兩種重要經(jīng)濟石斑魚腸道微生物群落結構及代謝功能的比較分析*
2024-02-24 08:45:26杜倩平謝廣婷鐘仲軒張明清吳錦輝
海洋與湖沼 2024年1期
杜倩平 謝廣婷 鐘仲軒 張明清, 吳錦輝 舒 琥①
(1. 廣州大學生命科學學院 廣東廣州 510006; 2. 中山大學生命科學學院 廣東廣州 510275; 3. 廣東省農(nóng)業(yè)技術推廣中心 廣東惠州 516081)
腸道微生物群落可以通過發(fā)酵宿主難以消化的食物成分、代謝多種化合物參與宿主的生長、代謝以及免疫反應等生理過程, 并能與中樞神經(jīng)系統(tǒng)相互作用, 影響大腦功能, 是宿主體內的重要組成部分(Flintetal, 2012; Clarkeetal, 2014; Liuetal, 2023)。Lee 等(2022)研究發(fā)現(xiàn), 提高腸道微生物來源的短鏈脂肪酸能夠緩解老年腦卒小鼠的神經(jīng)缺損與炎癥。Duca 等(2012)研究結果表明, 缺乏腸道微生物的無菌小鼠其瘦素和生長素釋放肽水平明顯降低, 即使攝入更多的熱量, 仍然比對照組小鼠更瘦。在魚類中, 腸道微生物同樣與營養(yǎng)物質的消化、吸收以及免疫反應息息相關。Abid 等(2013)揭示了腸道微生物對大西洋鮭(Salmosalar)腸黏膜細胞的保護作用: 腸道微生物可以改善腸道黏膜細胞的形態(tài)并刺激先天性免疫反應。Li等(2023)證明, 宿主遺傳學與腸道微生物群落的相互作用能夠改善雜交魚(Megalobramaamblycephala♀ ×carnivorousCulteralburnus♂)的攝食行為, 厚壁菌門(Firmicutes)、放線菌門(Actinobacteria)和綠菌門(Chloroflexi)的豐度與宿主生長水平呈正相關。因此,對魚類腸道微生物進行表征, 在解析其性狀方面具有重要的意義。
與淡水養(yǎng)殖相比, 海水養(yǎng)殖不受土地稀缺和水資源供應不足的影響, 其在滿足日益增長的食物需求方面具有巨大的潛力(Wuetal, 2021a)。自2000 年起, 中國海水養(yǎng)殖產(chǎn)量穩(wěn)步增長, 產(chǎn)值逐年提高:2020 年中國海水養(yǎng)殖產(chǎn)量達到2 135.31 萬t, 產(chǎn)值達到 3 836.20 億元(農(nóng)業(yè)農(nóng)村部漁業(yè)漁政管理局等,2021)。石斑魚因其營養(yǎng)價值高、肉質細嫩、味道鮮美等優(yōu)點深受消費者喜愛, 是重要的海水養(yǎng)殖魚種。2020 年, 中國石斑魚產(chǎn)量為19.20 萬t, 相較2016 年增長77.3%, 其中虎龍雜交斑(Epinephelusfuscoguttatus♀ ×E.lanceolatus♂)占市場份額的70%以上(Yanget al, 2021)。虎龍雜交斑繼承了其親本生長速度快、抗逆性能強的優(yōu)點, 并且口感極佳、經(jīng)濟效益高, 能夠承受較高的養(yǎng)殖密度, 是水產(chǎn)養(yǎng)殖戶和消費者的首選種類(Sunetal, 2016; Tanetal, 2019; Amoahetal,2022)。此外, 斜帶石斑魚(E.coioides)亦是東南亞國家常見的經(jīng)濟養(yǎng)殖石斑魚品種, 同樣具有營養(yǎng)豐富、味道鮮美、易于飼養(yǎng)的特點(Baietal, 2021; Xiao Joeetal, 2021; Laietal, 2022)。
