大腸桿菌肌苷酸節點代謝優化高效合成肌苷
2024-03-01 12:56:12劉鐵重高志強侯德欣謝希賢吳鶴云
食品與發酵工業 2024年4期
劉鐵重,高志強,侯德欣,謝希賢,2,吳鶴云,2*
1(天津科技大學 生物工程學院,天津,300457)2(工業發酵微生物教育部重點實驗室(天津科技大學),天津,300457)
肌苷(inosine),又稱次黃苷、次黃嘌呤核苷等,是復合鮮味劑“I+G”的重要原料,廣泛應用于食品生產領域,另外,肌苷作為抗病毒藥物的前體,以及具有鎮靜、擴張血管等作用在醫藥領域也發揮著重要作用[1-2],因此肌苷是一種具有較高經濟和社會價值的生物制品。目前微生物發酵法是生產肌苷的主要方法,但是目前已有肌苷生產菌株的生產效率仍舊較低,且存在營養缺陷、副產物積累和攜帶質粒等弊端,影響發酵法生產肌苷的工業化應用。
肌苷的生物合成是從5-磷酸核糖焦磷酸(phosphoribosyl pyrophosphate,PRPP)和谷氨酰胺(glutamine,Gln)開始,首先經過十步酶促反應生成肌苷酸(inosinic acid,IMP)[3],IMP在5′-核苷酸酶的催化下生成肌苷(圖1)。IMP作為肌苷合成的直接前體物,也是嘌呤生物合成的中心代謝產物,其生物合成受到嚴格的調控,另外,腺苷和鳥苷合成支路與肌苷合成競爭碳流,這是限制肌苷大量合成的關鍵因素。因此,為了構建肌苷生產菌,常規的育種策略包括增強IMP的從頭合成途徑,強化前體物供應,解除IMP合成的反饋調節機制,阻斷降解途徑和競爭途徑等。ASAHARA等[4]通過敲除枯草芽孢桿菌KMBS436中阻遏蛋白編碼基因purR和嘌呤操縱子核糖開關,優化嘌呤操縱子的啟動子序列,解除了嘌呤合成所受的反饋調節機制,并加強了嘌呤的從頭合成途徑;并進一步通過敲除腺苷酸琥珀酸合成酶的基因(purA)和IMP脫氫酶基因(guaB),阻斷肌苷合成的競爭途徑,最終使肌苷產量從1.8 g/L 提升至6.0 g/L。SHIMAOKA 等[5]在大腸桿菌突變菌 I-9 m中過表達了purFK326Q和prsD128A突變基因,解除了限速酶受到的反饋抑制并加強了前體物PRPP的合成,使肌苷產量達到了7.5 g/L。LI等[6]通過敲除枯草芽胞桿菌BS017中的purA基因來阻斷腺苷合成支路,肌苷產量從0.15 g/L提升至7.6 g/L。PEIFER等[7]通過阻斷谷氨酸棒桿菌(ATCC13032)中的腺苷和鳥苷合成支路,使IMP產量增加了45倍。WANG等[8]通過對枯草芽孢桿菌定向進化,降低腺苷酸琥珀酸合成酶的活性,使肌苷產量提高了約4.7倍。以往的研究發現,調整鳥苷和腺苷的合成通量對IMP合成起著非常關鍵的作用,但是直接阻斷鳥苷和腺苷途徑會造成缺陷型,導致菌體生長和肌苷生產受限,雖然補充一定量的鳥嘌呤和腺嘌呤會使生長恢復,但是必須嚴格控制用量,這增加了工藝控制的難度和生產成本。因此如何調控鳥苷和腺苷的合成通量,協調菌體生長和肌苷生產是肌苷生產菌株構建的難點。
