扁桃耐旱砧木木質部解剖結構與栓塞特性的關系分析
2024-02-22 00:00:00于秋紅許盼云郭春苗迪利夏提·哈斯木木巴熱克·阿尤普
新疆農業科學 2024年11期

摘"要:【目的】""基于莖木質部解剖結構與栓塞特性關系分析扁桃耐旱砧木資源對干旱脅迫環境的適應機理。
【方法】""以大巴旦(Amygdalus communis var.fragilis)(C1)、苦巴旦(A.communis var.amara)(C2)、甜仁桃巴旦[A.communis var.persicoides (West.) Rehd.](C3)和苦仁桃巴旦[A.communis var.persicoides (West.) Rehd.](C4)以及新疆毛桃(A.persica L.)(C5)等耐旱性程度不同5個扁桃砧木資源的當年生實生苗為材料,采用“外加壓力”法,配套使用“XYLEM木質部導水率及栓塞測量系統”和“PMS空穴壓力室”,測定在正常栽培條件下的莖自然栓塞程度以及栓塞脆弱性值(P50);測量和觀察已栓塞枝條不同時段(30,80,130,180 min)的栓塞修復程度以及木質部解剖結構特征,探究耐旱性不同扁桃砧木資源的莖木質部解剖結構、栓塞抗性以及栓塞修復能力間的關系。
【結果】""(1) 扁桃不同砧木資源中大巴旦、甜仁桃巴旦、苦巴旦和普通桃的栓塞脆弱曲線為“s”形,苦仁桃巴旦的栓塞脆弱曲線為“r”形;(2) 莖木質部栓塞抗性(P50)平均值由大到小依次為大巴旦gt;甜仁桃巴旦gt;苦巴旦gt;苦仁桃巴旦gt;普通桃,其中大巴旦的抗栓塞能力最強;(3) 莖木質部栓塞修復能力由大到小依次為普通桃gt;甜仁桃巴旦gt;苦仁桃巴旦gt;苦巴旦gt;大巴旦;(4) 較耐旱砧木大巴旦、苦巴旦和甜仁桃巴旦的木質部導管及微孔直徑小,木材密度、導管壁厚度及導管壁理論機械強度大,抗栓塞能力強,然而栓塞修復能力弱;‘普通桃’和苦仁桃巴旦的導管及微孔直徑大,木材密度、導管壁厚度以及導管壁理論機械強度小,抗栓塞能力弱,但栓塞修復能力強。
【結論】""扁桃砧木資源木質部栓塞修復能力與栓塞抗性呈負相關,與木質部解剖結構中的導管及微孔直徑呈正相關,與木材密度、導管壁厚度以及導管壁理論機械強度呈負相關。
關鍵詞:""扁桃;砧木資源;栓塞抗性;栓塞修復;木質部解剖結構
中圖分類號:"S662.9""""文獻標志碼:"A""""文章編號:"1001-4330(2024)11-2693-12
0"引 言
【研究意義】木質部栓塞(Xylem embolism)是一種在干旱條件下木本植物的木質部導管產生水分運輸功能受損的現象,是影響植物正常生長和發育的重要制約因素[1]。植物的木質部空穴化及栓塞會使其輸水功能受到影響,進而影響其耐旱能力,削弱植物防御機制,導致植物死亡[2,3]。依據Domec理論[4],木本植物木質部抵抗栓塞的能力可以用木質部導水率損失最大值的臨界水勢和P50值來表示[5]。在干旱地區,植物體內缺乏水分,木質部栓塞現象較為普遍[6]。耐旱性是植物感知水分缺失信號并啟動應對策略的能力,在一定程度上可以由栓塞特性所決定[7-9]。研究發現,對植物重新供水后,木質部水分壓力增大,導管內的氣泡脹破,溶解在周圍的溶液中,木質部栓塞可以得到修復[10]。栓塞修復率可以通過復水前后的導水率差值來表示,以此來描述栓塞修復能力[11,12]。植物木質部的抗栓塞能力和栓塞修復能力與其耐旱性關系密切,研究植物栓塞特性與木質部解剖結構間的關系,對于深入分析其適應干旱環境的生理生態機制的認識具有重要意義。【前人研究進展】目前木質部解剖結構與栓塞特性間的關系越來越成為研究植物耐旱性的熱點[13,14]。木質部結構中的導管直徑、密度、內徑跨度以及紋孔結構等因素均對水分運輸有重要影響[15,16]。例如木質部導管的直徑越大,其內部的水分壓力也就越高,當水分壓力高到一定程度時,可能會導致導管內部的氣泡凝聚形成氣栓,從而引發栓塞[16,17]。