目前, 虎龍雜交斑和斜帶石斑魚作為具有重要經(jīng)濟價值的石斑魚養(yǎng)殖品種, 相關研究主要集中在其營養(yǎng)、攝食、生長、免疫等方面, 關于其腸道微生物群落的結構與功能的比較研究暫未見報道。因此,本研究利用16S rDNA 高通量測序技術, 對虎龍雜交斑和斜帶石斑魚腸道微生物群落進行解析, 探究兩種石斑魚腸道微生物組成與功能上的差異。本研究將為石斑魚腸道微生物與性狀關聯(lián)研究提供理論基礎,為選擇合適的石斑魚養(yǎng)殖品種提供參考, 進而提高石斑魚養(yǎng)殖業(yè)的經(jīng)濟效益。
1 材料與方法
1.1 樣品采集
虎龍雜交斑[(47.47±5.92) g]和斜帶石斑魚[(43.26±2.33) g]購自廣東省海洋漁業(yè)試驗中心。在解剖前, 使用250 mg/L 丁香酚(生工生物, 中國)將魚麻醉。使用75%乙醇對魚表面消毒, 用無菌解剖工具無菌解剖,打開腹腔, 取魚的全腸, 去除表面附著物。隨后, 將腸道置于無菌2 mL 離心管中, 立即用液氮冷凍并保存于-80 °C, 直至提取DNA。每種魚各5 個重復。
1.2 16S rDNA 的提取及測序
使用細菌基因組DNA 提取試劑盒(諾唯贊, 中國)提取細菌基因組DNA, 利用Thermo NanoDrop One(Thermo Fisher Scientifif, USA)檢測DNA 的純度與濃度。
以基因組DNA 為模板, 使用帶標簽序列的特異引物(515 F: GTGCCAGCMGCCGCGGTAA 和806R:GGAC-TACHVGGGTWTCTAAT)及 TaKaRa Premix Taq?Version 2.0 (TaKaRa Biotechnology Co., China)通過PCR 擴增16S rDNA V4 區(qū)。
PCR 反應條件如下: 94 °C 5 min; 94 °C 30 s,52 °C 30 s, 72 °C 30 s, 30 個循環(huán); 72 °C 10 min。按照NEBNext? Ultra? II DNA Library Prep Kit for Illumina?(New England Biolabs, USA)標準流程進行建庫操作, 使用Illumina Nova 6000 平臺對構建的擴增子文庫進行PE250 測序(廣東美格基因科技有限公司, 中國)。
1.3 生物信息學分析
使用fastp (https://github.com/OpenGene/fastp)和cutadapt (https://github.com/marcelm/cutadapt/) 處理Paired-end Raw Reads 得到質控后的paired-end Clean Reads。利用 usearch-fastq_mergepairs (http://www.drive5.com/usearch/)拼接paired-end Clean Reads 以獲得Raw Tags。利用fastp (https://github.com/OpenGene/fastp)對Raw Tags 進行質量過濾得到Clean Tags。