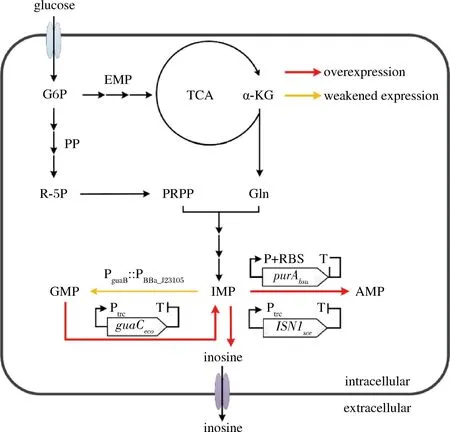
圖1 肌苷合成途徑及本研究的改造策略
本課題在前期的研究工作中,通過一系列代謝工程策略的組合運用,構建了一株肌苷生產菌株[9]。該菌株雖不是缺陷型,但是由于腺苷合成途徑太弱,菌株肌苷生產性能仍受到了一定的限制,為了獲得更高的肌苷生產效率,需要添加一定量的腺嘌呤。本研究在5 L發酵罐上對腺嘌呤的添加量進行了優化,發現在一定范圍內隨著腺嘌呤添加量的提升,肌苷的產量也顯著提升,但是相應副產物的積累也顯著增加。為此,本研究又繼續對肌苷酸節點代謝進行了優化,包括進一步強化腺苷代謝途徑,減弱鳥苷合成支路,以及通過5′-核苷酸酶的篩選和強化以進一步增強肌苷的合成(圖1),降低鳥苷酸等副產物的積累,并避免了腺嘌呤的添加,為肌苷的發酵法工業化生產奠定基礎。
1 材料與方法
1.1 菌株和質粒
菌株和質粒見表1。

表1 菌株和質粒Table 1 Strains and plasmids
1.2 培養基
斜面培養基(g/L):酵母粉5,蛋白胨10,氯化鈉5,牛肉膏10,葡萄糖5,瓊脂20。
種子培養基(g/L):酵母粉5,蛋白胨3,KH2PO4·3H2O 1.3,MgSO4·7H2O 0.5,FeSO4·7H2O 10 mg/L,MnSO4·H2O 10 mg/L。維生素B1、維生素B3、維生素B5、維生素B12和維生素H各2 mg/L。
發酵培養基(g/L):酵母粉4,蛋白胨5,KH2PO4·3H2O 2,MgSO4·7H2O 3,檸檬酸鈉·2H2O 2,FeSO4·7H2O 10 mg/L,MnSO4·H2O 10 mg/L。維生素B1、維生素B3、維生素B5、維生素B12和維生素H各2 mg/L。
1.3 菌株構建及發酵方法
1.3.1 基因編輯方法
本研究采用CRISPR/Cas9技術[10-11]對菌株基因組進行改造,該方法中包括pGRB和pREDCas9兩個質粒。將含有pGRB同源序列和特定sgRNA序列的雙鏈DNA與線性化pGRB在同源重組酶作用下形成gRNA表達質粒;重組片段則通過重疊PCR制備。首先將pREDCas9質粒導入大腸桿菌感受態細胞中,再將pGRB質粒和重組片段導入到含有pREDCas9質粒的感受態細胞,然后篩選陽性重組子,最后依次消除pGRB和pREDCas9質粒,完成菌株構建。
