碳酸鹽工廠研究進展
2024-01-01 00:00:00古強邢鳳存文嬌劉子琪馮山山
沉積學報 2024年4期
關鍵詞:研究方法






關鍵詞 碳酸鹽工廠;分類方案;研究方法;主控因素
0 引言
碳酸鹽巖作為沉積巖主要組成成分之一,廣泛分布于全球多個年代地層中,數量多,面積廣。碳酸鹽工廠是指碳酸鹽生產的空間與過程[1?7]。在碳酸鹽沉積系統中,碳酸鹽顆粒組合與相關的碳酸鹽工廠控制了沉積地層結構和幾何形態,因此對地質記錄中碳酸鹽工廠的識別和演化過程解釋有利于研究目標區域的沉積、構造和海平面變化歷史的重建[8?10],并對理解碳酸鹽工廠運行機制至關重要[11]。近年來,碳酸鹽工廠研究在國內受到重視,大量學者對特定年代地層或特殊的碳酸鹽沉積物已經進行了碳酸鹽工廠相關的研究[12?17]。但這些研究多針對類型較為單一的碳酸鹽工廠,且聚焦于碳酸鹽臺地的沉積與演化,尚存在以下問題:(1)具有多種碳酸鹽工廠分類方案,卻無針對碳酸鹽工廠劃分方案及其發育特征的總結及歸納;(2)對碳酸鹽工廠研究方法的發展、完善及針對不同研究目的研究方法選取還存在盲目性;(3)碳酸鹽工廠消亡的主控因素復雜多樣且各因素間存在一定的相關性,對消亡主控因素的歸納及各因素控制碳酸鹽工廠的機制認識不足。因此,在國內外有關碳酸鹽工廠研究的基礎上,本文系統綜述了碳酸鹽工廠的劃分方案、研究方法與消亡主控因素的研究進展,以期為國內學者研究碳酸鹽工廠提供參考。
1 碳酸鹽工廠分類方案
碳酸鹽工廠在內部和空間上高度復雜[18],區分不同的碳酸鹽工廠是過去三十年來碳酸鹽沉積學取得的主要進展之一[7]。其劃分方案多種多樣,但大多依據碳酸鹽巖類型、主要沉積環境、沉積環境和沉淀方式、碳酸鹽工廠建造的主導者及海洋環境學條件(溫度、鹽度和營養物等)與海洋地理參數(大洋深度及地貌等)等進行碳酸鹽工廠類型的劃分。
1.1 依據碳酸鹽沉積物類型的劃分方案
Surlyk[19]在對歐洲西北部上白堊統—古新統丹麥階沉積的碳酸鹽巖研究中,依據沉積的主要碳酸鹽巖類型將碳酸鹽工廠劃分為三類并列出了三種碳酸鹽工廠的主要底棲生物及不同碳酸鹽工廠發育的相對位置。(1)生產生屑砂巖/粉砂巖的碳酸鹽工廠:主要沉積生屑砂巖及生屑粉砂巖,沉積物在近海處發育交錯層理,生屑砂巖分選良好,沉積在與海岸平行的水下沙丘區。(2)生產苔蘚蟲泥粒灰巖生物丘—小型苔蘚蟲粒泥灰巖生物丘的碳酸鹽工廠:在離岸更遠的地方,碎屑碳酸鹽巖逐漸演化為泥粒灰巖/粒泥灰巖沉積,沉積在透光帶下方相對較深的水域中,形成了顯著的生物丘。向海方向,生物丘逐漸變小,灰泥逐漸豐富,底棲生物化石逐漸減少。(3)生產深海灰泥的碳酸鹽工廠:深海灰泥代表第三個碳酸鹽巖工廠,幾乎沒有生物化石,僅可見少量球粒。
1.2 依據主要沉積環境的劃分方案
James[20]提出基于過程的,與氣候無關的兩個碳酸鹽巖工廠,Photozoan和Heterozoan,適用于整個顯生宙。(1)Photozoan(光養)碳酸鹽工廠:是指淺部、暖水、底棲鈣質生物群落環境中建造的碳酸鹽,強調了碳酸鹽沉淀所需要的能量來自光合作用。(2)Heterozoan(異養)碳酸鹽工廠:代表了Photozoan(光養)碳酸鹽工廠外其他環境的碳酸鹽的集合。但該種劃分方案將碳酸鹽工廠簡單劃分為兩類,因此需要更詳細的碳酸鹽工廠劃分方案。
1.3 依據沉積環境和沉淀方式的劃分方案
Schlager[1?2]在Lowenstam et al.[21]區分三種碳酸鹽沉淀基本模式(非生物成因、生物誘導成因、生物控制成因)的基礎上,依據沉積環境和沉淀方式將碳酸鹽工廠劃分為3類,但因為3種碳酸鹽工廠的產物是重疊的,因此工廠之間的界限在空間上是漸變的(圖1a)。后來Reijmer[4]提出增加冷水珊瑚礁作為第4類碳酸鹽工廠。大規模的鮞粒發育建造在其工廠體系中位置不明,因此Li et al.[14]將鮞粒工廠單列出來(圖1b),如今該種碳酸鹽工廠劃分方案運用最廣,受到廣大學者的認可。由于前幾種類型的碳酸鹽工廠均為底棲性質的碳酸鹽工廠,因此Schlager[22]又增加浮游碳酸鹽工廠作為第5類碳酸鹽工廠(圖1c)。