使用UPARSE 將相似度≥97%的Clean Tags 分配到相同的操作分類單元(OTU) (Edgar, 2013)。隨后,利用usearch-sintax 將每個OTU 的代表序列與SILVA數(shù)據(jù)庫進行比對獲得物種注釋信息。利用Mothur 生成稀釋曲線并計算Alpha 多樣性指數(shù), 包括Chao1 指數(shù)、Robbins 指數(shù)、Shannon 指數(shù)和Simpson 指數(shù)。采用Kruskal-Wilcox 檢驗計算各組間Alpha 多樣性指數(shù)的顯著性差異。Beta 多樣性使用非度量多維尺度分析(NMDS)進行可視化(Cuietal, 2018)。采用線性判別分析(LDA)差異大小(LEfSe)法(http://huttenhower.sph.harvard.edu/lefse)對虎龍雜交斑和斜帶石斑魚腸道微生物進行鑒定, 以揭示標志微生物。LEfSe 方法使用Kruskal-Wallis 秩和檢驗檢測各組的顯著性差異豐度特征, 并使用歸一化的相對豐度矩陣進行LDA以估計每個特征。餅圖及百分比堆積圖使用Origin 2018 (OriginLab, USA)進行繪制。
1.4 網(wǎng)絡分析及功能預測
采用網(wǎng)絡分析評價腸道中微生物群落之間的相互作用(Zhouetal, 2021)。基于細菌類群間的Spearman 相關系數(shù), 建立了微生物群落的相關性網(wǎng)絡, 構建網(wǎng)絡的標準為r>0.6 和p<0.05。使用Gephi 軟件進行網(wǎng)絡可視化。
采用基于未觀察狀態(tài)重建的群落系統(tǒng)發(fā)育研究(PICRUSt)預測微生物群落的功能。對16S rRNA 基因拷貝數(shù)進行歸一化后, 參考京都基因和基因組百科全書(KEGG)同源性(KO)數(shù)據(jù)庫預測微生物區(qū)系功能(Maetal, 2019)。使用主成分分析(PCA)對腸道微生物區(qū)系功能多樣性進行可視化。
2 結果
2.1 16S rDNA 測序數(shù)據(jù)及Alpha 多樣性分析
通過將測序所獲得的序列進行篩選和聚類, 共獲得1 952 個操作分類單元(OTUs)。虎龍雜交斑樣品含有1 568 個OTUs, 斜帶石斑魚樣品含有1 283 個OTUs。二者有共同899 個OTUs, 虎龍雜交斑特有669個OTUs, 斜帶石斑魚特有384 個OTUs (圖1a)。稀釋曲線顯示, 各曲線隨著序列數(shù)的增加而趨于平緩,說明測序深度足以提供樣品中所有微生物的信息(圖1b)。Alpha 多樣性結果顯示, 虎龍雜交斑腸道微生物的Chao1 指數(shù)高于斜帶石斑魚, 但無顯著性差異(P>0.05),Simpson 指數(shù)則顯著高于斜帶石斑魚(P<0.05); 斜帶石斑魚腸道微生物的Robbins 指數(shù)、Shannon 指數(shù)高于虎龍雜交斑, 差異不顯著(P>0.05, 表1)。
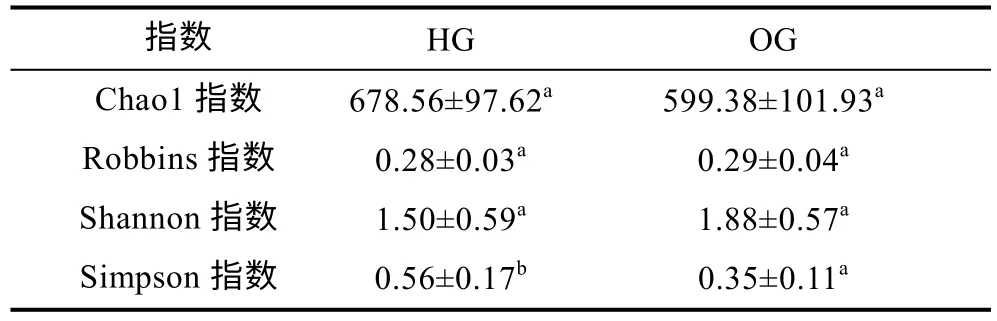
表1 虎龍雜交斑和斜帶石斑魚腸道微生物群落的Alpha多樣性指數(shù)Tab.1 The Alpha diversity of gut microbes from hybrid grouper and orange-spotted grouper