1.3.2 搖瓶發酵
將菌株接種至斜面培養基中,37 ℃培養12 h后,將菌體接種至發酵培養基中,37 ℃、200 r/min培養28 h,發酵培養基中添加苯酚紅作為指示劑,根據培養基顏色變化手動補加氨水以維持培養基pH值為7.0左右。
1.3.3 分批補料發酵
菌株在裝有種子培養基的5 L發酵罐中進行菌體活化,發酵過程pH為7.0左右,溫度37 ℃,溶氧保持在15%~25%,當OD600達到10~16時,按15%的接種量接入5 L發酵罐中進行發酵培養,發酵過程維持pH值為7.0左右,溫度35 ℃,溶氧控制在20%~30%。發酵過程中流加質量濃度為800 g/L的葡萄糖溶液,維持罐中葡萄糖質量濃度在0.1~5 g/L。
1.4 基因轉錄水平測定
使用試劑盒Easep?Super RNA kit(Promega,上海)提取總RNA,然后使用試劑盒PrimeScriptTM RT Master Mix (TaKaRa, 大連)進行反轉錄,相關操作參照說明書。通過RT-PCR測定基因相對轉錄量,采用嵌合熒光法(TB Green?Premix ExTaqTM Ⅱ,Takara),16S rRNA作為內標,RT-PCR結果用2-ΔΔCt法進行分析。
1.5 檢測方法
菌體生物量以發酵液在600 nm處的吸光度(OD600)表征。
肌苷及其他嘌呤類副產物濃度采用HPLC方法檢測:所用儀器為Thermo U3000;色譜柱為phenomenex Gemini 5u C18 110A(150 mm×4.6 mm);流動相為2%乙腈和0.05%三氟乙酸的混合水溶液;柱溫30 ℃;流速 1 mL/min;檢測波長260 nm。
1.6 數據統計分析
發酵數據代表3組平行發酵數據的平均值和標準差。利用t檢驗雙尾分布對兩組發酵參數進行單向方差分析。“*”表示P<0.05,表示結果差異顯著;“**”表示P<0.01,表示結果差異極其顯著。
2 結果與分析
2.1 腺嘌呤添加對INO4-5發酵的影響
INO4-5是實驗室前期構建的一株肌苷生產菌,分別對嘌呤從頭合成途徑、肌苷的降解途徑和肌苷轉運系統進行了改造。特別是弱化了腺苷酸琥珀酸合成酶(PurA)的活性,在不造成缺陷型的前提下實現了肌苷的積累,但是由于PurA的活性過低,生長還是受到了明顯抑制,添加一定量的腺嘌呤可恢復菌體生長并促進肌苷的生產。本研究在5 L發酵罐上對腺嘌呤的添加量進行了優化。結果如圖2所示,在不添加腺嘌呤時,菌體生長和肌苷生產性能都比較低,而隨著腺嘌呤添加量的增加,菌體的生長和肌苷的產量在也逐漸增加。