璐等[25]檢測發現,IL-12p40 在腎中的表達最高,在腦、鰓、心和肝等組織中表達量較低,表明IL-12p40 在抵御細菌侵染中起一定作用。本實驗中,第一次口灌后頭腎中的表達量顯著性上升,為對照組的9.3 倍,到第一周期免疫結束后保持一個較為穩定的狀態,但在脾和肝中表達量較低。
TNF-α 作為免疫系統中重要的免疫細胞因子之一,在機體被感染后,可調控不同免疫途徑,介導其他細胞因子的合成及釋放,以及細胞的凋亡,在魚類的免疫反應中承擔重要作用[26]。Tanekhy 等[27]將諾卡氏菌浸泡感染牙鲆(Paralichthys olivaceus)2 h 后,頭腎和脾臟中TNF-α 基因的表達量顯著增加,在24 h 后顯著降低。在本實驗中,TNF-α 均能在脾臟中高表達,在頭腎中表達量低于其他組織。TNF-α 在第一次口灌免疫后迅速上升,為對照組的117 倍,這種表達趨勢可能與它是前炎性細胞因子有關,揭示了它在機體開始發生炎癥反應中的關鍵作用。TNF-α 表達量的急速上升對于魚體迅速做出反應、抵御病原入侵具有重要意義,這些結果與先前的研究結果相似,即TNF-α 在早期表達顯著。
MHC 是一組能夠編碼主要組織相容性系統的基因群,其編碼產物廣泛參與免疫反應。在適應性免疫中,MHC 處于中心位置[28],能結合抗原并呈遞給T 淋巴細胞,刺激機體產生免疫反應。Yu 等研究表明,健康大黃魚MHC II 基因在腎、腸、鰓和脾表達較強,在肝和肌肉中表達量最弱,注射細菌疫苗24 h后,腎、腸和脾的相對表達量發生了很大變化[29]。哈維氏弧菌(V.harveyi)滅活疫苗浸泡免疫斜帶石斑魚(Epinephelus coioides)后,MHCⅡ在鰓、脾、前腎中的相對表達量顯著高于對照組[30];牙鲆注射愛德華氏菌疫苗后,脾臟中MHCⅠ、MHCⅡ均上調表達但MHCⅡ表達水平較高[31]。本研究中,免疫刺激后,MHCⅡ在各組織中均上調,脾中表達較強,但其原因與機理有待進一步研究。
IgM 主要由脾臟和淋巴結中的漿細胞分泌合成,主要參與先天免疫和體液免疫的調控。接種疫苗可提升魚類肝臟、脾臟、頭腎、胸腺組織中IgM 高表達,但表達強度與組織器官類型相關[32,33]。Nayak等[34]把亞滅活和滅活(抗原形式)的諾卡氏菌(1.2×106CFU/mL)注射進銀鯽腹腔內,3 d 后銀鯽的脾和頭腎中CD8α(+)T 細胞百分比明顯提高,血液中IgM抗體數量也明顯增加,接種15 d 后銀鯽體內抗體水平達最高,隨后下降,存活率分別為62.5%和75.0%。本實驗結果表明,口服疫苗免疫后,實驗組與對照組相比各組織IgM 的表達水平均提升,頭腎、脾臟、肝臟均在免疫后第2 d 達到峰值,IgM 表達量與組織差異相關。本研究中,口服疫苗使石斑魚頭腎、脾臟、肝臟組織中TLR2、TLR3、MyD88、TNF-α、IL-12p40、MHCII 和IgM 的表達部分顯著上調也存在顯著性下調,不同組織間表達量存在差異,說明制備的口服疫苗有效地引起了免疫反應,且大部分基因參于了免疫應答。
