呼倫湖流域魚類群落結構特征及與物理生境特征關系研究
2024-03-02 07:55:00李朝暉王鵬偉包薩茹徐德琳陳星瑤
生態與農村環境學報 2024年2期
關鍵詞:物理
李朝暉,單 楠①,王鵬偉,包薩茹,龐 博,徐德琳,陳星瑤②
(1.生態環境部南京環境科學研究所,江蘇 南京 210042;2.國家環境保護呼倫湖濕地生態環境科學觀測研究站,內蒙古 呼倫貝爾 021000;3.呼倫貝爾市北方寒冷干旱地區內陸湖泊研究院,內蒙古 呼倫貝爾 021000)
魚類在水域中占據較高營養層,其群落結構狀況可反映內源性、外源性因素對魚類資源及水域生態系統狀況的影響。魚類通過“上行效應”與“下行效應”參與水域生態系統的物質循環與能量流通,是水生態系統提供生態服務的核心[1]。魚類群落結構穩定性關乎水生態系統的穩定性及演替方向,魚類生物多樣性保護在水域生態環境修復與生物多樣性保護工作中占有重要地位[2]。魚類群落是魚類種群相互結合的一種結構單元,與周圍環境及其他物種相互依賴、相互作用,會隨水環境變化發生改變[3]。魚類對水生態系統的食物網動態、營養和物質循環具有重要的調節作用[4-5],在一定程度上可反映水生態系統的恢復能力[6]。因此,保持魚類群落結構穩定是水生態系統恢復的關鍵之一[7]。
魚類群落結構及其多樣性受物理生境特征影響較大。魚類棲息生境的改變會影響魚類的生存與繁衍[8],甚至可能造成部分魚類功能類群的減少或消失[9]。物理生境是生境的重要組成部分,是指影響水生生物群落結構和功能的物理結構特征,對維持水生態系統完整性和水生態系統健康具有重要作用[10-11]。水體污染物在水體中遷移運輸的重要影響因素之一為水體物理生境質量狀況,如土壤、地形、水文和植被等[12-13]。物理生境改變已成為水生態系統與水資源退化的主要脅迫因子之一[14]。遙感技術可以獲取大范圍地表信息,具有多時相、數據客觀、覆蓋全面等特點,可用于定量分析河流與湖泊的水面范圍、水面形態、地勢、植被覆蓋等物理生境指標[15]。例如,利用多源衛星遙感影像數據提取水面面積變化[16],計算河道岸線分形維數[17-20],反演水位[21],進行濕地監測[22]等。遙感技術不僅可以高效完成復雜生境的調查工作,還可以克服大面積調查中人工調查的局限性,在流域生境調查中發揮重要作用[23]。
呼倫湖作為我國北方寒冷干旱地區典型內陸湖泊,在維持呼倫湖流域生態系統平衡和生物多樣性方面發揮著巨大作用[24-25]。呼倫湖漁業資源的系統調查自20世紀80年代初開始[26],調查研究內容包括魚類資源組成、魚類種屬、魚類區系的演化以及生物學特征等[27]。近年來,呼倫湖魚類研究主要集中在魚類食性[28]、繁殖生物學[29]、魚類營養成分[30]和魚類群落結構現狀[31-32]等方面,且魚類影響因素方面多集中于水質環境[33-34],物理生境特征對魚類群落結構及其物種多樣性的影響研究仍有限。