當腺嘌呤添加量為3 g/L時,肌苷的最大產量可以達到30.6 g/L,OD600值可達85.8,繼續增加腺嘌呤的添加量雖然使得最大OD600值提高至92.4,但對肌苷生產影響不大。結果表明INO4-5中PurA的活性過低,雖然實現了不用額外添加腺嘌呤也能積累一定量的肌苷,但是由于菌體生長受到明顯抑制,需要補充定量的腺嘌呤才能獲得較高的肌苷產量。另外對不同腺嘌呤的添加量下,INO4-5發酵液中的副產物進行了檢測(圖2-C),發現有腺苷酸,鳥嘌呤和鳥苷酸等副產物的積累,并且這些副產物的濃度也會隨著腺嘌呤添加量的增加而增加。腺嘌呤的添加會增加發酵工藝的復雜程度,提高發酵成本;鳥苷酸等副產物的積累會抑制嘌呤操縱子的表達[8],不利于肌苷的合成,降低碳源的利用率,同時增加了下游的提取難度。為解決這兩個問題則需進一步優化IMP節點的代謝流。
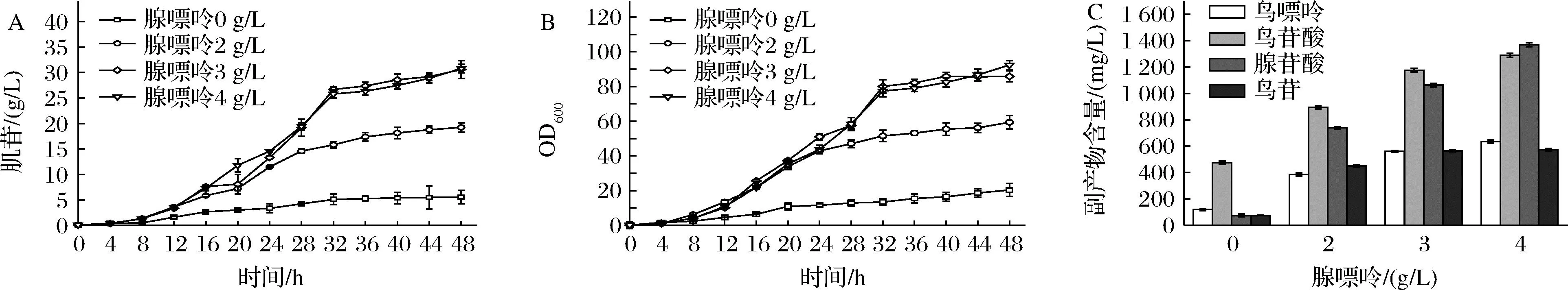
A-肌苷產量曲線;B-菌體生長曲線;C-副產物濃度圖2 腺嘌呤添加量對肌苷發酵的影響
2.2 優化purA基因的轉錄表達對肌苷發酵的影響
肌苷合成的直接前體是肌苷酸,但是肌苷酸除了在5′-核苷酸酶的作用生成肌苷外,還會在腺苷酸琥珀酸合成酶(purA基因編碼)的作用下生成腺苷酸琥珀酸,從而流向腺苷的合成,也會在IMP脫氫酶(guaB編碼)的作用下生成黃苷酸(xanthylic acid,XMP),進而流向鳥苷合成途徑。因此肌苷的過量生產需要適當弱化腺苷和鳥苷合成支路碳流來實現[12],使代謝通量轉向肌苷生產。INO4-5中腺苷合成支路的PurA活性太弱,導致菌體內的ATP供應不足,抑制了菌體的生長并限制了肌苷生產,因此在發酵過程中仍需要添加腺嘌呤才能獲得較好的發酵結果,這無疑增加了肌苷發酵生產的成本。因此為了避免腺嘌呤的添加,同時滿足菌體生長和肌苷生產的需要,需要對PurA的活性進行適當的強化。為此,本研究引入了來源于枯草芽孢桿菌的purAbsu基因,并分別使用了5個表達強度依次增加的啟動子(BBa_J23114、BBa_J23105、BBa_J23110、BBa_J23106和BBa_J23101,啟動子來源于https://parts.igem.org/Promoters/Catalog/Anderson)來啟動purAbsu基因的表達,構建了菌株INO5-1~5。將INO4-5和INO5-1~5六株菌進行搖瓶發酵,結果如圖3-A所示。結果顯示,強化purAbsu基因后,菌體的生長得以恢復,并且隨著啟動子強度的增強,菌體的生長也越來越好。但是purAbsu的表達過強,會導致代謝流過多流向腺苷支路,從而減少肌苷的生產,其中,INO5-2的肌苷產量最高,達到6.4 g/L,最大OD600值可恢復至37。