黨維等[18]在6個樹種1年生枝木質部水力特征和栓塞修復能力的關系研究中發現植物木質部水力特征與栓塞修復能力有較強的關聯,特別是栓塞脆弱性(P50)和水力直徑是影響栓塞形成和修復的重要因素,其與導管的結構和功能密切相關;李美琪等[19]對楊樹(Populus spp.)進行水分脅迫盆栽試驗發現,水分脅迫程度增強從而引起木質部的水勢的下降,導致木質部導水率減少,因此栓塞脆弱性降低,提高了植物抵抗栓塞的能力。【本研究切入點】扁桃(Amygdalus communis Linn.)屬于薔薇科桃屬植物[20]。目前新疆扁桃最常用的砧木是普通桃,該砧木具有抗旱性較差等缺陷,對扁桃產業的可持續性影響較大,評價和篩選耐旱扁桃砧木資源一直是扁桃砧木育種的重要研究方向之一[21-23]。【擬解決的關鍵問題】參考以往的扁桃砧木耐旱性評價研究結果[24],選擇大巴旦、苦巴旦、甜仁桃巴旦、苦仁桃巴旦和普通桃等耐旱性差異較明顯的代表性扁桃砧木資源為材料,比較扁桃不同砧木資源木質部栓塞抗性、栓塞修復程度以及木質部解剖結構之間的差異,分析其與耐旱性間的關系,為扁桃耐旱砧木育種以及深入了解扁桃耐旱砧木的干旱適應策略提供理論參考。
1"材料與方法
1.1"材 料
試驗地選取在新疆烏魯木齊市新疆農業科學院園藝作物研究所試驗基地(43°49′13″ N, 87°34′47″ E)。該地區氣候類型為中溫帶大陸性干旱氣候,年均溫4~13℃,最熱月(7月)平均溫度30℃,極端高溫37℃,極端低溫9℃;最冷月(1月)平均溫度-15.2℃;年均降水量在194 mm左右。大巴旦、苦巴旦、甜仁桃巴旦、苦仁桃巴旦和普通桃種子均來自新疆喀什地區莎車縣國有二林廠扁桃種質資源圃。各砧木資源分別選擇100粒種子,在2022年2~4月期間室內育苗,4月初均移栽在直徑25 cm、高32 cm的圓柱形花盆中,每盆1株。花盆土壤以沙土和沙壤土為主,土壤有機質含量為207.74 "g/kg,全氮含量為6.62 "g/kg,全磷含量為0.80 "g/kg,全鉀含量為15.45 "g/kg,pH值為7.55。從各砧木資源選擇生長健壯、長勢一致的實生苗50盆,所有苗木放入四周通風通光的試驗地,一直到9月中旬進行正常栽培管理。栽培期間,使所有盆栽土壤含水量達到飽和狀態(達到田間持水量的75%~80%),每隔5~7 d澆1次水,每2 d測一次土壤含水量(使用SM 150T便攜式土壤水分速測儀,英國,Delta公司,測定土壤電導率,參照儀器使用說明書計算土壤含水量)),使土壤含水量保持在田間最大持水量的75%~80%,保持樣株生長旺盛。表1
1.2"方 法
1.2.1"栓塞脆弱性曲線的建立
試驗始于2022年9月中旬,此時所有樣株木質部已硬化,基本停止生長。試驗當日06:00,在每個砧木資源中選取長勢基本一致且主莖直徑為6~8 mm的樣株5株;剪取樣株主莖的地上部分,在主莖上部套上黑色塑料袋,底部置于水中;在水中切除枝條基部2~3 cm的莖段,切取主莖中部長約15 cm的莖段,用“外加壓力”法,配套使用“XYLEM木質部導水率及栓塞測量系統”和“PMS空穴壓力室”建立栓塞脆弱性曲線[25]。
1.2.2"栓塞修復能力的測定
從每個砧木資源中選取長勢基本一致且主莖直徑為6~8 mm的樣株4株,從每個樣株上切取長20 cm的主莖,并對其進行人為誘導木質部栓塞,使其導水率損失值(PLC)達到95%以上。之后將莖水平放置于水中,經過30、80、130和180 min的恢復后,在靠近基部的一側從中部截取5個連續的2 cm莖段(需在水下完成枝條的截取),隨后使用“XYLEM木質部導水率及栓塞測量系統”測定其導水率的損失百分比(PLC)[6,18]。
木質部栓塞的修復能力可以通過導水率修復百分比來量化表示。
導水率修復百分比(%)=1-PLC=1-100(Kmax-Kh)/Kmax.
式中, Kh代表截段初始的導水率值,而Kmax是指截段在0.18 MPa壓力下經過反復沖洗后達到的最大導水率值。對于每種扁桃砧木資源,在每個時間段內5次重復測量,取測量結果平均值表示該資源木質部栓塞的修復程度。
1.2.3"莖木質部解剖結構的測定