為了提高對呼倫湖魚類群落結構特征的認識,該研究利用衛星遙感技術探討枯水期和豐水期呼倫湖流域物理生境、魚類群落結構空間分布特征及功能群的差異,通過聚類分析和冗余分析明確物理生境因子對魚類群落結構的影響程度,以期為呼倫湖魚類資源保護及其生境恢復提供參考。
1 材料與方法
1.1 研究區概況
呼倫湖位于呼倫貝爾草原西部,湖水來源主要包括克魯倫河、烏爾遜河、呼倫溝、海拉爾河等主要入湖河流。氣候屬于中溫帶大陸性草原氣候,年平均氣溫為1.2 ℃,年均降水量為235 mm,年均蒸發量達1 470 mm[35]。呼倫湖地勢呈中心低、四周高的特點,且呼倫湖北岸海拔高于南岸。
1.2 魚類數據獲取與鑒定
分別于2021年枯水期(5月)和豐水期(7—8月)對呼倫湖流域進行了2次魚類調查采樣。枯水期設采樣點18個,豐水期設采樣點14個,采樣點設置如圖1所示。魚類樣品主要通過人工捕撈方式進行采集,在流動性較高的河流采用下網捕撈方式,在流動性較低的湖泊則采用抄網捕撈方式。參照文獻[36]中的監測方法,調查網具為歐洲標準采樣網具——復合網目刺網(2 m×30 m,由12種網目類型組成,每種網目網長2.5 m、網高3 m)和底層定置網具(俗稱“地籠”,0.5 m×0.5 m×15 m)。現場對采集到的魚類樣品進行物種鑒定和生物學指標測量,樣品鑒定與分類主要依據文獻[37-39]。
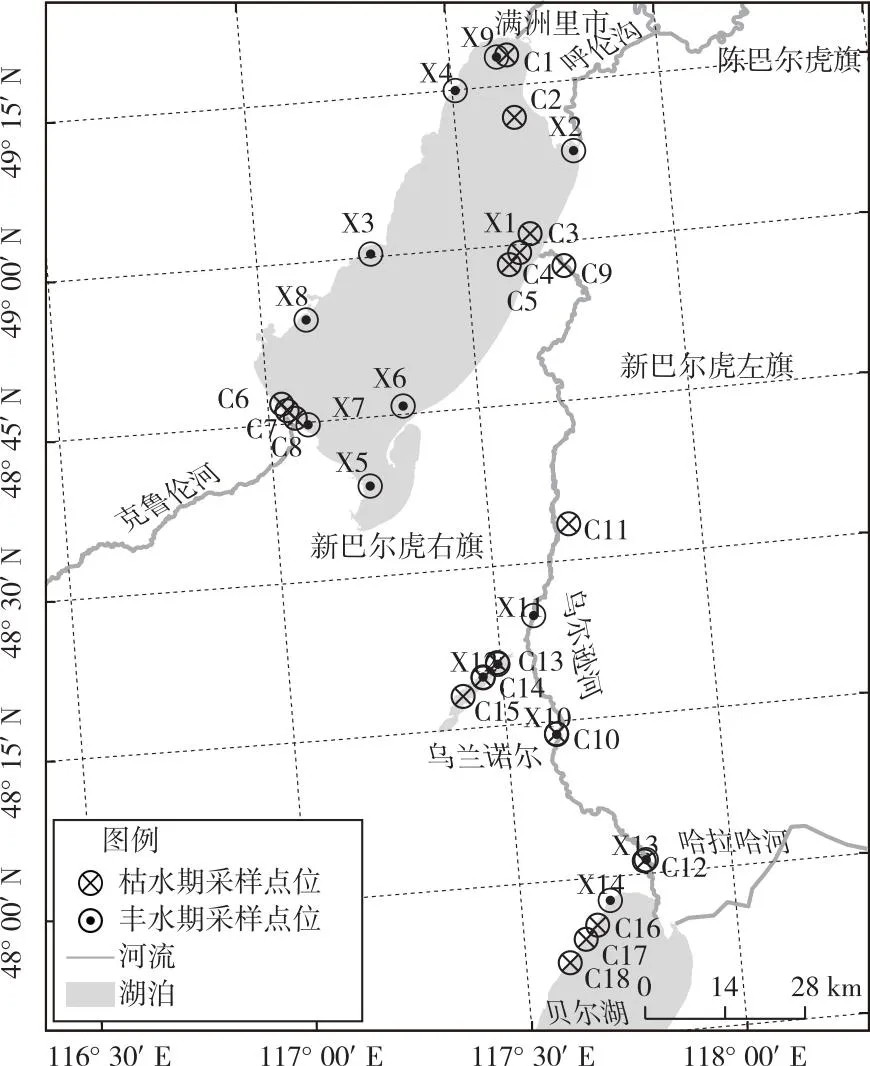
圖1 2021年呼倫湖流域枯水期和豐水期魚類采樣點位分布
1.3 遙感影像獲取
基于Google Earth Engine獲取2021年呼倫湖流域5月和8月的Sentinel-2反射率影像數據,分別對應枯水期和豐水期采樣時間。每景圖像采用Sentinel-2去云算法去云,并鑲嵌多期影像用于水體邊界提取,空間分辨率為10 m。呼倫湖湖底地形數據采用美國地質調查局(USGS)和國家地理空間情報局(NGA)共同制作的GMTED2010數據集,空間分辨率為900 m。
1.4 生境指標計算
考慮到呼倫湖流域面積大、點位組群間距離遠等因素,為了方便統計和分析,以采樣點為中心,構建邊長為10 km的正方形格網,分別計算每一個格網內的物理生境特征參數,包括分維數(fractal dimension,FD)、水面面積(Area)和高程值(digital elevation model, DEM),作為呼倫湖物理生境特征評估參數。