另外,基因的表達也跟核糖體結合位點(ribosomebinding site,RBS)有關,于是本研究對INO5-2中purAbsu基因的RBS進行了替換,繼續微調purAbsu基因的表達強度。選用了RBS03、RBS09和RBS10[13]3個強度依次增強的RBS,分別構建了菌株INO5-2a~c。將INO5-2與INO5-2a~c同時進行搖瓶發酵實驗,結果如圖3-B所示,RBS的替換對菌體生長的影響不顯著,而INO5-2b的肌苷產量達到了6.9 g/L,相比對照菌INO5-2提高了7.5%。
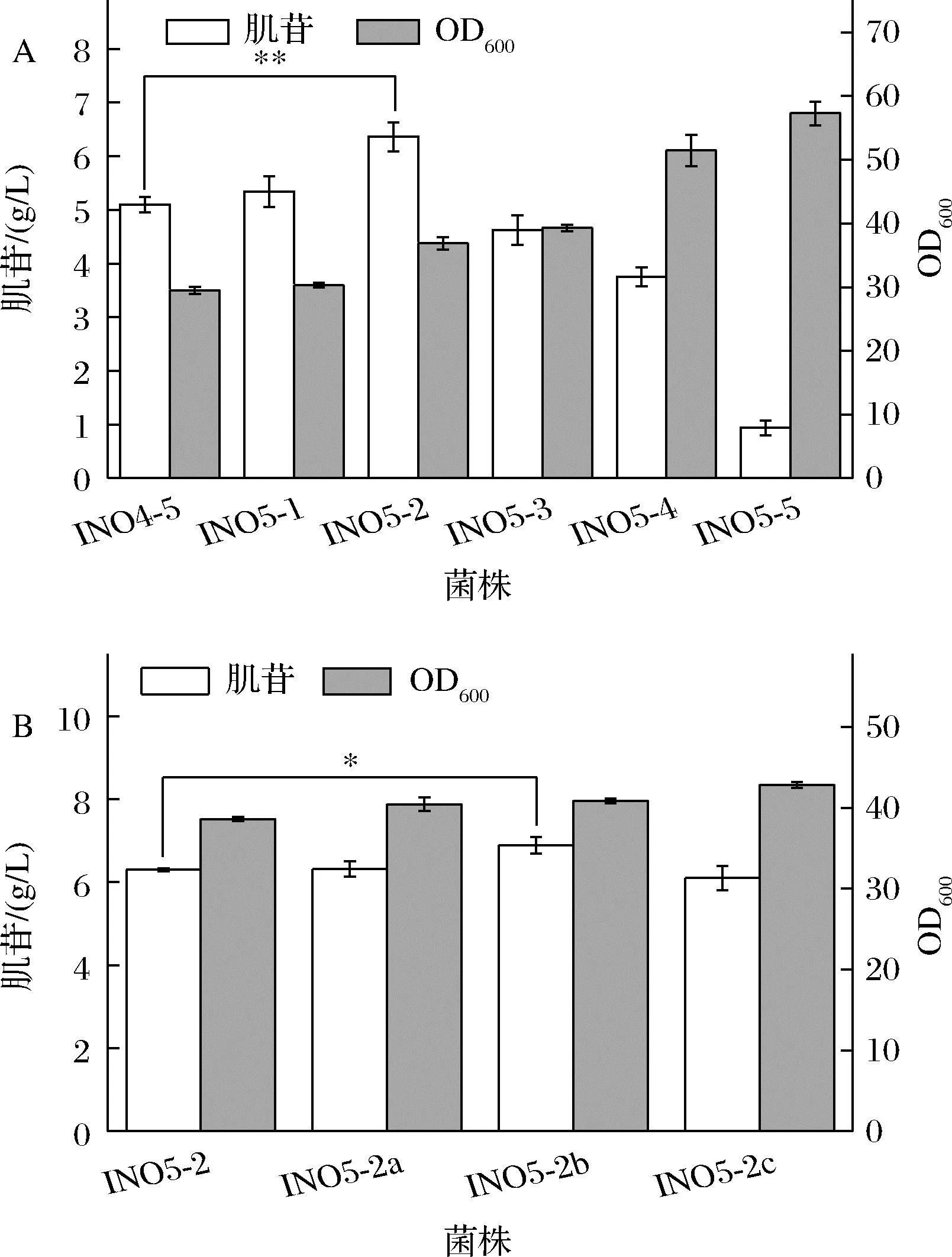
A-不同強度啟動子啟動purAbsu對肌苷發酵的影響;B-替換purAbsu的RBS對肌苷發酵的影響圖3 改造腺苷合成支路對肌苷發酵的影響
將本研究中搖瓶肌苷產量最高的INO5-2b在5 L發酵罐中進行分批補料發酵,結果如圖4-A所示,不用額外添加腺嘌呤,INO5-2b發酵48 h可以生產肌苷29.5 g/L,與INO4-5的發酵產量(添加3 g/L腺嘌呤)相當。結果說明通過合理調整purAbsu的表達強度可以很好地平衡菌體生長與肌苷生產的平衡,在不造成缺陷型的前提下實現肌苷的高效生產。但經檢測發現,發酵結束時,發酵液中仍舊積累了鳥苷酸、鳥苷和鳥嘌呤等副產物(圖4-B),未檢測到腺苷酸,INO4-5在添加腺嘌呤的發酵過程中會積累腺苷酸可能是腺嘌呤通過補救合成途徑生成的。因此,為了降低甚至阻斷這些副產物的合成,本研究接著對INO5-2b的鳥苷合成支路進行了改造。
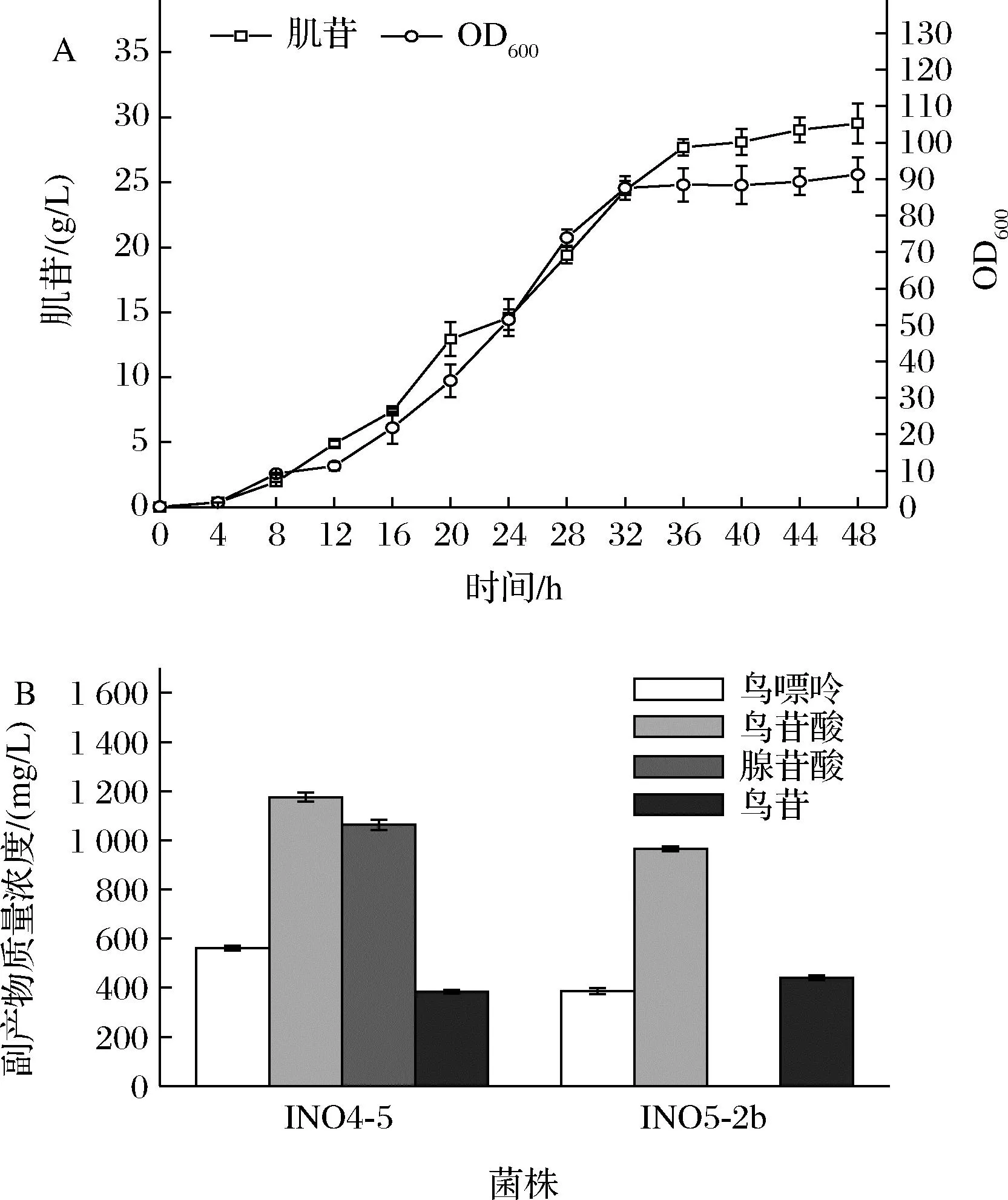
A-發酵生產曲線;B-副產物濃度圖4 菌株INO5-2b的5 L發酵罐發酵結果
2.3 鳥苷合成途徑改造對肌苷發酵的影響
INO5-2b在發酵過程中,鳥苷途徑的一些副產物如鳥苷酸、鳥嘌呤和鳥苷均有不同程度積累。有報道指出,上述副產物在胞內積累會損害細胞活力,使菌體快速老化[14],還會抑制PRPP合成酶(Prs)的活性[7]以及嘌呤操縱子基因的轉錄表達,不利于肌苷的生產。為了驗證這些副產物積累對肌苷生產的影響,本研究將INO5-2b分別在添加不同濃度鳥苷酸、鳥苷或鳥嘌呤的條件下進行發酵實驗,分別選擇了0、0.2、0.4和0.8 g/L 4個梯度(圖5)。結果顯示,隨著鳥苷酸和鳥嘌呤的濃度增加,其對肌苷產量和菌體生長的抑制作用也愈發顯著,而在實驗范圍內,鳥苷的添加未對肌苷產量和菌體生長產生顯著影響。
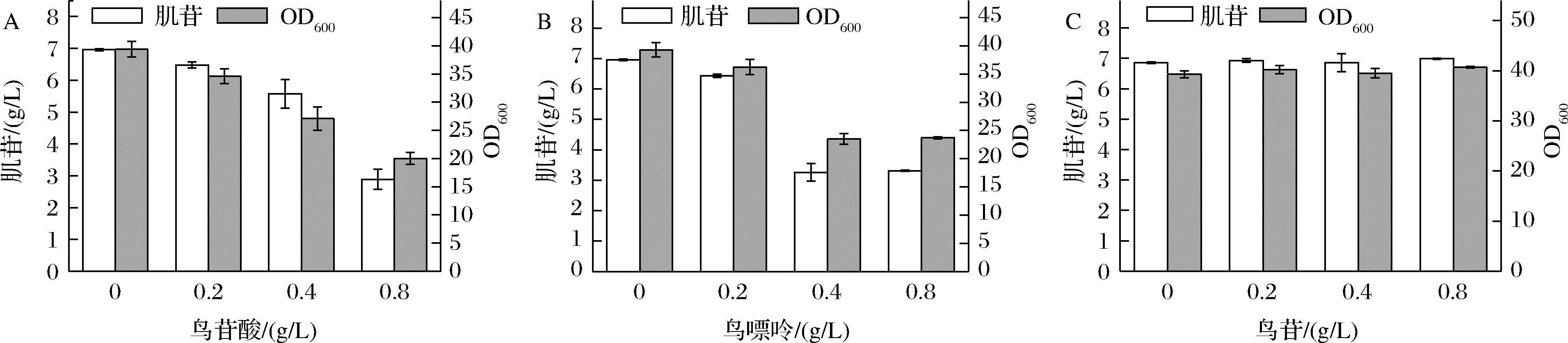