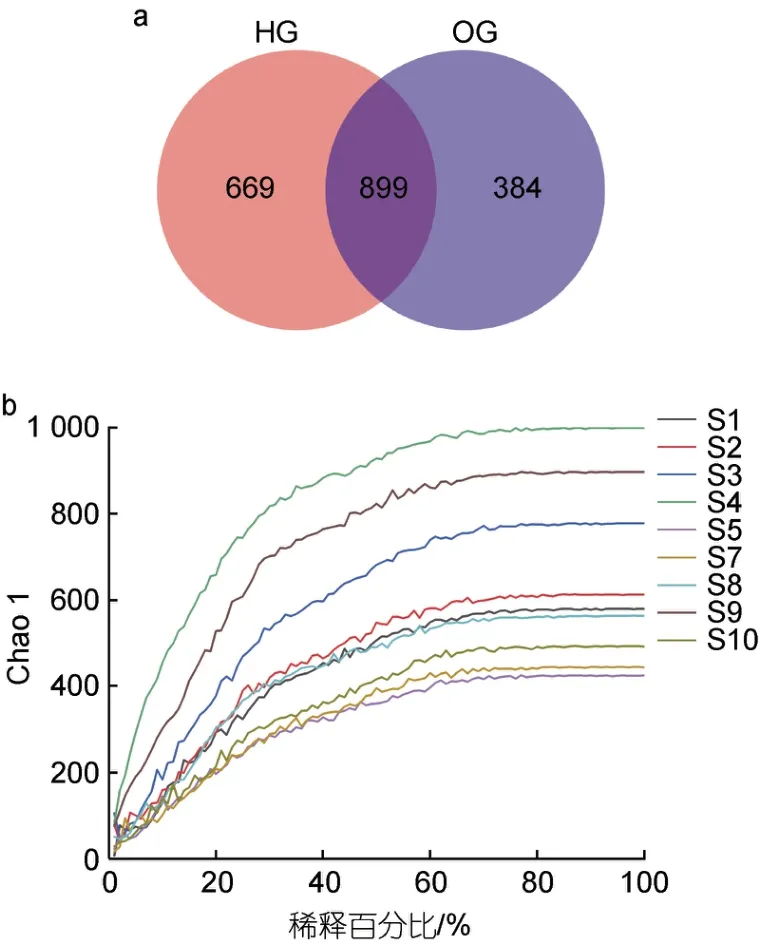
圖1 虎龍雜交斑和斜帶石斑魚腸道微生物OTU 數(shù)目和稀釋曲線Fig.1 OTU number and dilution curve of gut microbes of hybrid grouper and orange-spotted grouper
2.2 腸道微生物群落組成
在兩種石斑魚腸道中共鑒定出7 個主要的門類(相對豐度>0.1%)和21 個屬類(相對豐度>0.1%), 優(yōu)勢門類為變形菌門(Proteobacteria, 68.14%)、厚壁菌門(19.18%)和梭桿菌門(Fusobacteria, 10.00%) (圖2); 優(yōu)勢屬類為弧菌屬(Vibrio, 45.26%)、羅姆布茨菌屬(Romboutsia,13.50%)和鯨桿菌屬(Cetobacterium, 9.99%) (圖3)。
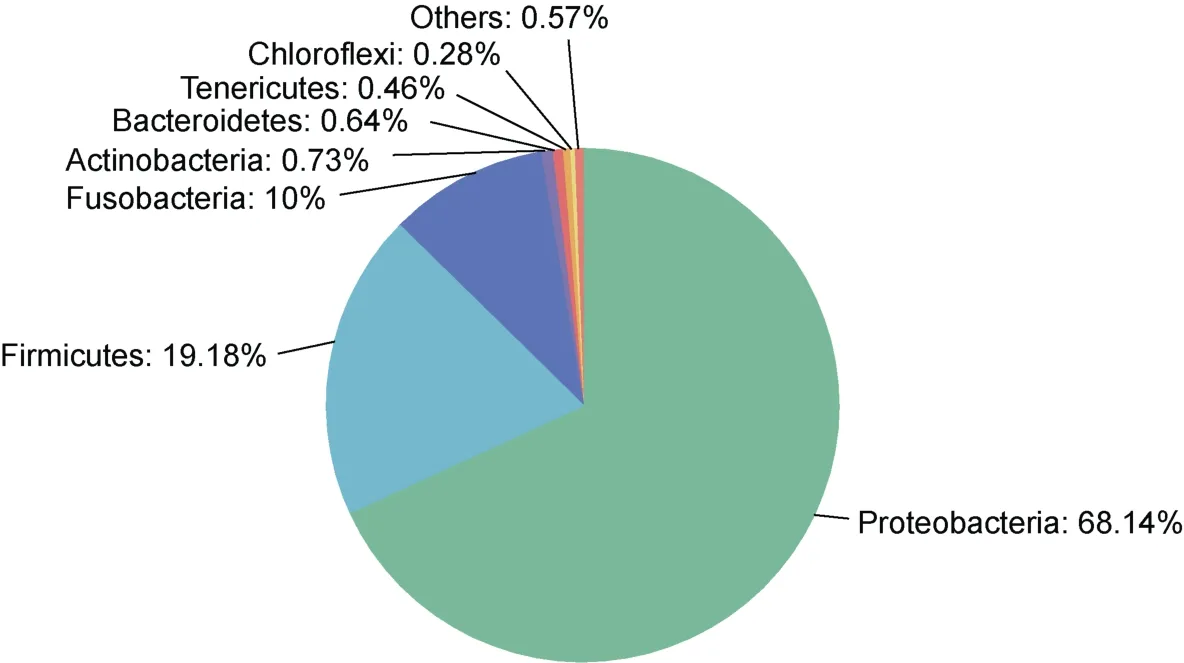
圖2 石斑魚腸道微生物在門水平的組成(相對豐度>0.1%)Fig.2 Composition of gut microbes of grouper at the phylum level (relative abundance > 0.1%)
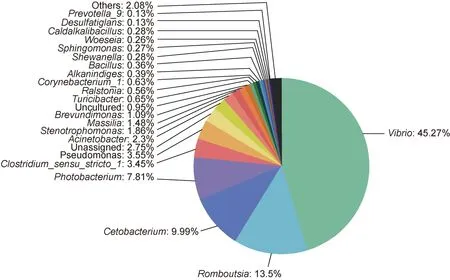
圖3 石斑魚腸道微生物在屬水平的組成(相對豐度>0.1%)Fig.3 Composition of gut microbes of grouper at the the genus level (relative abundance > 0.1%)
在虎龍雜交斑中, 變形菌門(63.89%)和厚壁菌門(32.65%)是其腸道中的優(yōu)勢門類, 而在斜帶石斑魚腸道中為變形菌門(73.72%)和梭桿菌門(22.72%) (圖4a)。在屬類水平中, 虎龍雜交斑的優(yōu)勢屬類為弧菌屬(52.39%)、羅姆布茨菌屬(24.74%)和狹義梭菌屬1 (Clostridium sensustricto1, 5.94%), 斜帶石斑魚的優(yōu)勢屬類為弧菌屬(40.04%)、鯨桿菌屬(22.71%)和發(fā)光桿菌屬(Photobacterium, 15.00%) (圖4b)。
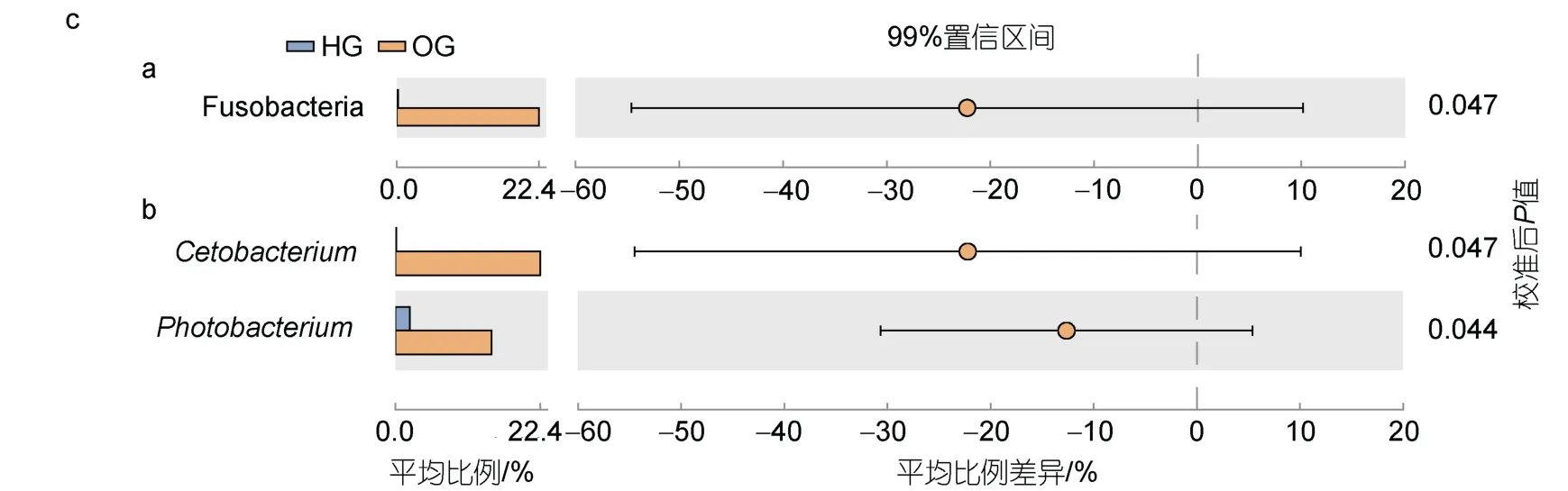
圖4 虎龍雜交斑和斜帶石斑魚的腸道微生物組成Fig.4 Composition of gut microbes of hybrid grouper and orange-spotted grouper
與虎龍雜交斑相比, 斜帶石斑魚腸道中梭桿菌門、鯨桿菌屬和發(fā)光桿菌屬的相對豐度顯著更高(P<0.05,圖4c)。LEfSe 分析表明, 鯨桿菌屬、發(fā)光桿菌屬以及丙酸菌屬(Propionigenium)是斜帶石斑魚腸道中的標志微生物, 虎龍雜交斑腸道中的標志微生物為狹義梭菌屬7 (Clostridiumsensustricto7)、棒狀桿菌屬1(Corynebacterium1)和瘤胃菌屬(Ruminococcaceae UCG008)。