分維數又稱分形維數,是對非光滑、非規則、破碎等分形體進行量化描述的重要參數,表征分形體的復雜程度。分維數值越大,表示格網內水域邊界形狀越復雜、越不規則;反之,分維數值越小,表示格網內水域邊界形狀越簡單、越規則。研究采用盒計法計算呼倫湖湖區分維數。盒計法是 GRASSBERGER[40]設計的一種分維計算方法, 基本思想是使用不同長度的正方形網格覆蓋被測線體,當正方形網格長度ε出現變化時,覆蓋有被測線體的網格數目N(ε) 就會出現相應的變化, 根據分形理論得到:
N(ε)∝ε-D。
(1)
當正方形網格長度為ε1,ε2,…,εk時, 則覆蓋有被測線體的正方形網格數目為N(ε1),N(ε2),…,N(εk), 對上式兩邊取對數得下式:
lgN(ε)=Dlgε+A。
(2)
式(1)~(2)中,A為截距;D為被測線體的分維數,其值等于該式斜率值的絕對值。
水面面積表示格網內的水域面積,反映了采樣點位附近的水域范圍大小。高程值表示格網范圍內的平均高程值,反映區域地形地勢。分維數、水面面積和高程值均利用Python 3.7.9 完成計算。
1.5 魚類優勢種與群落多樣性
采用相對重要性指數( Index of Relative Importance,IRI,IIR)對魚類群落優勢種進行度量,其計算公式為
IIR=(N+W)×F×104。
(3)
式(3)中,N為某物種數量占總數的比例;W為某物種質量占總質量的比例;F為此物種在調查中出現的次數占總調查次數的比例。該研究將IRI值>2 000 的物種確定為群落優勢種。
采用Shannon-Wiener指數(H′)、Margalef種類豐度指數(D)和Pielou 均勻度指數(J′)進行魚類群落多樣性分析,計算公式為
(4)
D=(S-1)/lnN,
(5)
J′=H/log2S。
(6)
式(4)~(6)中,S為采集魚類樣本的總物種數;Pi為i物種樣本數量的比例;N為樣本總數。
1.6 聚類分析、相似度與冗余分析
聚類分析用于劃分魚類群落,根據魚類采樣結果,依據魚類在各采樣點中有無分布,有分布則編碼為“1”,無分布則編碼為“0”,建立采樣點位-魚類分布矩陣[23,41]。以采樣點位為分類單元,以魚類有無分布為性狀進行聚類分析,距離系數為Jaccard距離,聚類方法為層次聚類法[42-43]。物理生境聚類分析則利用枯水期和豐水期衛星遙感影像計算,根據分維數、水面面積和高程值等計算結果,建立采樣點位-物理生境矩陣。以采樣點位為分類單元,以歸一化后的物理生境為性狀進行聚類分析,距離系數為Euclidean距離,聚類方法為層次聚類法。采用相似性度量(similarity measurement)描述魚類群落聚類與生境聚類相似性。2種事物越接近,其相似性度量越大;反之,相似性度量越小[44]。另外,選擇冗余分析(RDA)對多環境指標進行統計檢驗,確定對魚類群落空間分布格局和多樣性具有顯著影響的生境指標,探究不同物理生境因子對呼倫湖流域魚類群落結構的影響。利用ArcGIS 10.8軟件生成采樣點格網,利用Python 3.7.9對魚類采樣數據和物理生境數據進行聚類分析和RDA分析。