1.3.1 熱帶淺水工廠
熱帶淺水工廠分布于南北緯30°內的熱帶和亞熱帶、溫暖、營養貧乏、光照充足及富含氧氣的海洋表層,碳酸鹽沉淀很大程度上以生物控制和非生物為主。主要建造者為光自養綠藻、珊瑚、有孔蟲和某些軟體動物。溫度是其界限的主要控制因素,在目前的海洋中,熱帶碳酸鹽工廠的緯度界限指標為最冷月份平均氣溫約為20 ℃,而緯度只是其界限的一個粗略的替代指標。與其他碳酸鹽工廠相比,熱帶淺水工廠生產碳酸鹽的區域更淺更窄,但平均生產率最高[2]。熱帶淺水工廠可向冷水工廠過渡,例如在海洋的暖表層和溫躍層之間的邊界以及上升流將富含營養的冷水攜帶至熱帶淺水區域[23?25]。
1.3.2 冷水工廠
冷水工廠是深水和高緯度地區碳酸鹽的重要生產者,其范圍為從熱帶淺水工廠的邊界向兩極延伸,但在低緯度的溫躍層中也可能出現[23]。碳酸鹽沉淀多為生物控制型,異養生物占優勢,但光能自養生物的貢獻變化很大。碳酸鹽沉積物為砂粒大小的碳酸鹽生物骨骼碎片組成,缺乏熱帶型珊瑚礁和鮞粒,碳酸鹽灰泥很少,過渡到熱帶淺水工廠的區域通常延伸超過1 000 km[26]。在熱帶地區,如果環境惡劣,冷水工廠可以替代熱帶淺水工廠[1?2]。目前冷水碳酸鹽巖作為碳酸鹽的重要組成部分,隨著碳酸鹽工廠研究的深入,冷水碳酸鹽逐漸受到重視[20,27?30]。Wisshak et al.[31]對Azores冷水工廠不同水深、水動力和光分帶下的碳酸鹽生產、降解過程和主要生物種群進行了研究。
1.3.3 灰泥丘工廠
灰泥丘工廠與熱帶淺水工廠及冷水工廠有根本的不同,其主要產物是泥晶碳酸鹽,通常與微生物誘導和非生物沉淀有關。灰泥丘是在富含營養、低氧但不缺氧的水域沉積形成的,而非經沖刷形成的細小沉積物,從根本上不同于潟湖泥或深海等深流的水動力沉積物堆積。灰泥丘的主要成分泥晶灰巖在形成期間或形成不久后是固結的,而不是松散沉積物;泥晶灰巖是由無機和有機化學反應經過復雜的相互作用沉淀而成,其中微生物和腐爛的有機物起著核心作用。在重大生物危機之后,灰泥丘工廠的統治地位在短時間內重新確立[2]。在地質記錄中,淺水區和熱帶地區的灰泥丘工廠是不尋常的,它的出現通常是因為環境條件或大滅絕導致的缺乏更有競爭、更高效的碳酸鹽工廠[32?33]。
隨后增加的3種不同類型的碳酸鹽工廠中,冷水珊瑚工廠分布于全球范圍內,出現造礁珊瑚,并具有非常高的生物多樣性。很大程度上依賴營養物的穩定輸入,碳酸鹽產量低,生產范圍大,水深40~2 500 m。浮游碳酸鹽工廠通過浮游有孔蟲,顆石藻和其他浮游碳酸鹽生物在開闊海域產生碳酸鹽沉積物的工廠,也廣泛分布于全球范圍內。鮞粒碳酸鹽工廠概念的提出始于20世紀70年代,一般用來描述大規模鮞粒發育建造,其主要成分是海相碳酸鹽鮞粒。Li et al.[14]、李飛[34]通過對華南早三疊世溫室條件下的鮞粒工廠研究表明由于表層海水溫度和碳酸鹽飽和狀態之間的強線性關系,低緯度地區具有(季節性)干燥條件的極端溫室氣候顯著促進了當時鮞粒工廠的大規模碳酸鹽生產。
1.4 依據碳酸鹽工廠建造的主導者的劃分方案
Pomar et al.[8]強調多種物理和生態因素對碳酸鹽沉積過程的調節,注重碳酸鹽發育的影響因素研究。該方案將碳酸鹽工廠劃分為底棲自生泥晶工廠(圖2a)、淺水灰泥工廠(圖2b)、骨屑工廠初始期(圖2c)、厚殼蛤骨骼工廠繁盛期(圖2d)、底棲有孔蟲—藻類骨屑工廠(圖2e)和珊瑚—藻類,底棲—浮游共生骨屑工廠(圖2f)等,并可進一步細分為更多類型的碳酸鹽工廠。該方案并未對碳酸鹽工廠類型進行嚴格的定義,更多的是強調碳酸鹽工廠建造的主導者,且不同類型的碳酸鹽工廠可以共存于同相區或不同相區,并可發生相互轉化。
1.5 依據海洋環境學條件與海洋地理參數的劃分方案