與斜帶石斑魚不同, 虎龍雜交斑三個標志性屬類的相對豐度較低, 且其腸道內的標志微生物源自更多不同的分類分支(圖5)。此外, NMDS 顯示虎龍雜交斑和斜帶石斑魚腸道微生物群落基本分離(圖6)。

圖5 虎龍雜交斑和斜帶石斑魚的LDA 得分圖(a)和LEfSe 分支圖(b)Fig.5 Histogram of the LDA scores (a) and the LEfSe cladogram (b) of hybrid grouper and orange-spotted grouper

圖6 虎龍雜交斑和斜帶石斑魚的非度量多維尺度分析Fig.6 Non-metric multidimensional scaling (NMDS) analysis of gut microbes of hybrid grouper and orange-spotted grouper
2.3 腸道微生物群落的共現(xiàn)網(wǎng)絡分析
采用微生物相關性網(wǎng)絡分析, 評估腸道微生物群落的相互作用。虎龍雜交斑腸道中微生物的相互作用以一個模塊為主, 而斜帶石斑魚中包含多個孤立模塊(圖7a, 7b)。據(jù)圖7c、圖7d 可見, 變形菌門主導著兩種石斑魚的腸道微生物網(wǎng)絡。此外, 厚壁菌門也在虎龍雜交斑腸道微生物網(wǎng)絡中發(fā)揮著較重要的作用(圖7c)。
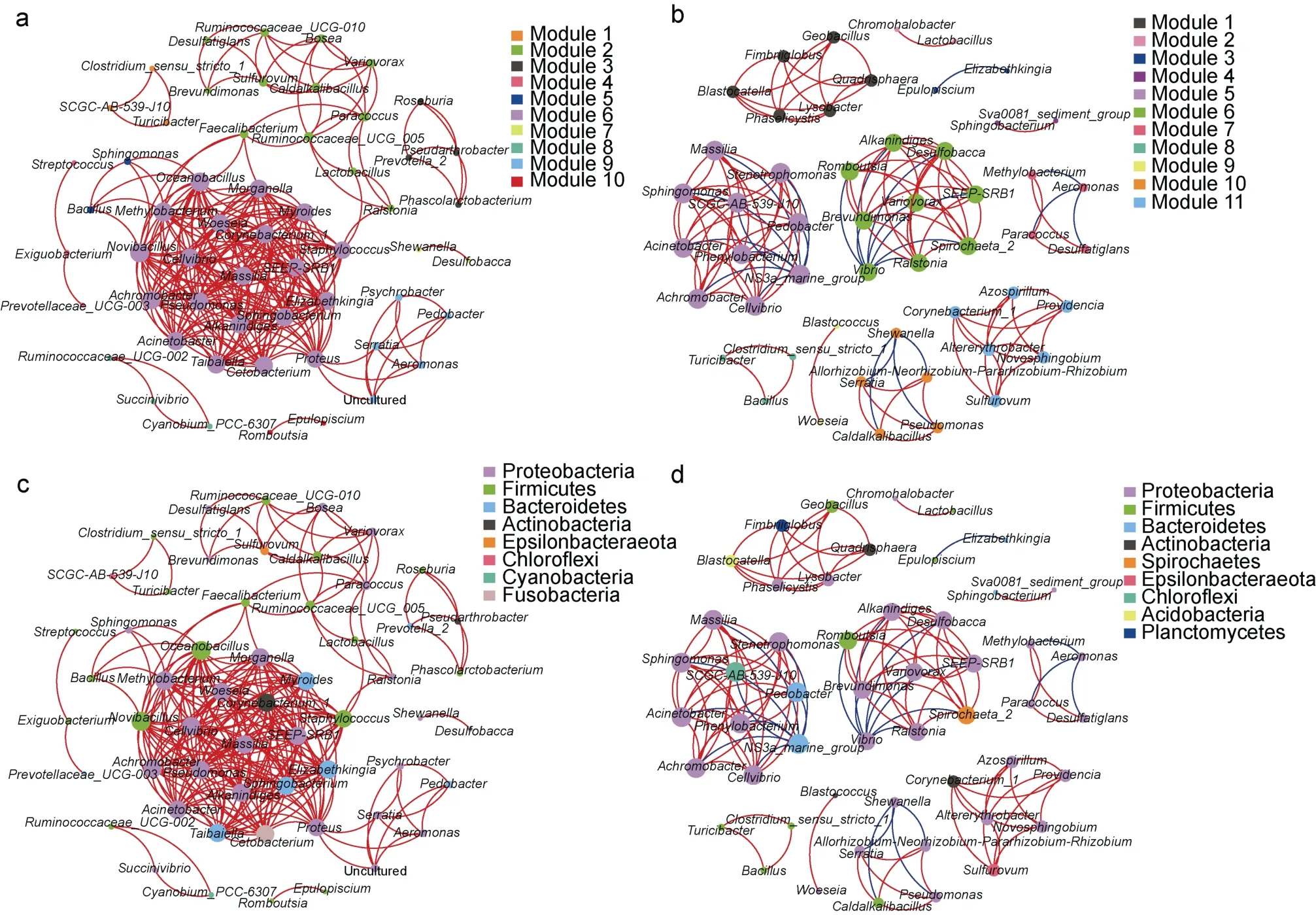
圖7 虎龍雜交斑(a、c)和斜帶石斑魚(b、d)的腸道微生物共現(xiàn)網(wǎng)絡Fig.7 Co-occurrence network of gut microbes in hybrid grouper (a, c) and orange-spotted grouper (b, d)
2.4 腸道微生物群落功能預測
PCA 顯示, 虎龍雜交斑和斜帶石斑魚腸道微生物群落的功能基本分離。PCA 中, PC1 占總變異的77.9%, PC2 占總變異的10.7%, 兩個PCA 軸解釋了虎龍雜交斑和斜帶石斑魚腸道微生物群落之間88%以上的總變異(圖8a)。

圖8 虎龍雜交斑和斜帶石斑魚腸道微生物的預測功能Fig.8 The predicted function of gut microbes of hybrid grouper and orange-spotted grouper
圖8b 展示了基于2 級KEGG 通路的微生物群落功能組成, 結果顯示虎龍雜交斑和斜帶石斑魚腸道微生物群落的功能組成類似, 主要由碳水化合物、輔因子和維生素、氨基酸、萜類化合物與聚酮類化合物、其他氨基酸和脂類代謝組成, 占比將近60%。基于3級KEGG 通路, 本研究發(fā)現(xiàn)了兩種石斑魚腸道中具有顯著差異的功能通路。抗壞血酸和糖醛酸代謝、植物-病原體相互作用和胰島素信號通路在虎龍雜交斑腸道中顯著更高, 而斜帶石斑魚中以萬古霉素類抗生素和鏈霉素的生物合成、亞油酸代謝、雙酚降解和類黃酮的生物合成通路顯著更高(圖8c, 表2)。
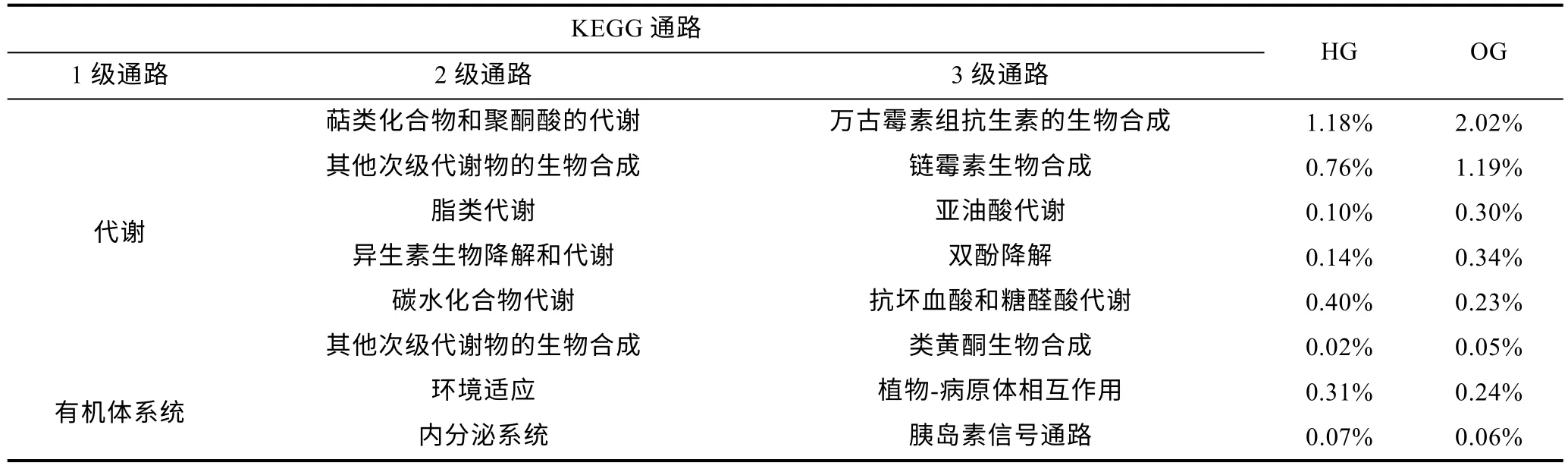
表2 虎龍雜交斑和斜帶石斑魚具有顯著性差異的腸道微生物功能Tab.2 Significantly different functions of gut microbes between hybrid grouper and orange-spotted grouper