用于建立栓塞脆弱性曲線的莖段測定完成后,切取莖段中部相同位置的長度1~2 cm小段,采用番紅-固綠染色法制成石蠟切片,切片厚度8~10 μm。每個扁桃砧木資源隨機選取5個切片,使用Olympus BX51光學顯微鏡進行觀察和拍照,采用Image-J圖像處理軟件測量木質部導管直徑(Vessel diameter, D)、連接導管壁厚度(Intervessel wall thickness, TVW)、單導管指數(Solitary vessel index, VS)、導管密度(Vessel density, VD)、導管組指數(Vessel grouping index, VG)、導管壁理論機械強度(Theoretical vessel implosion resistance, (TVW/D連接導管)2)、導管水力直徑(Hydraulic diameter of vessel, Dh)和木材密度(Wood density, WD)[26]。在剩余莖段中部切取長度5-10 mm的小段,參考Jansen等[27]方法在不同濃度梯度的乙醇溶液中脫水,于室內自然風干,將樣品縱切后固定在樣品架上,噴白金3 min;采用Zeiss Supar 55 (VP)掃描電子顯微鏡(德國,Carl Zeiss公司)在2 kV電壓下觀察紋孔膜結構并拍照,采用Image-J圖像處理軟件測量紋孔膜面積(Pit membrane area, Pa)、紋孔膜直徑(Pit membrane diameter, Pd)、微孔面積(Microporous area, Ma)和微孔直徑(Microporous diameter, Md)。
1.3"數據處理
利用SPSS 19.0 (SPSS Inc., Chicago, IL, USA)和Origin 2022 (OriginLab Corporation, Northampton, MA, USA)分析數據和繪圖,分析前需滿足正態分布和方差齊性。采用單因素方差分析自然栓塞度(PLC)、栓塞脆弱性(P50)、栓塞修復能力以及木質部解剖結構在扁桃不同砧木資源間的差異,采用LSD法進行差異顯著性分析,Plt;0.05表示差異性顯著。
2"結果與分析
2.1"扁桃砧木資源莖木質部栓塞抗性的差異
2.1.1"莖栓塞脆弱性曲線的建立
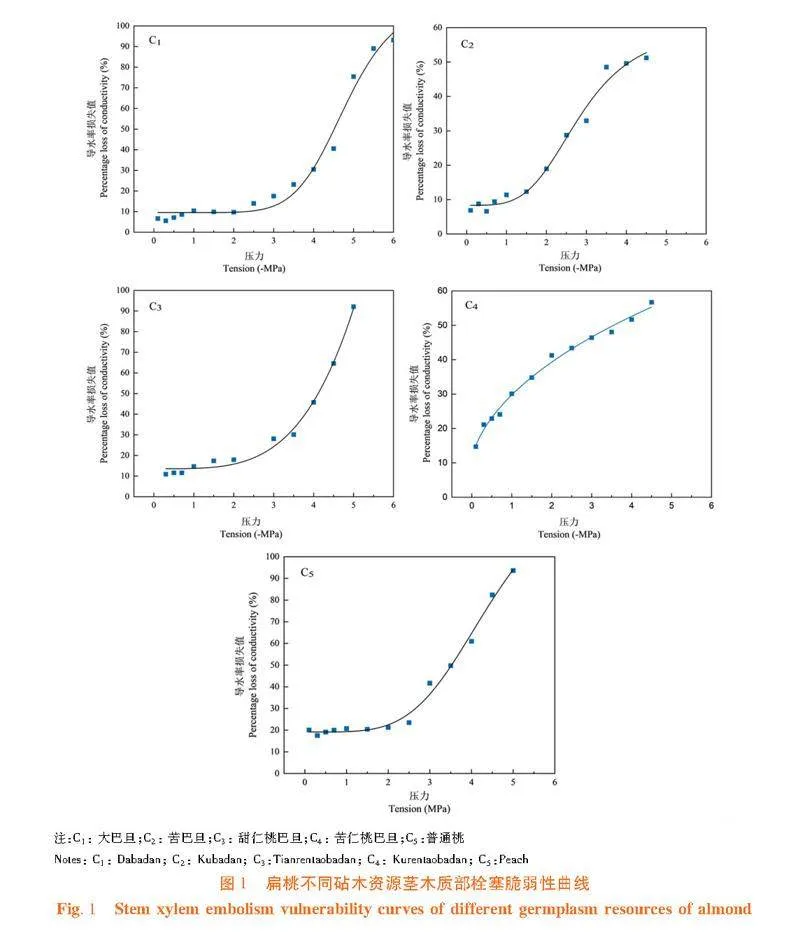
研究表明,苦仁桃巴旦的莖對木質部栓塞較敏感,在木質部壓力出現負值的初始階段,枝條木質部導水率損失值迅速增大,且隨著水勢的不斷降低,木質部導水率損失百分數(PLC%)即栓塞化程度迅速增大,呈典型的“"r"”形。在木質部負壓為-3.55 MPa時,其導水率損失值約為50%,達到最大導水率損失值時的木質部壓力約為-6 MPa。大巴旦和苦巴旦莖木質部栓塞脆弱曲線呈典型的“S”形,當木質部壓力大于-2.0 MPa時,其導水率損失值很小,木質部負壓達到-2.0 MPa時,導水率損失值迅速增大,當導水率損失值達到最大時,木質部壓力約為-5~-6 MPa。苦巴旦莖木質部壓力大于-1.5 MPa時,導水率損失值幾乎不變,但當壓力處于-4.0~-1.5 MPa時,木質部導水率損失值迅速增大,木質部發生較大程度的栓塞。甜仁桃巴旦和普通桃的莖木質部壓力大于-2 MPa時,導水率損失值幾乎不變,但當壓力處于-5.0~-2.0 MPa時,木質部導水率損失值迅速增大,木質部發生較大程度的栓塞。大巴旦的莖木質部壓力大于-3.0 MPa時,導水率損失值幾乎不變,僅發生較小程度的栓塞,當壓力處于-5.5~-3.0 MPa時,木質部導水率損失值迅速增大,木質部發生較大程度栓塞。圖1"
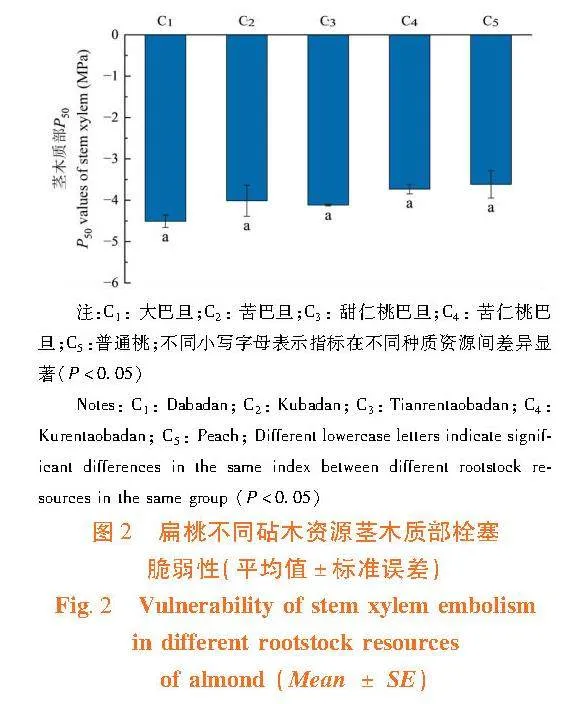
2.1.2"莖木質部栓塞抗性的差異
研究表明,扁桃不同砧木資源莖栓塞脆弱性之間差異不顯著(Pgt;0.05)。各資源莖木質部栓塞脆弱性程度高到低為:普通桃(3.62 ± 0.33)gt;苦仁桃巴旦(3.73 ± 0.11)gt;苦巴旦(4.01 ± 0.38)gt;甜仁桃巴旦(4.12 ± 0.02)gt;大巴旦(4.51 ± 0.15) 。植物木質部的栓塞脆弱性大,栓塞抗性小,在水分運輸過程中植物木質部易發生栓塞,對干旱脅迫的抵御能力差。因此,所有供試資源中普通桃的抗栓塞能力最弱,大巴旦的抗栓塞能力最強。圖2