2 結果與分析
2.1 呼倫湖流域物理生境特征
2021年呼倫湖流域魚類采樣點位覆蓋呼倫湖、貝爾湖、烏蘭諾爾、烏爾遜河及主要河流入湖河口。對枯水期采樣點所在網格的分維數、水面面積和高程值進行聚類分析,在聚類距離為1.1水平上,2021年枯水期18個采樣點位可以分為4個組群,分別記為Ac(C2、C4)、Bc(C8、C10、C9、C6、C7、C3、C13、C5)、Cc(C14、C15、C12、C17、C18)和Dc(C16、C1、C11)組(圖 2)。對豐水期采樣點所在網格的生境指標進行聚類分析,在聚類距離為1.0水平上,14個采樣點位可以分為4個組群,分別記為Ax(X14、X6)、Bx(X8、X7、X3、X4)、Cx(X11、X12、X13)和Dx(X2、X5、X9、X1、X10)組(圖2)。
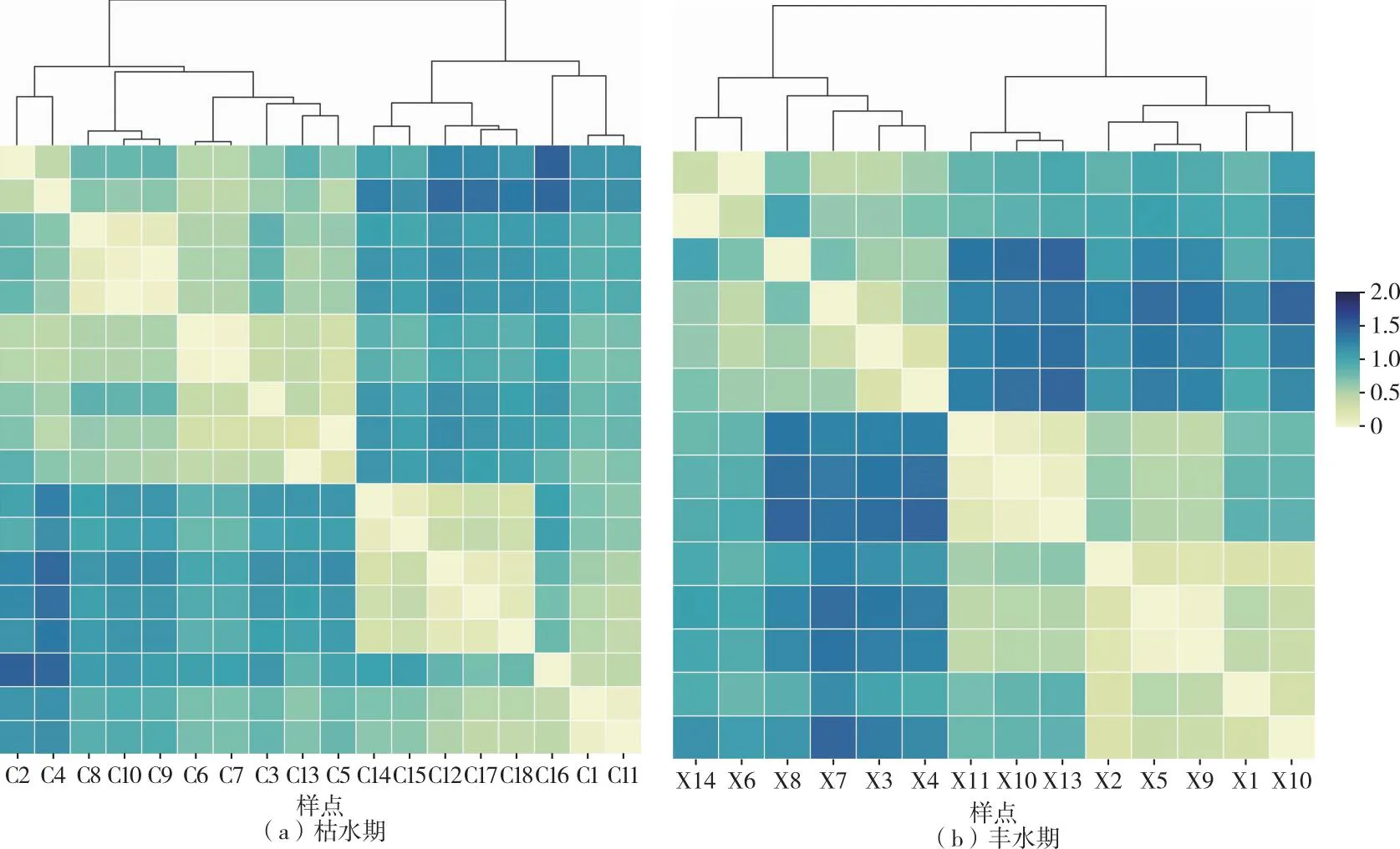
圖2 2021年呼倫湖流域枯水期和豐水期物理生境的聚類分析
2021年,呼倫湖流域枯水期采樣點位物理生境指標的平均值與豐水期相近,并呈現相似規律。枯水期呼倫湖流域各采樣點位分維數、水面面積、高程值平均值分別為1.17、59.72 km2和559.58 m,豐水期則分別為1.17、58.51 km2和555.00 m,變化率絕對值均小于2%。枯水期物理生境聚類組群的水面面積隨高程值降低而升高,這一規律在豐水期也有體現(圖3)。
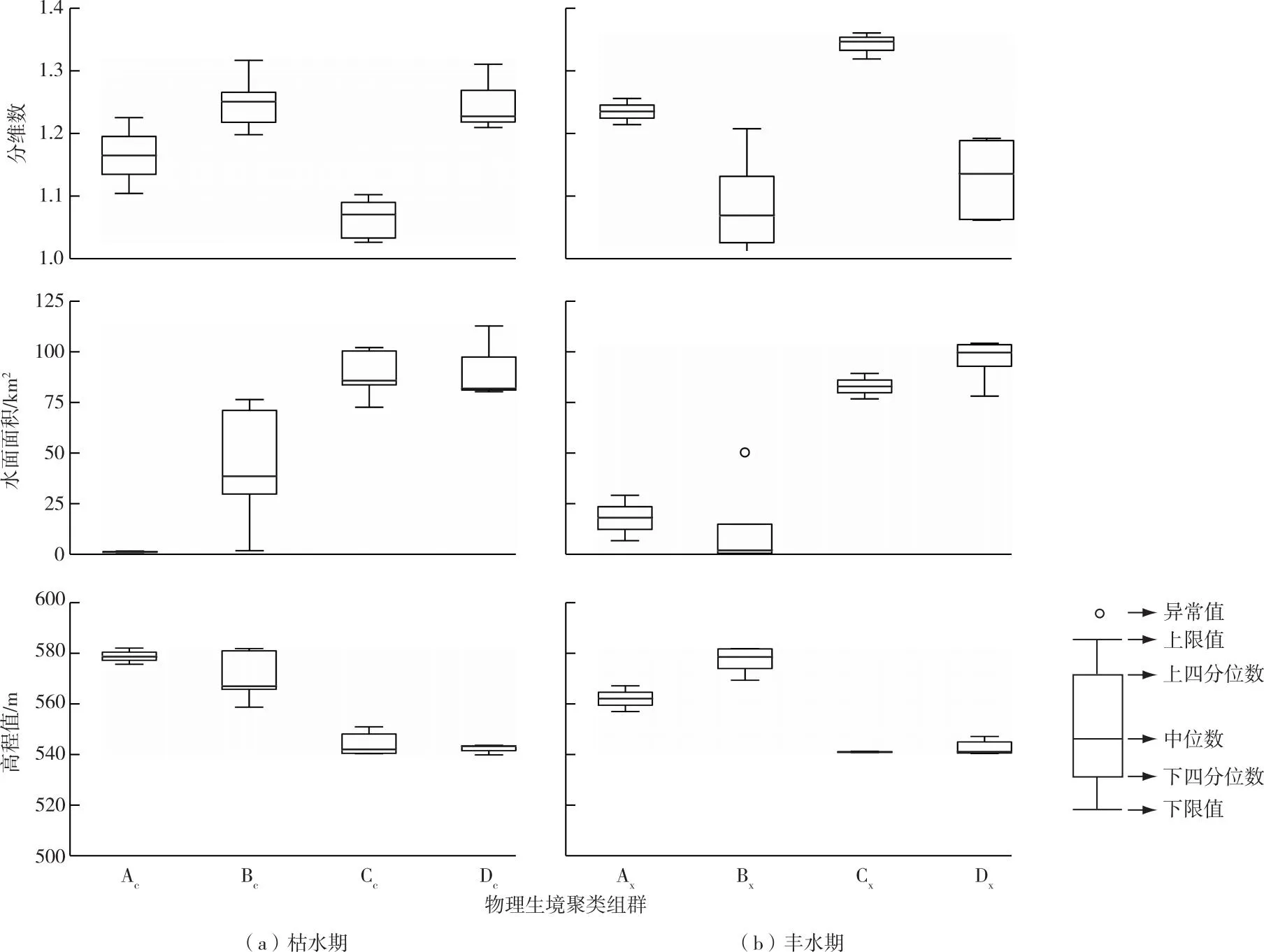
圖3 2021年呼倫湖流域枯水期和豐水期物理生境聚類組群生境指標對比
枯水期物理生境聚類4個組群空間分布規律明顯。Ac組群主要包含呼倫溝河和烏爾遜河河口,岸線分維數均值為1.15,岸線較平緩;平均高程值為578.61 m,地勢相對較高,水深較低;平均水面面積較小,僅為1.16 km2。Bc組群主要分布在克魯倫河入湖河口處以及烏爾遜河,平均岸線分維數高于1.2,岸線較曲折;平均高程值為570.72 m,地勢相比Ac組群更低;平均水面面積也升高為44.25 km2。
Cc組群則主要分布在烏蘭諾爾和貝爾湖等湖區,岸線分維數均值僅1.05,岸線平緩;同時具有較低的地勢和更寬闊的水面,平均高程值和水面面積分別為544.48 m和88.80 km2。Dc組群分布規律不明顯,在新開河河口、烏爾遜河中段以及貝爾湖湖區均有分布。Dc組群具有較高的岸線分維數(1.23),岸線較曲折,但平均高程值較低(542.40 m),也具有較寬闊的水域范圍(91.52 km2)(圖3)。