3 討論
腸道微生物能夠參與營養(yǎng)代謝并影響宿主免疫系統(tǒng), 在宿主健康中發(fā)揮著重要的作用, 其組成受到環(huán)境因素、攝食和宿主遺傳學的影響(O'Haraetal,2006; Marquesetal, 2010; Zhangetal, 2019; Lietal,2023)。腸道微生物的比較分析在疾病防控(Daietal,2020; Wuetal, 2021b; Mondaletal, 2022)、環(huán)境適應(Linetal, 2022)以及攝食管理(Pengetal, 2019;Salas-Leivaetal, 2020)方面具備重要意義, 相關研究在林氏異腕蝦(Heterocarpushayashi)、長鰭鰤 (Seriola rivoliana)、凡納濱對蝦(Litopenaeusvannamei)、黃鱔(Monopterusalbus)及克氏原螯蝦(Procambarusclarkia)中均有報道, 但對石斑魚腸道微生物群落的研究甚少。本研究對兩種具有重要經(jīng)濟價值的養(yǎng)殖石斑魚品種的腸道微生物群落進行了表征, 并對其共現(xiàn)模式和預測功能進行了探究。
以往許多研究表明, 變形菌門和厚壁菌門在海洋魚類腸道微生物群落中占主導地位(Dongetal,2020; Johnyetal, 2022; Yuetal, 2022)。本研究結果顯示, 石斑魚腸道微生物群落的優(yōu)勢門類為變形菌門、厚壁菌門以及梭桿菌門, 與花斑刺鰓 鮨(Plectropomusleopardus)、虹鱒魚(Oncorhynchusmykiss)、軍曹魚(Rachycentroncanadum)、尖吻鱸(Latescalcarifer)和布氏 鯧鲹 (Trachinotusblochii)的結果相一致(Michlet al, 2017; Rasheedaetal, 2017; Mekuchietal, 2018)。但兩種石斑魚腸道的優(yōu)勢門豐度有所不同, 兩種石斑魚腸道中變形菌門的相對豐度最高; 其次, 在虎龍雜交斑中相對豐度較高的門是厚壁菌門(32.65%)和放線菌門(1.03%), 在斜帶石斑魚中則是梭桿菌門(22.72%)和厚壁菌門(2.49%)。兩個石斑魚品種腸道微生物群落中門水平相對豐度的差異, 表明腸道微生物群落的組成可能與宿主的遺傳背景相關。Li 等對四種重要經(jīng)濟亞洲鯉魚的腸道微生物群落的研究揭示了亞洲鯉魚腸道微生物群落結構的影響因素, 其研究表明, 即使不同的物種生存在同一環(huán)境中, 各物種的腸道微生物群落仍具有物種特異性(Lietal, 2018)。對匙吻鱘(Polyodonspathala)和鳙魚(Aristichthysnobilis)的研究也得出了類似的結果, 宿主遺傳背景可能是導致同一養(yǎng)殖環(huán)境中攝食相似的養(yǎng)殖物種腸道微生物群落存在差異的主要因素(Lietal, 2014)。