2.2"扁桃砧木資源莖木質部栓塞修復能力差異
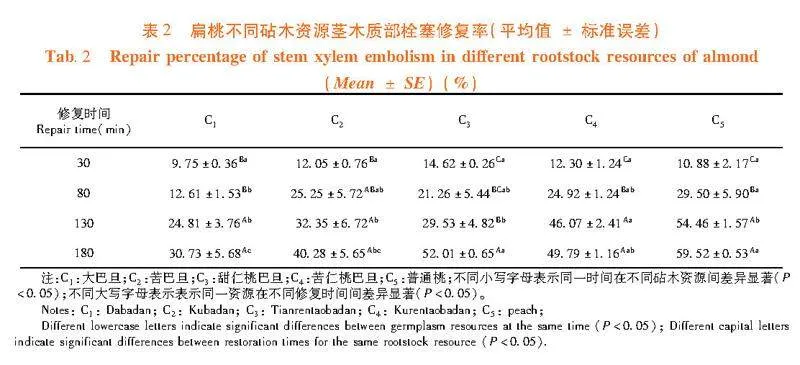
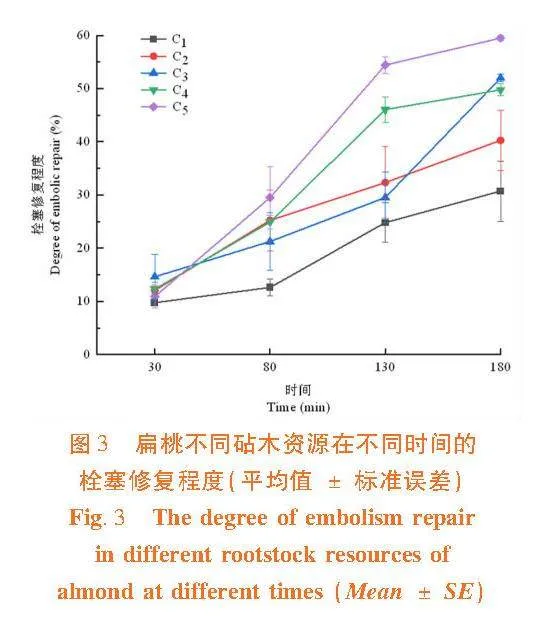
研究表明,在栓塞修復初期30 min時,甜仁桃巴旦的栓塞修復程度相對較好,而大巴旦的栓塞修復程度相對較慢。在栓塞修復中期80 min時,普通桃的栓塞修復程度相對較好,大巴旦的栓塞修復程度相對較慢;在栓塞修復進行130 min時,普通桃和苦仁桃巴旦的栓塞修復程度均較好。除普通桃和苦仁桃巴旦外,其余3個砧木資源在修復130 min時的栓塞修復程度差異不顯著;在栓塞修復后期180 min時,栓塞修復程度最好的是普通桃,而大巴旦的修復程度最弱。在栓塞修復180 min時,“s”形曲線的大巴旦、苦巴旦、甜仁桃巴旦、普通桃和“r”形脆弱性曲線的苦仁桃巴旦之間栓塞修復程度存在顯著性差異(Plt;0.05)。表2
木質部栓塞的修復程度逐漸提高的趨勢。在修復30 min時,甜仁桃巴旦的修復程度大于"苦仁桃巴旦、苦巴旦和普通桃,但在修復"80~130 min時甜仁桃巴旦的修復程度要低于苦仁桃巴旦、苦巴旦和普通桃,甜仁桃巴旦在短期內修復程度較優,而苦仁桃巴旦、苦巴旦和普通桃在中期修復程度較甜仁桃巴旦優;在修復130~180 min時甜仁桃巴旦的修復程度要高于苦仁桃巴旦和苦巴旦,甜仁桃巴旦在長時間內修復程度較苦仁桃巴旦和苦巴旦要優;大巴旦在各個時間段其栓塞修復程度均較弱,顯著低于其它砧木資源。
扁桃各砧木資源在180 min時的栓塞修復程度間差異顯著(Plt;0.05),其木質部栓塞修復能力強到弱為普通桃gt;甜仁桃巴旦gt;苦仁桃巴旦gt;苦巴旦gt;大巴旦。即植物重新供水后,木質部栓塞修復能力大的砧木資源,其栓塞導管能迅速恢復其輸水功能。圖3
2.3"扁桃砧木資源莖木質部栓塞特性與解剖結構間的關系"
2.3.1"莖木質部解剖結構的差異
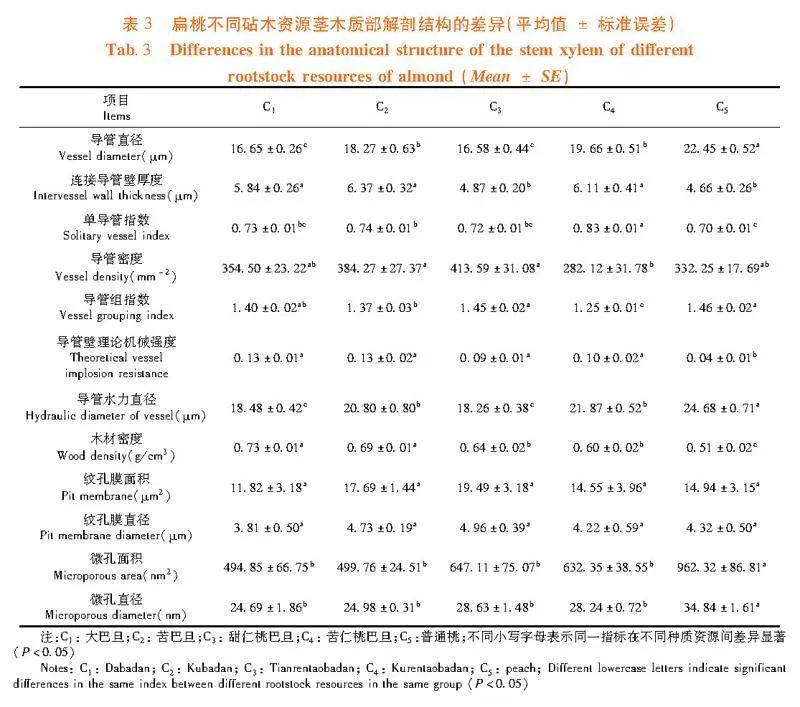
研究表明,莖木質部解剖結構的各項指標在扁桃砧木資源間均呈顯著性差異,扁桃種質資源植株的莖木質部栓塞脆弱性曲線可以用單威布爾函數很好地擬合,并且苦仁桃巴旦的栓塞脆弱性"曲線呈典型的“"r"”形,大巴旦和苦巴旦為典型的“S”形曲線,這與前人研究提出的兩種栓塞脆弱性曲線類型相符lt;0.05) 。其中,大巴旦和苦巴旦的(TVW/D連接導管)2均顯著高于其它砧木資源;甜仁桃巴旦的VD顯著高于其它砧木資源(Plt;0.05),但其D和Dh均顯著低于其它扁桃種質資源;苦仁桃巴旦的VD和VG均顯著低于其它扁桃種質資源,而VS較高;普通桃除了D、VG和Dh均顯著高于其它砧木資源外,其它各項莖木質部解剖結構指標均顯著低于其它砧木資源(Plt;0.05)。表3,圖4
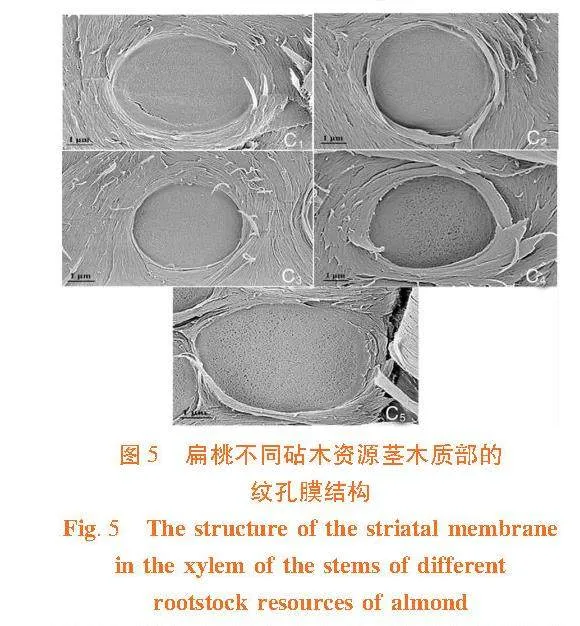
各扁桃砧木資源的莖紋孔膜上均有微孔,微孔的大小在各扁桃砧木資源間具有一定差異。普通桃的Ma和Md顯著高于其它砧木資源,大巴旦顯著低于其它資源(Plt;0.05)。而Pa和Pd在各砧木資源間差異不顯著。圖5
2.3.2"莖木質部栓塞抗性、栓塞修復與解剖結構間的內在關系"
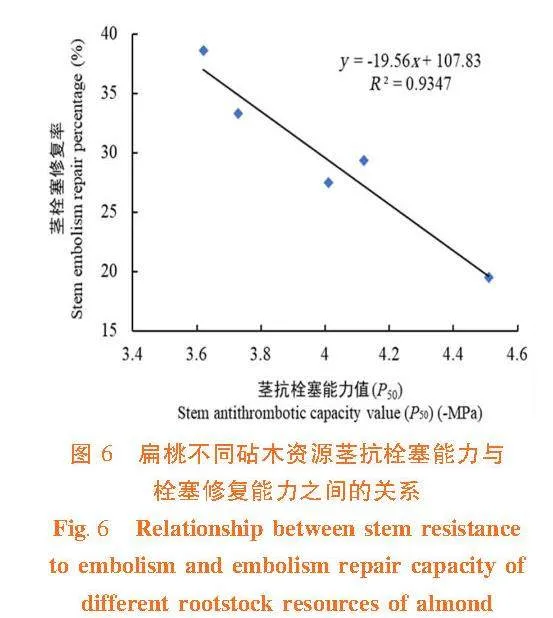
研究表明,扁桃不同砧木資源莖木質部抗栓塞能力(P50)、栓塞修復能力、木材密度和木質部解剖結構之間有較好的相關性。不同砧木莖木質部抗栓塞能力與栓塞修復能力之間具有負相關關系(R2 = 0.934 7),表現為抗栓塞能力強,不易發生栓塞,則栓塞修復能力弱;大巴旦的抗栓塞能力強,卻栓塞修復能力弱,普通桃的抗栓塞能力弱,但栓塞修復能力強。圖6
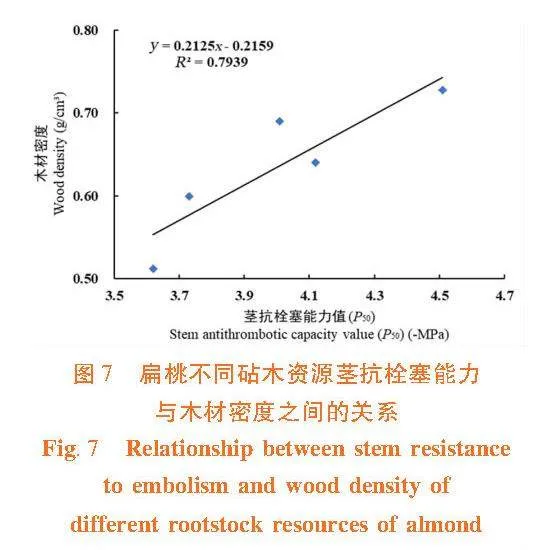