豐水期采樣點聚類組群也有較明顯的空間分布規律。Bx組群主要分布在呼倫湖北岸和克魯倫河河口處,岸線較平緩,平均分維數低于1.1,地勢最高,平均高程值為577.04 m,但水面面積最小,僅為13.75 km2。Cx組群主要分布在烏蘭諾爾湖區和烏爾遜河,具有最高的平均分維數(1.33),岸線曲折,但地勢較低,平均高程值為541.08 m,平均水面面積較高,為83.09 km2。Dx組群則主要分布在呼倫湖東岸和南岸的河流入湖河口處,如新開河河口、呼倫溝河口、烏爾遜河河口等。該組群岸線較簡單,地勢也相對較低,平均水面面積也較高。Ax組群在呼倫湖和貝爾湖均有分布(圖3)。
2.2 呼倫湖流域魚類群落結構

呼倫湖流域魚類聚類分析結果(圖4~5)表明,2021年枯水期在聚類距離為0.8水平上,18個采樣點位可以分為4個組群,分別記為Ac′(C10、C1、C9)、Bc′(C6、C7、C5、C3、C4)、Cc′(C2、C12、C17、C18、C11、C14)和Dc′(C16、C8、C13、C15)組。2021年豐水期在聚類距離為0.5水平上,14個采樣點位可以分為4個組群,分別記為Ax′(X9、X6、X7)、Bx′(X8、X3、X4)、Cx′(X11、X14、X10、X13)和Dx′(X2、X1、X12、X5)組(圖4)。
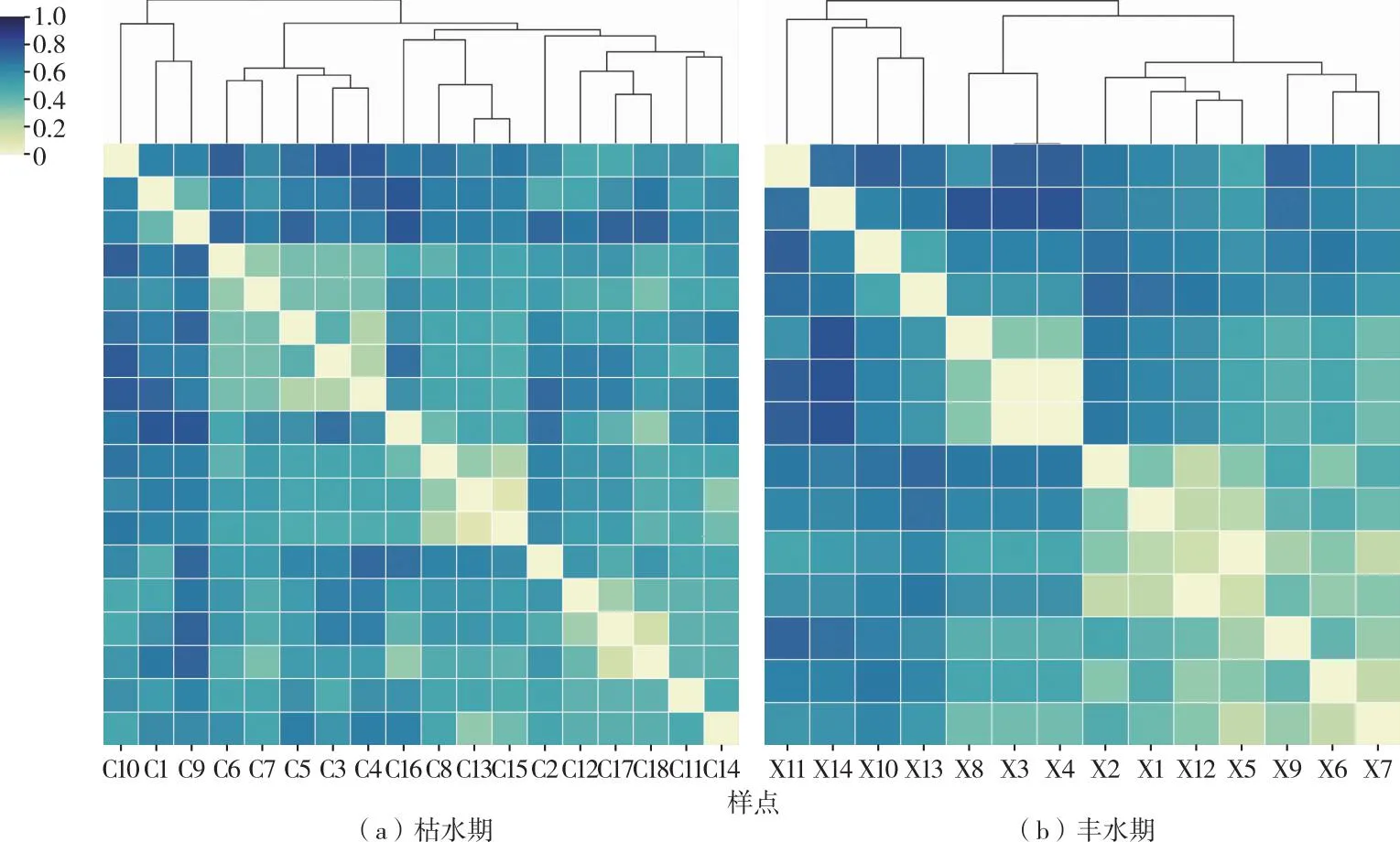
圖4 2021年呼倫湖流域枯水期和豐水期魚類群落聚類
枯水期,Bc′組群主要位于烏爾遜河河口和克魯倫河河口,Shannon-Wiener指數、Margalef豐富度指數、Pielou均勻度指數平均值分別為0.70、0.88和0.25,具有最低的魚類多樣性和豐富度。Cc′組群主要集中于貝爾湖和烏爾遜河,Shannon-Wiener指數、Margalef豐富度指數、Pielou均勻度指數平均值分別為1.94、1.64和0.56,魚類多樣性、豐富度和均勻度相比于Bc′組群均有所增加。
Dc′組群主要集中于烏蘭諾爾和貝爾湖,其與Cc′組群空間分布相似,但魚類多樣性和均勻度均有提高,Shannon-Wiener指數和Pielou均勻度指數平均值分別增加至2.02和0.58,但魚類豐富度降低至1.52。Ac′組群空間分布較離散,無明顯特征,但具有最高的魚類多樣性、豐富度和均勻度(圖 5)。豐水期采樣點魚類群落聚類組群也具有較明顯的空間分布規律。