值得注意的是, 在屬水平上, 兩個石斑魚品種均以弧菌屬為相對豐度最高的屬。在大西洋鱈(Gadusmorhua)、凡納濱對蝦、軍曹魚和布氏鯧 鲹等養(yǎng)殖物種中也曾觀察到較高的弧菌屬豐度(Reidetal, 2009;Rasheedaetal, 2017; Zoqrattetal, 2018)。弧菌屬常作為具有巨大影響力的致病菌而廣受關注, 副溶血性弧菌(Vibrioparahaemolyticus)、溶藻弧菌(V.alginolyticus)和哈維弧菌(V.harveyi)等常見的弧菌屬致病菌可引起海洋魚類和貝類嚴重疾病(Ina-Salwanyetal, 2019;Trigaetal, 2023)。早期的數(shù)據(jù)顯示, 石斑魚易受弧菌屬感染(Ina-Salwanyetal, 2019)。在本研究中, 石斑魚腸道中含有較高豐度的弧菌屬卻未見疾病的發(fā)生,可能與弧菌屬致病菌的條件致病性以及弧菌屬存在潛在益生菌相關(Thompsonetal, 2004; Egertonetal,2018; Zhuetal, 2023)。Ring? 等已分離出能夠大量產(chǎn)生二十碳五烯酸的海弧菌(Vibriopelagius) (Ring?et al, 1992), Zhu 等(2023)則已分離出可作為飼料添加劑增強宿主免疫能力的河流弧菌(Vibriofluvialis) (Zhuetal, 2023)。此外, 也有研究表明, 弧菌屬可以分泌淀粉酶、蛋白酶、卵磷脂酶和幾丁質酶, 幫助宿主消化體內的營養(yǎng)物質, 如脂肪、蛋白質和碳水化合物(Hansenetal, 1999)。總而言之, 弧菌屬在石斑魚腸道中的作用機制仍有待進一步研究。
斜帶石斑魚腸道中的鯨桿菌屬及發(fā)光桿菌屬相對豐度較高, 與虎龍雜交斑相比差異顯著, 是LEfSe證實的腸道標志微生物。已有報道顯示, 鯨桿菌屬具有產(chǎn)生維生素B12的能力, 并且對魚類的葡萄糖穩(wěn)態(tài)具有有益作用(Wangetal, 2021; Zhangetal, 2022a)。維生素B12在DNA 合成與調節(jié)、神經(jīng)系統(tǒng)功能、紅細胞形成等生物過程中至關重要。發(fā)光桿菌屬則與弧菌屬相同, 是具有生產(chǎn)多不飽和脂肪酸能力的主要微生物之一(Moietal, 2018)。虎龍雜交斑腸道中相對豐度較高的屬與標志微生物不相一致, 相對豐度較高的羅姆布茨菌屬可能在維持宿主腸道健康狀況方面發(fā)揮關鍵作用(Mangifestaetal, 2018)。羅姆布茨菌屬主要利用單糖和雙糖形成乙酸、甲酸和乳酸, 通過上調或下調多種代謝產(chǎn)物以達到調控宿主免疫功能的作用(Songetal, 2022)。另一相對豐度較高的狹義梭菌屬 1 則被認為能夠抑制條件致病性埃希菌屬(Escherichia)細菌(Liangetal, 2023)。
虎龍雜交斑腸道中的標志微生物棒狀桿菌屬1、變形桿菌屬(Proteus)、葡萄球菌屬(Staphylococcus)以及類香菌屬(Myroides)值得關注, 人們通常將其視為致病菌(Hamiltonetal, 2018; Ningetal, 2020; Beathardetal, 2021; Lisowska-?ysiaketal, 2021)。虎龍雜交斑具備的較強抗病性可能是本研究中未出現(xiàn)疾病或死亡的原因(Tanetal, 2018)。值得注意的是, 一些研究顯示上述菌屬的物種在病原菌抑制、水體凈化、宿主保護等方面也存在一定的功效, 可能有助于虎龍雜交斑抗病能力的增強(Nayaketal, 2011; Hamzaetal,2018; Apines-Amaretal, 2022)。