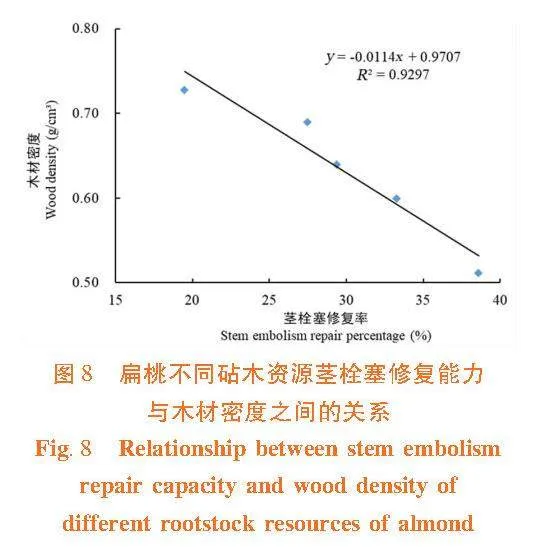
莖木質部抗栓塞能力與木材密度之間呈顯著正相關關系(R2 = 0.793 9) 。木材密度大,栓塞抗性強,植物不易發生栓塞。栓塞抗性強的大巴旦和苦巴旦木材密度明顯高于栓塞抗性弱的甜仁桃巴旦、苦仁桃巴旦和普通桃。并且,莖木質部栓塞修復能力與木材密度間呈負相關關系(R2 = 0.929 7)。由此可見,木材密度是木質部抗栓塞能力和栓塞修復能力權衡關系的調節因子。大巴旦和苦巴旦的木材密度大,抗栓塞能力強,栓塞修復能力弱;普通桃的抗栓塞能力最弱,但它的木材密度小,具有較高的儲水能力,有利于栓塞的木質部恢復輸水功能。圖7,圖8"
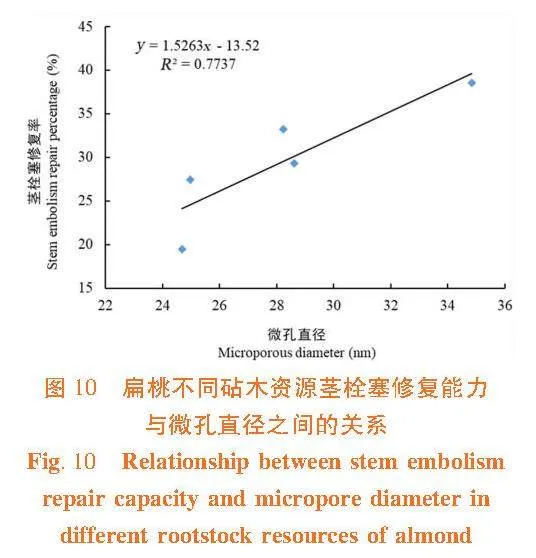
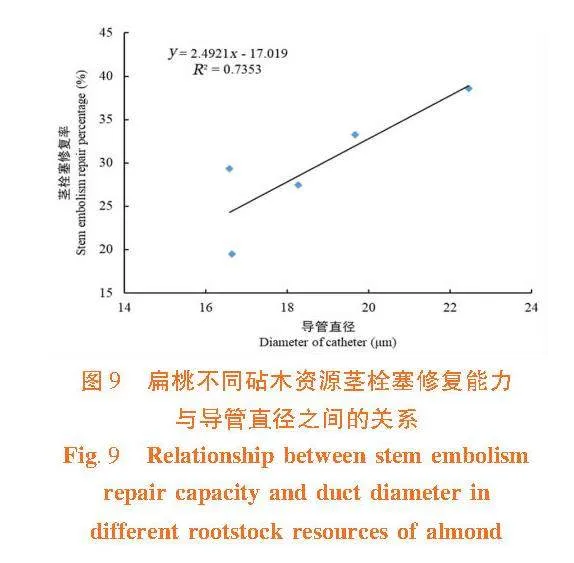
莖木質部栓塞修復能力與木質部導管直徑和紋孔膜上的微孔直徑間呈正相關關系(R2 = 0.735 3,R2 = 0.773 7),莖木質部導管直徑和紋孔膜上的微孔直徑越大,在栓塞修復過程中空氣容易排出或溶解于周圍溶液中,則栓塞越容易修復。 大巴旦的抗栓塞能力在5個砧木資源中最強,導管直徑和紋孔膜上的微孔直徑最小,栓塞修復能力最弱,在缺水環境下大巴旦莖木質部不容易發生栓塞,保持一定的導水能力,但一旦發生栓塞,其不易修復。普通桃的抗栓塞能力最弱,導管直徑和紋孔膜上的微孔直徑最大,栓塞修復能力最強。圖9,圖10
3"討 論
3.1"栓塞抗性與木質部解剖結構間的關系
扁桃種質資源植株的莖木質部栓塞脆弱性曲線可以用單威布爾函數很好地擬合,并且苦仁桃巴旦的栓塞脆弱性曲線呈典型的“"r"”形,大巴旦和苦巴旦為典型的“S”形曲線,這與前人研究提出的2種栓塞脆弱性曲線類型相符[28,29]。建立栓塞脆弱性曲線可以更好地了解植物栓塞抵抗力的特征和變化,并進一步研究樹木耐旱性和抵御干旱等環境壓力的能力[30,31]。扁桃砧木資源的木質部解剖結構與其栓塞抗性間存在一定的相關性。首先,普通桃和苦仁桃巴旦的直徑D較其它扁桃砧木資源大,且其與P50值相關性較高,D越大,易發生栓塞,栓塞抗性越小,與前人研究中大徑導管植物比小徑導管植物更容易發生栓塞的結論相似[32,33]。普通桃和苦仁桃巴旦之所以容易發生栓塞也可能與前人研究提出的氣種假說有關,導管的紋孔膜透性以及紋孔膜上微孔的大小,尤其是在干旱等脅迫條件下,對植物的耐旱性具有至關重要的影響[34,35]。隨著D的增加,紋孔占據的面積相應增大,紋孔膜上出現較大微孔的概率也隨之提高,降低了植物對栓塞的抵抗力。在變化的水分環境和生長條件下,樹木通過調整自身形態和結構來協調導水效率與栓塞抗性,從而確保其生長發育的成功,特別是在干旱或不穩定環境中尤為關鍵[36]。
除導管直徑外,栓塞抗性還與(TVW/D連接導管)2有關。有研究表明,(TVW/D連接導管)2與抗栓塞能力有一定的關系,(TVW/D連接導管)2較大的植物可以減小水勢負壓對木質部導管壁的破壞[37]。Cochard等[30]對薔薇科李屬植物研究得出相同結論,導管壁理論機械強度越大,P50值與其相關性越好。紋孔膜厚度和導管壁厚度較大的植物具有較高的木材密度和力學性質,這是因為較厚的導管壁可以增加木材的強度和抗壓能力,而紋孔膜的厚度則可以減少木材的孔隙率,從而增加木材的密度[14,27]。木巴熱克等[26]對扁桃引進品種和新疆本地品種的莖木質部解剖結構和栓塞抗性關系進行研究,發現其木材密度和栓塞抗性間有較強的相關性,木材密度越大,栓塞抗性越強,植物不易發生栓塞。在研究中,栓塞抗性較強的大巴旦和苦巴旦的WD和(TVW/D連接導管)2大,栓塞抗性強,可以更好的承受水勢負壓對其導管壁的破壞。"
3.2"栓塞修復與木質部解剖結構間的關系
植物木質部內部相鄰導管間水勢是否存在一定的差值是決定栓塞可否消除的前提,水勢差高時栓塞易修復[6]。鄰近部位或區域的水勢越高,栓塞修復越好。在修復過程中,莖木質部輸水結構尤其是紋孔膜結構特征,會影響所需水勢梯度的大小[11]。研究中,栓塞修復能力與導管直徑以及紋孔膜上微孔直徑呈正相關,導管直徑和微孔直徑小,木質部抗栓塞能力強,栓塞修復能力弱。