A-鳥苷酸對肌苷發酵的影響;B-鳥嘌呤對肌苷發酵的影響;C-鳥苷對肌苷發酵的影響圖5 鳥苷支路副產物對肌苷發酵的影響
為了減少鳥苷支路副產物的積累,本研究首先將guaB基因自身的啟動子進行替換,以期降低guaB基因的表達,減弱鳥苷合成支路。仍舊選用了強度依次增強的啟動子BBa-J23114、BBa-J23105、BBa-J23110、BBa-J23106和BBa-J23101,分別替換了INO5-2b的guaB自身啟動子,構建了菌株INO6-1a~e, 將INO5-2b和INO6-1a~e六株菌進行搖瓶發酵驗證,結果顯示(圖6-A),與對照菌株INO5-2b相比,INO6-1a的OD600由39.5降低到了23.7,降低了40%,肌苷產量為5.9 g/L;INO6-1b的OD600相比對照菌株基本無變化,而肌苷產量由6.9 g/L提升至7.5 g/L,提高了8.7%;INO6-1c和INO6-1d同出發菌株相比,菌體生長和肌苷產量都未發生明顯變化;INO6-e相比對照菌株,菌體生長無明顯差異,但是肌苷產量下降至5.5 g/L,降低了20%。對發酵不同階段guaB基因的轉錄量進行檢測,結果顯示(圖6-B)發酵6 h和12 h時,相比于對照菌株,INO6-1a和INO6-1b的guaB基因的轉錄量分別降低了40.1%和15.2%,INO6-1c和INO6-1 d的guaB基因的轉錄量與對照菌相當,而INO6-1e的guaB基因轉錄量增加了65%。從副產物的檢測情況來看(表2),發酵結束時,INO6-1a的發酵基本檢測不到鳥苷酸和鳥嘌呤,INO6-1b中鳥苷酸、鳥嘌呤和鳥苷的質量濃度分別為0.051、0.126和0.167 g/L,較對照菌分別降低了76.4%、52.1%和18.1%;INO6-1c和INO6-1 d中3種副產物的濃度和對照菌株相當;而INO6-1e的鳥苷酸、鳥苷和鳥嘌呤的濃度分別較出發菌株提高了11.1%、4.6%和19.6%。結果說明通過啟動子替換,降低guaB基因的轉錄量,從而減弱鳥苷合成途徑,不僅有助于肌苷產量的提升,還有效降低了鳥苷酸等副產物的積累,但是guaB基因的轉錄強度太弱,則會抑制菌體的生長。綜合比較來看,用啟動子BBa_J23105替換guaB基因原有啟動子的效果最佳,在保證菌體生長的前體下,有效提升了肌苷產量并減低了副產物的積累。

表2 不同工程菌株搖瓶發酵副產物積累情況Table 2 Byproducts accumulation of different engineered strains in shake flasks