此外, 共現(xiàn)網(wǎng)絡顯示,棒狀桿菌屬1 和類香菌屬是網(wǎng)絡中主要模塊的重要組成部分, 變形桿菌屬及葡萄球菌屬則起到連接模塊的作用, 它們的生長與代謝受到其他各類微生物的影響, 這可能使其穩(wěn)定在一定水平而不致引起相關疾病。因此, 該四種菌屬的有益功能可能是通過連接及豐富微生物網(wǎng)絡, 使虎龍雜交斑腸道微生物群落更加復雜, 幫助虎龍雜交斑適應更廣泛的生存環(huán)境(Pengetal, 2019)。總之, 虎龍雜交斑腸道標志微生物與宿主生理之間的關系值得進一步探索。
PICRUSt 通過將測序數(shù)據(jù)與參考基因組進行比較來預測微生物群落的功能潛力(Langilleetal, 2013)。根據(jù)預測, 最豐富的功能類別是與碳水化合物、輔因子和維生素以及氨基酸代謝相關的功能類別, 虎龍雜交斑與斜帶石斑魚的KEGG 通路存在顯著差異。在虎龍雜交斑腸道中, 抗壞血酸和糖醛酸代謝與胰島素信號通路更為豐富。抗壞血酸和糖醛酸代謝是抗氧化防御相關途徑的組成部分, 參與超氧化物的清除, 有助于增強免疫系統(tǒng)(Bietal, 2023)。此外, 抗壞血酸在提高存活率和生長性能方面也具有一定的作用(Liaoetal, 2023)。在魚類中, 胰島素信號通路參與葡萄糖攝取和糖原、脂質與蛋白質的合成, 在調節(jié)細胞生長和分化方面具重要意義(Jiaetal, 2022)。因此,虎龍雜交斑腸道中上述兩個通路的富集可能會進一步增強其生長表現(xiàn)和抗病性。不同的是, 在斜帶石斑魚中, 與亞油酸代謝、雙酚降解相關的KEGG 通路顯著富集。亞油酸是魚類必需的膳食營養(yǎng)素(Glencross,2009), 而接觸雙酚化合物(BPs), 包括雙酚A (BPA)、雙酚AF (BPAF)、雙酚F (BPF)和雙酚S (BPS)等可能會對水生脊椎動物的發(fā)育、生殖和性別比例產(chǎn)生影響(Jiaoetal, 2010; Zhangetal, 2022b; Huangetal,2023)。值得注意的是, 在斜帶石斑魚腸道中萬古霉素類抗生素和鏈霉素的生物合成以及類黃酮生物合成相關通路也被顯著富集, 這些通路均與抗菌相關, 可能會增加腸道微生物群對病原菌的抑制能力。然而,由于功能通路分析是根據(jù)微生物群落預測的, 因此需要進一步分析以確定腸道微生物的功能。
4 結論
本研究揭示了虎龍雜交斑和斜帶石斑魚腸道微生物的組成及功能差異。在兩種石斑魚腸道中變形菌門、厚壁菌門和梭桿菌門占主導地位。在屬水平, 豐度較高的鯨桿菌屬和發(fā)光桿菌屬是斜帶石斑魚腸道中的標志微生物, 虎龍雜交斑腸道標志微生物為相對豐度較小的棒狀桿菌屬1、變形桿菌屬、葡萄球菌屬和類香菌屬, 該四種菌屬的作用可能是增加腸道微生物群落的復雜性, 進而增強虎龍雜交斑的環(huán)境適應性。功能預測顯示, 腸道微生物群可能與虎龍雜交斑較好的生長表現(xiàn)以及抗病性相關, 而斜帶石斑魚需要腸道微生物參與亞油酸代謝、雙酚降解以及抗菌物質合成。總的來說, 本研究支持了腸道微生物群落結構與宿主遺傳背景相關的觀點, 加深了對養(yǎng)殖石斑魚腸道微生物群落的認識, 為后續(xù)的宿主相關益生菌篩選、養(yǎng)殖品種選擇提供了參考。