Ogasa等[8]的研究結果表明,木材密度和栓塞抗性與修復能力之間存在負相關關系。這表明,木材密度越高,植物的抗空穴化能力強,但栓塞的修復能力可能會降低。因此,木材密度低的植物可能會在栓塞修復方面表現更加出色。當受到干旱脅迫時,高密度的木材可能會為植物提供更好的保護,可以更好地防止空穴的發生。然而,當植物需要頻繁修復栓塞時,低密度的木材可能更有利于栓塞的修復。研究發現,栓塞修復能力與木材密度呈負相關,木材密度較小的普通桃,栓塞抗性較弱,但其栓塞修復能力強,以此適應長期干旱的環境。
3.3"栓塞抗性與栓塞修復間的關系
栓塞抗性較高的植物能夠有效地防止栓塞的發生,從而減少了栓塞修復的需求[6]。劉麗等[38]研究發現,84K楊(P.abla × P.glandulousa ‘84K’)的木質部導管直徑越大,其抗栓塞能力越弱,自然栓塞度越高,栓塞修復能力越強;黨維等[18]在6個樹種的1年生枝條中,木質部的水力特性與栓塞修復能力有著密切的聯系。尤其是栓塞脆弱性(P50)和水力直徑,是影響栓塞形成與修復的重要因素,與導管的結構和功能緊密相關;安鋒等[39]對8種木本植物木質部栓塞抗性、脆弱性和修復能力進行了測試和比較,結果表明,栓塞脆弱性越大,植物組織的破壞程度越大,反映出該植物對于環境的適應性較低,重新補水后,栓塞修復速率越快,意味著在干旱環境中具備更好的適應性和修復能力。在扁桃不同砧木資源中,普通桃的栓塞抗性最弱,P50最大,更容易發生栓塞;而大巴旦較耐旱,抗栓塞能力強,但栓塞后的修復能力弱。
4"結 論
扁桃5個砧木資源中,耐旱性較強大巴旦資源的莖木質部導管直徑以及微孔直徑小、木材密度大,栓塞抗性強,但栓塞修復能力弱;而缺水較敏感砧木普通桃的莖木質部導管直徑以及微孔直徑大、木材密度小,栓塞抗性弱,但栓塞修復能力強。
參考文獻"(References)
[1]"木巴熱克·阿尤普, 伊麗米努爾, 荊衛民.不同水分處理下幾種檉柳屬植物幼株木質部栓塞及其解剖結構特征[J].北京林業大學學報, 2017, 39(10): 42-52.Mubareke Ayoupu, Yiliminuer, JING Weimin.Xylem anatomical features and native xylem embolism of several Tamarix spp.species seedlings under different water treatments[J].Journal of Beijing Forestry University, 2017, 39(10): 42-52.
[2] McDowell N G, Beerling D J, Breshears D D, et al.The interdependence of mechanisms underlying climate-driven vegetation mortality[J].Trends in Ecology amp; Evolution, 2011, 26(10): 523-532.
[3] Nardini A, Battistuzzo M, Savi T.Shoot desiccation and hydraulic failure in temperate woody angiosperms during an extreme summer drought[J].New Phytologist, 2013, 200(2): 322-329.
[4] Domec J C, Gartner B L.Cavitation and water storage capacity in Bole xylem segments of mature and young Douglas-fir trees[J].Trees, 2001, 15(4): 204-214.