例如,Ax′組群主要位于新開河和克魯倫河入湖河口處,Shannon-Wiener指數、Margalef豐富度指數、Pielou均勻度指數平均值分別為1.19、1.13和0.37。Bx′組群主要分布在呼倫湖北岸,Shannon-Wiener指數、Margalef豐富度指數、Pielou均勻度指數平均值分別為0.93、0.58和0.40,魚類群落豐富度明顯低于Ax′組群。Cx′組群主要分布在貝爾湖及與之相通的烏爾遜河南段,Shannon-Wiener指數、Margalef豐富度指數、Pielou均勻度指數平均值分別為2.26、1.81和0.75,具有豐水期4個組群中最高的魚類多樣性、豐富度和均勻度。Dx′主要位于呼倫溝入湖河口、烏爾遜入湖河口以及烏蘭諾爾等區域,Shannon-Wiener指數、Margalef豐富度指數、Pielou均勻度指數平均值分別為2.04、1.65和0.56,魚類群落結構劣于Cx′組群,但優于Ax′和Bx′組群(圖5)。

圖5 2021年呼倫湖流域枯水期和豐水期魚類群落聚類組群群落多樣性指標對比
2.3 呼倫湖流域物理生境與魚類群落空間聚類相似度
2021年呼倫湖流域物理生境和魚類群落空間聚類組群空間分布具有較高相似性,枯水期和豐水期魚類群落聚類和物理生境聚類結果平均相似度分別達41.29%和63.06%(表1、圖6~7)。
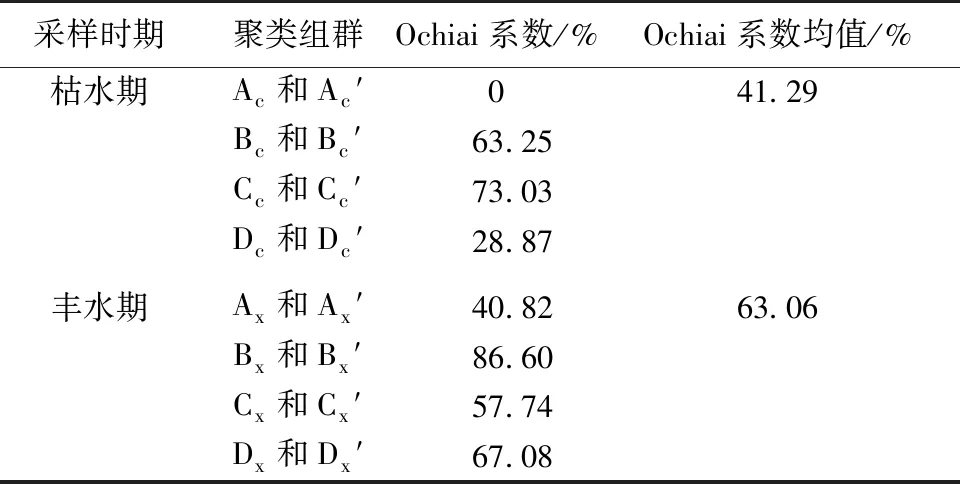
表1 2021年枯水期和豐水期的魚類群落聚類和物理生境聚類結果相似度
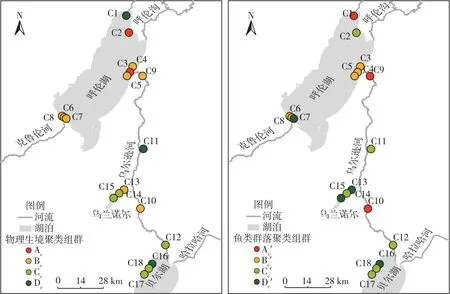
圖6 2021年呼倫湖流域枯水期物理生境和魚類群落聚類組群空間分布
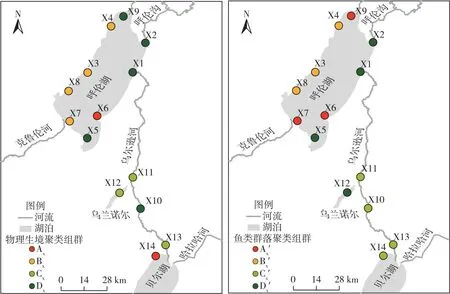
圖7 2021年呼倫湖流域豐水期物理生境和魚類群落聚類組群空間分布
其中,枯水期呼倫湖流域物理生境聚類Bc、Cc組群和魚類群落聚類Bc′、Cc′組群的Ochiai系數分別達到63.25%和73.03%(表1),組群相似度較高。豐水期呼倫湖流域物理生境和魚類群落聚類組群相似度較枯水期有所提升。物理生境聚類的Bx、Cx和Dx組群和魚類群落聚類的Bc′、Cc′和Dx′組群的Ochiai系數分別達86.60%、57.74%和67.08%。
2.4 呼倫湖流域魚類群落結構和物理生境因子的關系
魚類群落多樣性與物理生境指標的RDA分析結果如圖8所示。呼倫湖流域魚類群落多樣性均受分維數、水面面積和高程值(DEM)的影響,不同時期影響魚類群落多樣性的主導生境因子存在差別。枯水期主要受水面面積和DEM的影響,多樣性指數與水面面積呈顯著正相關關系,與DEM呈顯著負相關關系(表2)。豐水期受DEM影響較大,其中多樣性指數H′、豐富度指數D和均勻度指數J′均與DEM呈顯著負相關關系(表2)。