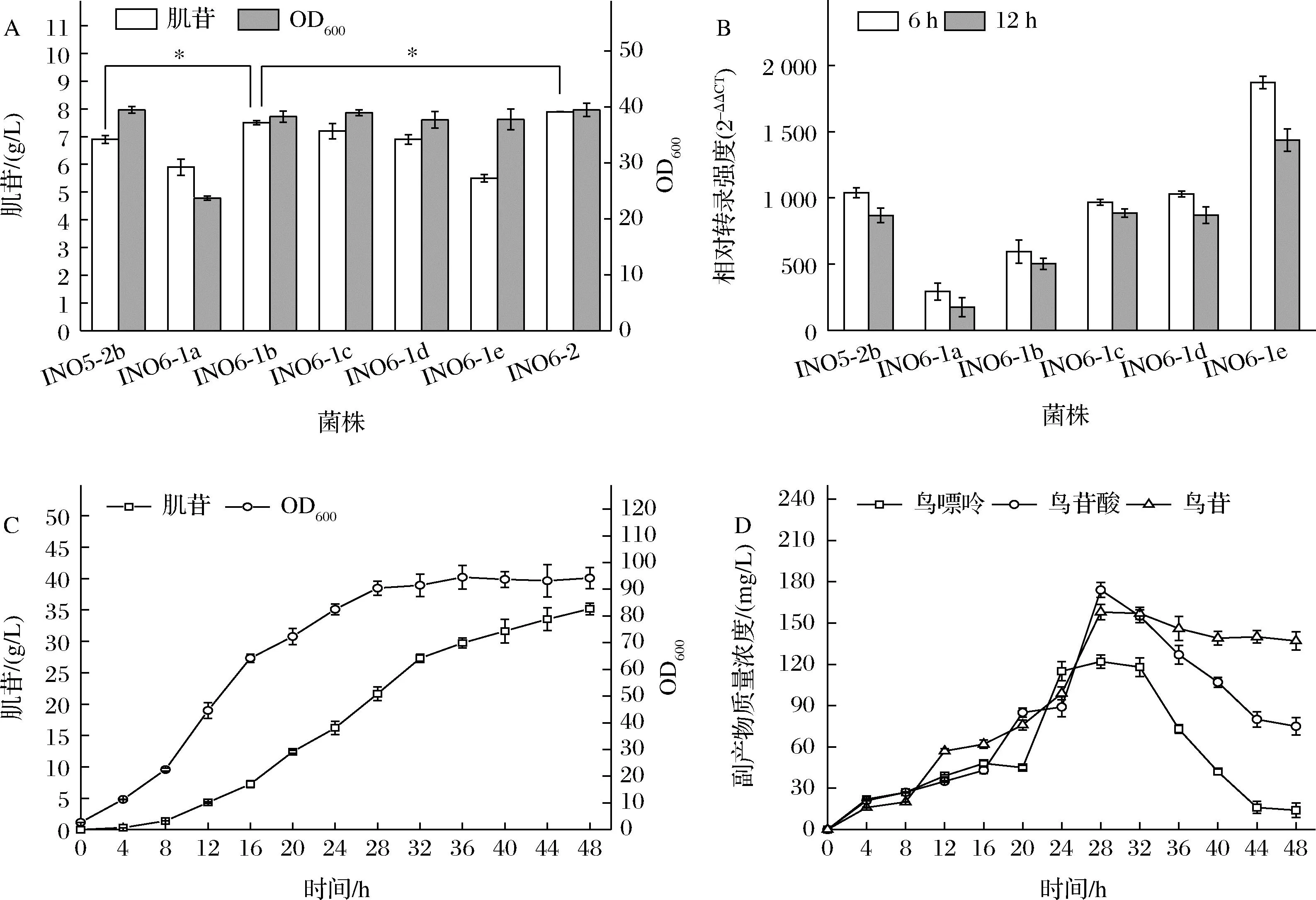
A-搖瓶發酵結果;B-guaB基因轉錄水平;C-INO6-2的5 L發酵罐發酵結果;D-INO6-2發酵過程中副產物濃度圖6 鳥苷支路改造對肌苷發酵的影響
另外,GMP還原酶(guaC基因編碼)可催化鳥苷酸生成肌苷酸,理論上有助于降低鳥苷合成支路副產物并促進肌苷積累。于是本研究在INO6-1b中對guaC基因進行了過表達,構建了菌株INO6-2。搖瓶結果顯示(圖6-A),相比INO6-1b,INO6-2的肌苷產量由7.5 g/L提升至7.9 g/L,提高了5.3%,副產物檢測結果顯示,發酵液中幾乎檢測不到鳥苷酸和鳥苷,只有少量的鳥嘌呤存在(表2)。將INO6-2在5 L發酵罐進行發酵實驗(圖6-C),結果顯示INO6-2經5 L發酵罐測試,OD600達到了94.2,肌苷的產量達到了35.2 g/L,比INO5-2b提升了19.3%,發酵過程中鳥苷支路副產物均處于較低的水平(圖6-D)。
2.4 強化5′-核苷酸酶對肌苷發酵的影響
5′-核苷酸酶作為菌體內參與合成肌苷的最后一個酶,水解5′-核苷酸生成核苷和無機磷酸鹽。但是大多數的5′-核苷酸酶的底物特異性差,可作用于各種嘌呤或嘧啶核苷酸等。因此篩選活性更高、特異性更強的5′-核苷酸酶將有利于肌苷的生產。大腸桿菌中有多種5′-核苷酸酶[15-16],分別由yrfG、ushA、umpH和umpG等基因編碼;目前常用于發酵生產肌苷的菌株枯草芽孢桿菌5′-核苷酸酶基因為yfkN[17];在釀酒酵母中發現了催化肌苷酸生成肌苷的特異性5′-核苷酸酶基因ISN1[18]。本研究將上述具有催化肌苷酸生成肌苷的核苷酸酶基因yrfG、ushA、umpH、umpG、yfkN和ISN1,分別整合在了INO6-2基因組的ycgH假基因位點上,并由Ptrc啟動子啟動表達,構建了菌株INO7-1~6。搖瓶發酵結果如圖7-A所示。
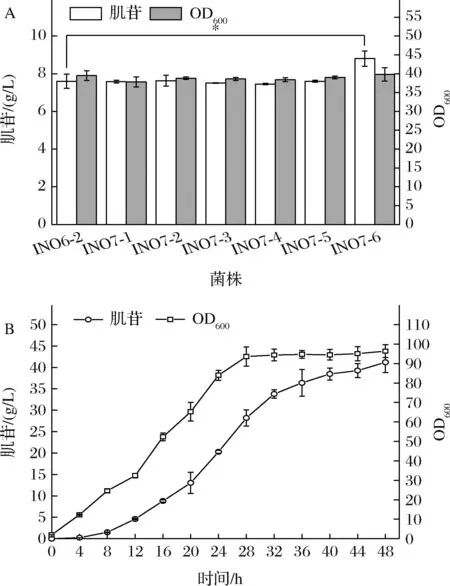
A-INO6-2和INO7-1~6的搖瓶發酵結果;B-工程菌INO7-6的5 L發酵罐發酵結果圖7 強化5′-核苷酸酶對肌苷發酵的影響
結果顯示,與對照菌INO6-2相比,INO7-1~5這五株菌的生長和肌苷生產基本無差別,說明來源于大腸桿菌和枯草芽孢桿菌的5′-核苷酸酶基因的過表達對菌體生長和肌苷生產量沒有影響。而INO7-6的菌體生長相比對照菌變化不大,但是肌苷產量由7.6 g/L提升至8.8 g/L,相比對照菌株提升了15.8%。說明來源于釀酒酵母的ISN1基因的表達有效促進了肌苷的生產。