[5] Choat B, Jansen S, Brodribb T J, et al.Global convergence in the vulnerability of forests to drought[J].Nature, 2012, 491(7426): 752-755.
[6] 黨維.耐旱樹種木質部栓塞恢復狀況的研究[D].楊凌: 西北農林科技大學, 2016.DANG Wei.The Refilling of Embolized Xylem of Drought-tolerant Tree Species[D].Yangling: Northwest A amp; F University, 2016.
[7] Awad H, Barigah T, Badel E, et al.Poplar vulnerability to xylem cavitation acclimates to drier soil conditions[J].Physiologia Plantarum, 2010, 139(3): 280-288.
[8] Ogasa M, Miki N H, Murakami Y, et al.Recovery performance in xylem hydraulic conductivity is correlated with cavitation resistance for temperate deciduous tree species[J].Tree Physiology, 2013, 33(4): 335-344.
[9] Rolland V, Bergstrom D M, Lenné T, et al.Easy come, easy go: capillary forces enable rapid refilling of embolized primary xylem vessels[J].Plant Physiology, 2015, 168(4): 1636-1647.
[10] Love D M, Sperry J S.In situ embolism induction reveals vessel refilling in a natural aspen stand[J].Tree Physiology, 2018, 38(7): 1006-1015.
[11] 孟鳳.七種槭樹科植物木質部栓塞及其恢復與植物抗旱性的關系[D].楊凌: 西北農林科技大學, 2019.MENG Feng."The Relationship between Xylem Embolism and Embolism Recovery and Plant Drought Resistance in Seven Genus Acer[D].Yangling: Northwest A amp; F University, 2019.
[12] McCulloh K A, Meinzer F C.Further evidence that some plants can lose and regain hydraulic function daily[J].Tree Physiology, 2015, 35(7): 691-693.
[13] Hacke U G, Jansen S.Embolism resistance of three boreal conifer species varies with pit structure[J].The New Phytologist, 2009, 182(3): 675-686.
[14] Plavcová L, Hacke U G, Sperry J S.Linking irradiance-induced changes in pit membrane ultrastructure with xylem vulnerability to cavitation[J].Plant, Cell amp; Environment, 2011, 34(3): 501-513.
[15] Levionnois S, Kaack L, Heuret P, et al.Pit characters determine drought-induced embolism resistance of leaf xylem across 18 Neotropical tree species[J].Plant Physiology, 2022, 190(1): 371-386.
[16] Avila R T, Kane C N, Batz T A, et al.The relative area of vessels in xylem correlates with stem embolism resistance within and between Genera[J].Tree Physiology, 2023, 43(1): 75-87.
[17] Christensen-Dalsgaard K K, Tyree M T.Frost fatigue and spring recovery of xylem vessels in three diffuse-porous trees in situ[J].Plant, Cell amp; Environment, 2014, 37(5): 1074-1085.
[18] 黨維, 姜在民, 李榮, 等.6個樹種1年生枝木質部的水力特征及與栓塞修復能力的關系[J].林業科學, 2017, 53(3): 49-59.DANG Wei, JIANG Zaimin, LI Rong, et al.Relationship between hydraulic traits and refilling of embolism in the xylem of one-year-old twigs of six tree species[J].Scientia Silvae Sinicae, 2017, 53(3): 49-59.
[19] 李美琦, 姜在民, 趙涵, 等.加楊水力學與生理特性對不同土壤水分條件響應研究[J].植物生理學報, 2017, 53(4): 632-640.LI Meiqi, JIANG Zaimin, ZHAO Han, et al.Study on the adaptability of hydraulic and physiological characteristics to different soil moisture conditions in Populus x canadensis Moench[J].Plant Physiology Journal, 2017, 53(4): 632-640.
[20]梁艷霞, 張亞楠, 王占和.扁桃薄殼品種‘蒙特瑞’在山西中部地區的引種表現及栽培技術[J].果樹資源學報, 2022, 3(6): 88-90.LIANG Yanxia, ZHANG Yanan, WANG Zhanhe.Introduction performance and cultivation technology of the thin-shelled variety 'Monterey' of lentil in central Shanxi[J].Journal of Fruit Tree Resource, 2022, 3(6): 88-90.
[21] 侯江濤, 張毅芳, 沈聰聰, 等.扁桃引種栽培技術研究綜述[J].林業科技通訊, 2020,(2): 11-14.HOU Jiangtao, ZHANG Yifang, SHEN Congcong, et al.Review on introduction and cultivation techniques of Amygdalus communis[J].Forest Science and Technology, 2020,(2): 11-14.
[22] 梁艷霞, 王占和, 張亞楠, 等.扁桃特性及其豐產栽培技術[J].黑龍江糧食, 2021,(10): 108-109.LIANG Yanxia, WANG Zhanhe, ZHANG Yanan, et al.Characteristics of almond and its high-yield cultivation techniques[J].Heilongjiang Grain, 2021,(10): 108-109.
[23] 任哲, 貢翔, 張銳, 等.扁桃優株葉片解剖結構與其抗旱性關系研究[J].農業與技術, 2022, 42(3): 10-14.REN Zhe, GONG Xiang, ZHANG Rui, et al.扁桃優株葉片解剖結構與其抗旱性關系研究[J].Agriculture and Technology, 2022, 42(3): 10-14.
[24] Ayup M, Yang B, Gong P, et al.Evaluation of drought resistance of native almond-rootstock varieties in Xinjiang, China[J].New Zealand Journal of Crop and Horticultural Science, 2022, 50(1): 48-68.
[25] 李榮.耐旱樹種木質部結構與耐旱性關系研究[D].楊凌: 西北農林科技大學, 2016.LI Rong.Relationships between Xylem Strucrure And Drought Tolerance of Drought Tolerant Tree Species[D].Yangling: Northwest A amp; F University, 2016.
[26] 木巴熱克·阿尤普, 楊波, 艾沙江·買買提, 等.基于當年生枝木質部解剖結構的扁桃品種栓塞抗性分析[J].西北林學院學報, 2021, 36(5): 99-105.Rebareke Ayoupu, YANG Bo, Ashajiang Maimaiti, et al.Xylem embolism resistance of different AlmondCultivars based on the xylem anatomical characteristics of current-year shoot[J].Journal of Northwest Forestry University, 2021, 36(5): 99-105.
[27] Jansen S, Choat B, Pletsers A.Morphological variation of intervessel pit membranes and implications to xylem function in angiosperms[J].American Journal of Botany, 2009, 96(2): 409-419.
[28] Torres-Ruiz J M, Jansen S, Choat B, et al.Direct X-ray microtomography observation confirms the induction of embolism upon xylem cutting under tension[J].Plant Physiology, 2015, 167(1): 40-43.
[29] Sperry J S, Christman M A, Torres-Ruiz J M, et al.Vulnerability curves by centrifugation: is there an open vessel artefact, and are ‘r’ shaped curves necessarily invalid?[J].Plant, Cell amp; Environment, 2012, 35(3): 601-610.