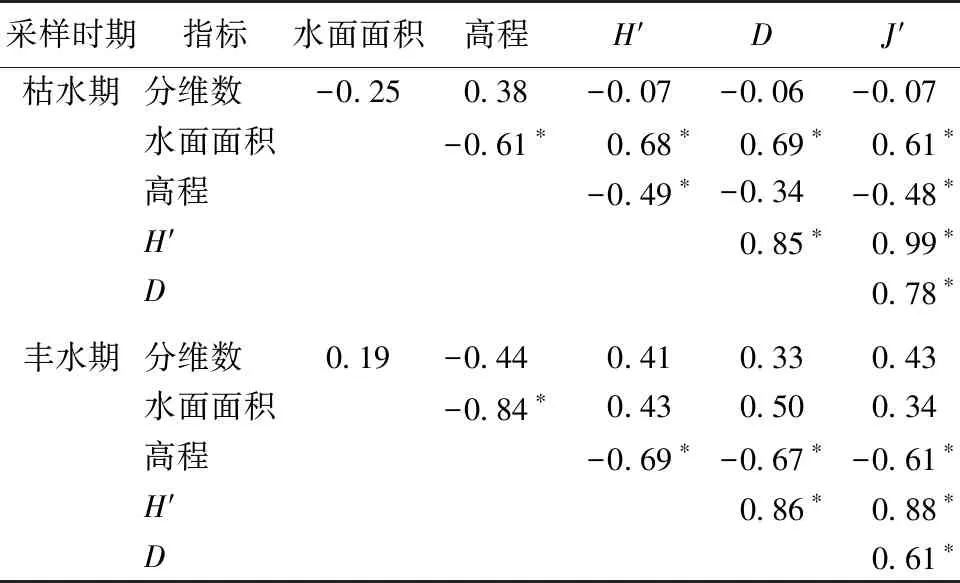
表2 多樣性指數H′、豐富度指數D、均勻度指數J′與物理生境指標的Pearson相關性分析
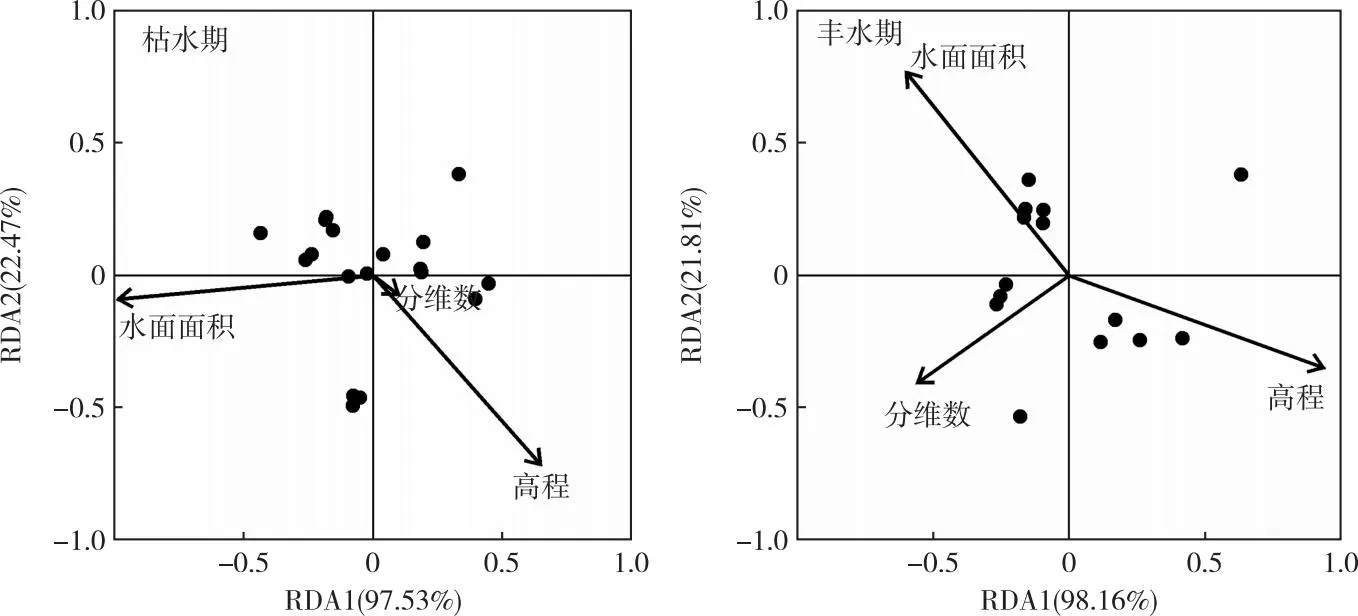
圖8 呼倫湖流域枯水期和豐水期魚類群落多樣性與物理生境的RDA分析結果
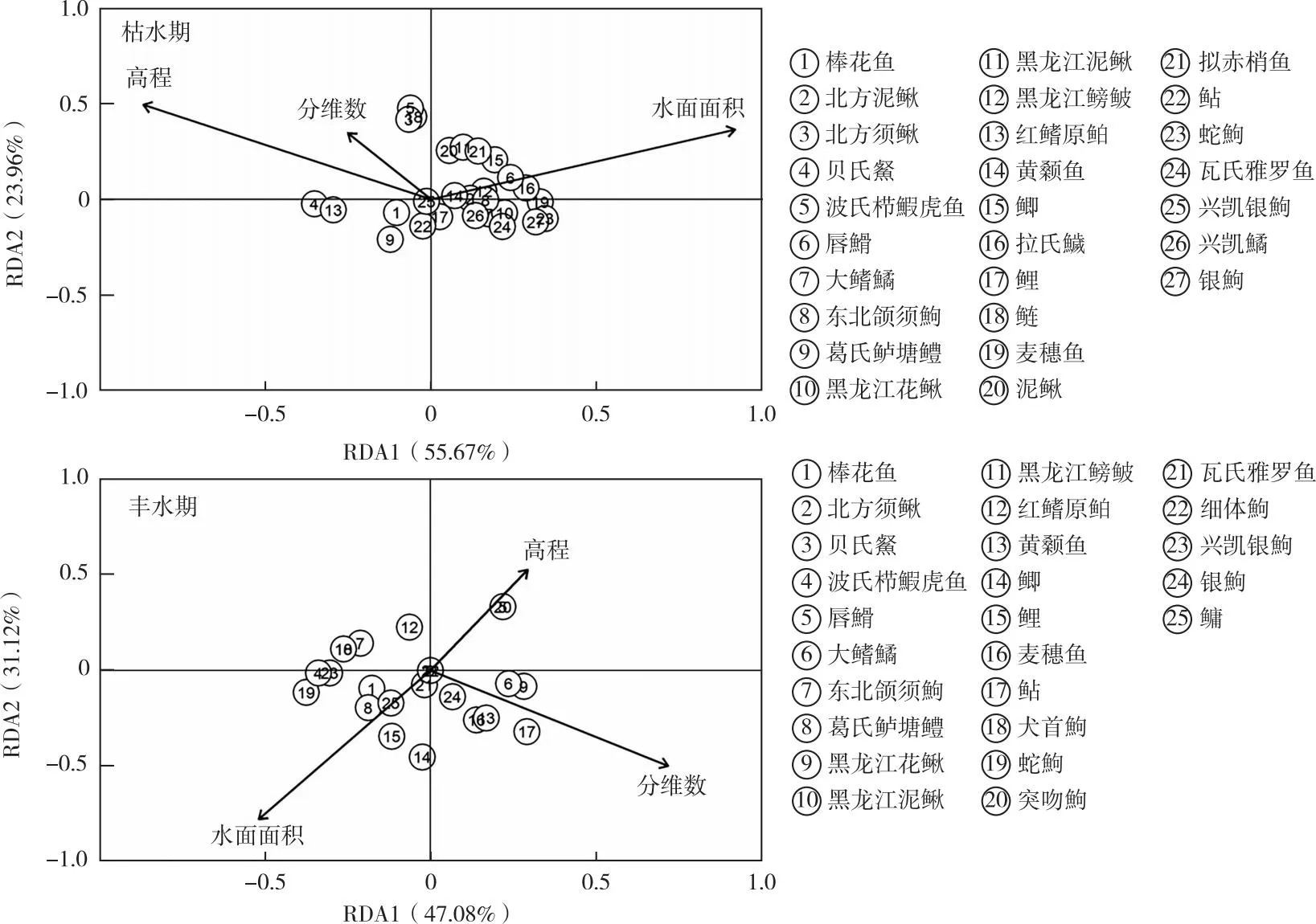
圖9 呼倫湖流域枯水期和豐水期魚類物種與物理生境的RDA分析結果
3 討論
3.1 呼倫湖流域魚類群落組成特征變化趨勢
與歷史資料記錄的呼倫湖魚類物種數量合計33種[45-46]相比,該研究調查結果中呼倫湖流域魚類物種數量有所下降,可能原因是近年來生境條件改變不利于魚類生存繁衍,因此魚類物種數有一定程度降低。與近期魚類物種調查結果相比,2021年該研究共記錄31種魚類,高于2014—2015年調查的21種[31],等于2019年調查的31種[32]。毛志剛等[31]調查的魚類物種數明顯低于2019和2021年調查結果,可能原因是調查區域存在差異,毛志剛等僅在呼倫湖進行魚類資源調查,而霍堂斌等[32]與筆者的魚類調查還涉及呼倫溝、烏爾遜河和克魯倫河等入湖河流以及烏蘭諾爾和貝爾湖等區域。
筆者研究與霍堂斌等[32]調查的魚類物種數存在一定差異,但差異較小。例如,筆者于枯水期在克魯倫河入湖口處采集到2尾擬赤梢魚,但未曾捕獲到黑斑狗魚,而這2種魚類在2019年的魚類調查中均被采集到[32],可能原因是調查點位、網具、采樣方式、采樣時間以及呼倫湖生境條件的變化。另一方面,筆者于豐水期僅在貝爾湖以及靠近貝爾湖出湖口的烏爾遜河采集到5尾黑斑狗魚,而在呼倫湖未捕獲黑斑狗魚,說明呼倫湖仍具備一定大型食魚性土著魚類棲息生境條件,但其生境質量低于貝爾湖。
呼倫湖流域枯水期和豐水期魚類聚類組群空間分布存在一定相似性,同時也表現出一定差異(圖6~7)。因此,季節變化可能是呼倫湖流域魚類群落結構空間分布變化的原因之一。魚類洄游、遷移和繁殖等習性與季節變化密切相關,不同魚類在不同時間可能聚集在不同生境環境中以滿足自身洄游、遷移和繁殖目標。例如,烏爾遜河和克魯倫河等屬于季節性河流,枯水期河水水位變動較大,而枯水期是呼倫湖流域冷水性魚類的繁殖期。枯水期呼倫湖、烏爾遜河和克魯倫河水位降低,水面面積減少,豐水期的黑斑狗魚產卵場面積大幅縮減,導致黑斑狗魚不斷減少[23]。
3.2 呼倫湖流域物理生境對魚類群落結構影響