為測試菌株 INO7-6的放大生產性能,在5 L發酵罐中進行了分批補料發酵,發酵結果如圖7-B所示,菌株在發酵初期生長緩慢,28 h OD600逐漸達到穩定,并進入穩定期。0~12 h,肌苷合成速率較慢;12~32 h,肌苷保持了較高的合成速率,32 h之后肌苷合成速率逐漸減慢。48 h的肌苷的最終產量為41.2 g/L,葡萄糖產率為0.17 g/g葡萄糖,生產強度為0.86 g/(L·h)。
3 結論與討論
為了解決肌苷生產菌INO4-5在發酵過程中副產物高、需外源添加腺嘌呤等問題,并進一步提高其肌苷的發酵效率,本研究對其肌苷酸節點代謝流進行了優化。首先為了強化腺苷合成途徑,表達了來源于枯草芽孢桿菌的腺苷酸琥珀酸合成酶編碼基因purAbsu,并使用不同強度的啟動子和核糖體結合位點調整其轉錄表達的強度,所得最優菌株在不添加腺嘌呤的前提下,5 L罐上的肌苷發酵產量達到了29.5 g/L;鳥嘌呤和鳥苷酸等副產物的積累會抑制菌體的生長和肌苷生產,為了降低這些副產物的積累,本研究對鳥苷合成支路進行了改造,通過啟動子的替換,弱化了IMP脫氫酶編碼基因guaB的轉錄表達,并過表達guaC基因,促使鳥苷酸回流至肌苷酸,使鳥苷酸等副產物的含量顯著降低,并有效提高了肌苷產量。最后對5′-核苷酸酶進行了篩選和強化,發現引入來自釀酒酵母的特異性5′-核苷酸酶基因(ISN1),可有效促進肌苷生產。最終所得菌株INO7-6,搖瓶發酵28 h肌苷產量為8.8 g/L;在5 L發酵罐上進行分批補料發酵48 h,肌苷產量達到41.2 g/L,生產強度為0.86 g/(L·h),葡萄糖的產率為0.17 g/g葡萄糖。菌株INO7-6不含質粒,且在發酵過程不用額外添加腺嘌呤或鳥嘌呤等營養物,可簡化發酵工藝并降低生產成本,具有良好的工業化應用前景。為進一步提高肌苷發酵生產效率,下一步的研究可以針對中心代謝進行改造,比如重新分配糖酵解與磷酸戊糖途徑的碳流,增強前體物PRPP的供應水平,促進葡萄糖向肌苷的轉化,提高轉化率,以滿足肌苷工業化生產的需求。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
小學科學(學生版)(2020年10期)2020-10-28 07:52:12
中國化肥信息(2020年7期)2020-03-19 01:54:02
中國果業信息(2019年10期)2019-11-13 01:21:34
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國軍轉民(2017年6期)2018-01-31 02:22:28
汽車零部件(2014年11期)2014-09-18 11:57:16