[30] Cochard H, Barigah S T, Kleinhentz M, et al.Is xylem cavitation resistance a relevant criterion for screening drought resistance among Prunus species[J].Journal of Plant Physiology, 2008, 165(9): 976-982.
[31] Tyree M T, Engelbrecht B M J, Vargas G, et al.Desiccation tolerance of five tropical seedlings in Panama.relationship to a field assessment of drought performance[J].Plant Physiology, 2003, 132(3): 1439-1447.
[32] Cai J, Zhang S X, Tyree M T.A computational algorithm addressing how vessel length might depend on vessel diameter[J].Plant, Cell amp; Environment, 2010: 33(7): 1234-1238.
[33] Markesteijn L, Poorter L, Paz H, et al.Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits[J].Plant, Cell amp; Environment, 2011, 34(1): 137-148.
[34] Wheeler J K, Sperry J S, Hacke U G, et al.Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants: a basis for a safety versus efficiency trade-off in xylem transport[J].Plant, Cell amp; Environment, 2005, 28(6): 800-812.
[35] Cochard H, Bréda N, Granier A, et al.Vulnerability to air embolism of three European oak species (Quercus petraea (Matt) Liebl, Q pubescens Willd, Q robur L)[J].Annales Des Sciences Forestières, 1992, 49(3): 225-233.
[36] Johnson D M, McCulloh K A, Woodruff D R, et al.Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different[J].Plant Science, 2012, 195: 48-53.
[37] Hacke U G, Sperry J S, Pockman W T, et al.Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure[J].Oecologia, 2001, 126(4): 457-461.
[38] 劉麗, 張立, 蔡靖, 等.干旱脅迫及復水后84K楊栓塞修復及其他水力學特性的研究[J].北京林業大學學報, 2021, 43(7): 22-30.LIU Li, ZHANG Li, CAI Jing, et al.Hydraulic characteristics and embolism repair of Populus alba × P.glandulosa after drought stress and rehydration[J].Journal of Beijing Forestry University, 2021, 43(7): 22-30.
[39] 安鋒, 蔡靖, 姜在民, 等.八種木本植物木質部栓塞恢復特性及其與PV曲線水分參數的關系[J].西北農林科技大學學報(自然科學版), 2006, 34(1): 38-44.AN Feng, CAI Jing, JIANG Zaimin, et al.Refilling of embolism in the xylem of eight tree species and its relationship with Pressure-Volume parameters[J].Journal of Northwest Sci-Tech University of Agriculture and Forestry (Natural Science Edition), 2006, 34(1): 38-44.
Relationship between the anatomical structure of xylem"""and embolization characteristics of drought tolerant rootstocks of almond
YU Qiuhong1, XU Panyun2, GUO Chunmiao2, Dilixiati Hasimu1, Mubareke Ayoupu1
(1. College of Life Sciences, Xinjiang Agricultural University, Urumqi 830052, China; 2. Institute of Horticultural Crops, Xinjiang Academy of Agricultural Sciences, Urumqi 830091, China)
Abstract:【Objective】 ""To analyze the adaptation mechanism of drought resistant rootstocks of almond to the drought stress environment from the relationship between the stem xylem anatomical structure and the embolism characteristics.
【Methods】 ""The one-year old seedlings of five different almond rootstocks with different drought tolerance levels, including Da Badan (Amygdalus communes var.fragilis)(C1), Ku Badan ( A.communs var.amara)(C2), Tianren Taobadan [A.communs var."persistides (West.) Rehd.](C3), Kuren Taobadan [A.communs var.persistides (West.) Rehd.](C4), and Xinjiang local peach (A.persista L.)(C5), were selected as research materials.Using the \"external pressure\" method, combined with the \"XYLEM xylem hydraulic conductivity and embolism measurement system\" and \"PMS pressure chamber\", the degree of stem natural embolism level and embolism vulnerability value (P50) were measured under normal cultivation conditions.The embolism repairment of the embolized branches at different time periods (30, 80, 130, 180 min) and the xylem anatomical characteristics were analyzed to explore the relationship between the anatomical structure of the stem xylem, the embolism resistance, and the embolism repairment of different drought resistant almond rootstocks.
【Results】 ""(1) The vulnerability curves(CV) of Da Badan, TianRen Taobadan, Ku Badan, and local peaches were in an \"S\" shape, while the CV curves of Kuren Taobadan were in an \"R\" shape; (2) The order of stem xylem embolism resistance (P50) was Da Badangt; TianRen Taobadangt; Ku Badangt; Kuren Taobadangt;Local Peach, with Da Badan having the most embolism resistant xylem; (3) The order of xylem embolism repair ability was as follows: Local peachgt; TianRen Taobadangt; Kuren Taobadangt; Ku Badan gt; Da Badan; (4) The drought resistant rootstocks as Da Badan, Ku Badan , and TianRen Taobadan all had small vessels and micropores, high wood density, thick conduit walls and theoretical mechanical strength of conduit walls.They had relative strong embolism resistant ability, but were weaker to embolism repairment.Local peaches and Kuren Taobadan had larger vessels and micropores, lower wood density, and its vessel wall thickness, and theoretical mechanical strength of the vessel wall were smaller than other rootstocks.
【Conclusion】 ""In summary, the xylem embolism resistance is negatively correlated with embolism repair ability in different almond rootstocks, and the drought resistant rootstocks have embolism resistant xylem.The xylem embolism repairment is positively correlated with the diameter of vessel and micropores, and negatively correlated with wood density, vessel wall thickness, and theoretical mechanical strength of the vessel wall in xylem.
Key words:""almond; rootstocks; embolism resistance; embolism repairment; xylem anatomical structure
Fund projects:""Key research and development program of the Xinjiang Uygur Autonomous Region (2024B02018-1);Basic Scientific R amp;D Program of Public Welfare Research Institutions of Xinjiang Uygur Autonomous Region(KY2020110)
Correspondence author:"""Mubareke Ayoupu (1981-), female, from Shache,Xinjiang, associate researcher, doctorate, research direction: the physiological ecology of fruit tree adversity, (E-mail) mubarek@ms.xjb.ac.cn
收稿日期(Received):
2024-04-19
基金項目:
新疆維吾爾自治區重點研發計劃項目(2024B02018-1);新疆維吾爾自治區公益性科研院所基本科研業務經費項目(KY2020110)
作者簡介:
于秋紅(1998-),女,河北滄州人,碩士研究生,研究方向為植物逆境生理,(E-mail)2074276722@qq.com
通訊作者:
木巴熱克·阿尤普(1981-),女,新疆莎車人,副研究員,博士,研究方向為果樹逆境生理生態,(E-mail)mubarek@ms.xjb.ac.cn