魚類群落的空間分布特征除了受其自身生活習性影響外,還受環境因子影響,且主導因子空間上存在一定差異。該研究通過對物理生境指標和魚類群落聚類分析,得出呼倫湖流域魚類群落空間分布與物理生境空間分布存在較高相似度,揭示了魚類群落空間分布對物理生境因子的響應特征。呼倫湖湖水來源主要是克魯倫河、烏爾遜河、呼倫溝、海拉爾河等入湖河流匯入以及降水帶來的地表徑流等。由于呼倫湖形狀較曲折,湖水水位變化以及隨之的岸線復雜度和水面面積等變化導致呼倫湖流域物理生境發生改變,水溫、水容量、鹽度、營養鹽和浮游動物等也可能產生變化,進而可能導致魚類群落結構發生變化。呼倫湖東部湖岸地勢平緩,部分淺水區為魚類優良產卵場,烏爾遜河連通了貝爾湖、烏蘭諾爾和呼倫湖,有利于魚類的洄游、遷移和繁殖。烏爾遜河入湖口、烏蘭諾爾、貝爾湖以及烏爾遜河出貝爾湖段具有相近的生境指標,魚類群落結構分組相近。呼倫湖北岸生物多樣性和豐富度均低于其他地區,可能原因是北岸地勢較高,水位較淺,不利于深水魚類生存,由此導致鯉、銀鯽等中下層大型魚類數量較少。此外也有研究發現,不同水體類型的魚類群落結構主要環境因子不同。例如,水溫、水深和鹽度是影響近海和河口等水域魚類群落結構的主要環境因子[47-48],河寬、溫度、海拔、距河口與水壩距離等生境因子則是影響河流魚類群落結構的主要環境因子[49]。
4 結論與建議
目前,呼倫湖流域魚類個體小型化趨勢明顯,魚類多樣性降低,魚類生存環境亟需改進。根據研究結果,可通過提高湖泊和河流岸線復雜度、減少河流的人工截流等,有效提升流域內岸線分維數,在一定程度上提升浮游動植物和水生植物乃至魚類的生存環境。此外,在枯水季等水位較低乃至影響呼倫湖流域魚類繁殖期生態需水要求時,需人工增加流域進水量同時降低蒸發,以滿足魚類產卵需求。
猜你喜歡
中學生數理化·八年級物理人教版(2023年11期)2023-12-26 07:50:02
井岡教育(2022年2期)2022-10-14 03:11:44
中學生數理化(高中版.高考理化)(2022年5期)2022-06-01 06:27:46
中學生數理化(高中版.高考理化)(2022年1期)2022-04-26 13:53:42
中學生數理化(高中版.高考理化)(2022年3期)2022-04-26 13:41:26
中學生數理化(高中版.高考理化)(2022年3期)2022-04-26 13:41:24
課堂內外(初中版)(2022年2期)2022-02-28 02:00:30
云南教育·中學教師(2020年9期)2020-11-16 00:27:58
中學生數理化·八年級物理人教版(2019年9期)2019-11-25 07:33:00
中學生數理化·八年級物理人教版(2017年9期)2017-12-20 08:11:28