Michel et al.[35]在綜述大量碳酸鹽巖臺地研究文獻的基礎上,使用一種側重于鑒別成分、地層結構和環境特征的確定性方法將淺水碳酸鹽巖產量與海洋學參數聯系起來。建立了基于海面海洋學參數(溫度、鹽度和初級生產力)的碳酸鹽工廠函數。該模型使用現代遙感和現場海洋學數據進行了測試,而古代海洋學模型的輸出則用白堊紀建模,從而確定5種碳酸鹽工廠(圖3)。
1.5.1 海洋生物化學工廠
對應于Pomar et al.[8]的淺水灰泥工廠以及一些淺水的微生物引起的碳酸鹽沉淀,如鮞粒和疊層石。這個工廠最典型的特征是寬達100 km的夾心蛋糕狀地層結構(layer-cake stratigraphic architecture)。發育非骨骼顆粒,受限制的海洋環流、相對中低的水動力及高的碳酸鹽飽和度。
1.5.2 光養生物T型工廠
典型特征是加積堆積狀的地層結構,對應于熱帶碳酸鹽陸架或分離的邊緣臺地,具有光養骨骼顆粒、生物礁及最高的碳酸鹽生產率。包括珊瑚、層孔蟲、藻類(特別是綠藻)、海綿和與光養藻類共生的底棲有孔蟲。局限于低緯度、溫暖貧營養及清澈的海洋環境。
1.5.3 光養生物C型工廠
較小的碳酸鹽堆積,大型底棲有孔蟲是沉積物成分的特征,多樣化的生物碎屑組成,通常位于溫暖帶或溫帶地區,具有明顯的季節性,初級生產力較低,光照是控制工廠布局的主要環境因素。相較于光養生物T型工廠對濁度和營養物質更具耐受性。
1.5.4 異養生物C型工廠
富營養化環境的異養生物群以及通常所說的冷水碳酸鹽巖。這個工廠的特點是異養生物群,如苔蘚蟲和軟體動物的聚集。位于營養豐富、富含有機物和浮游生物的海洋區域,如受河流徑流影響具有較強水動力條件的上升流區和沿海地區。
1.5.5 流體相關的工廠
同微生物工廠[22],通常建造非常陡峭的碳酸鹽巖沉積體,生產發生于大陸及深海環境。形成于缺氧條件下,通過降解海底大量有機生,有機物的含量和氧化條件是其主要驅動因素。細菌腐爛產生氨的過程或硫酸鹽還原導致海水的碳酸鹽飽和度局部增加。
2 碳酸鹽工廠研究方法物的微生物活動產
碳酸鹽巖研究方法多樣,綜合前人研究成果(表1),統計分析其針對碳酸鹽巖的研究方法。結果顯示,進行碳酸鹽巖研究使用的主要研究方法有:(1)巖相分析;(2)層序地層分析;(3)沉積相分析;(4)電子探針;(5)壓實及沉降速率計算;(6)激光測深和地形學;(7)碳氧同位素;(8)鍶同位素;(9)生物組合分析;(10)磷連續提取法;(11)牙形刺分析;(12)雙殼尺寸測量分析;(13)XRF分析;(14)掃描電鏡分析;(15)陰極發光分析;(16)拉曼光譜;(17)TOC分析;(18)地震分析;(19)Fe形態分析;(20)黃鐵礦分析;(21)天文旋回分析等。以上方法可總結為巖相、沉積相、層序、生物、組成成分、地球化學、特殊礦物及天文旋回等8類分析方法。
針對碳酸鹽工廠的研究中,采用的技術手段同樣源自碳酸鹽巖的各種研究方法。但早期研究受研究技術與設備條件的限制,前人主要利用碳酸鹽工廠所處的地理位置、野外宏觀巖性、結構、構造及宏觀古生物信息并結合室內偏光顯微鏡下對薄片進行巖性的鑒定、生物種類、組合及含量的定量分析,從而劃分出不同的微相組合來進行碳酸鹽工廠類型的劃分。
在碳酸鹽工廠發育特征及類型劃分的基礎上,通過層序地層分析、沉積相分析及地震分析等方法,并結合研究區地質背景、海平面升降變化與構造沉降速率等可用于判斷碳酸鹽工廠的消亡是否受構造演化及相對海平面的變化控制。但營養條件、氣候變化、氧化還原條件及鹽度等碳酸鹽工廠消亡的其他主控因素則需要結合地球化學分析等技術手段來進行綜合判斷。
隨著分析技術的發展,碳酸鹽工廠的研究方法逐漸精細,更定量化。激光測深和地形學方法被運用于現代多個碳酸鹽工廠的研究中。目前,針對碳酸鹽工廠的研究僅達到碳酸鹽巖組成成分及生物種類的定性研究是不夠的,進行野外精細取樣,室內光學顯微鏡下生物種屬及含量的定量分析,非生物組分顆粒的粒度分析是必要的。從而使研究結果能達到更準確,更高分辨率地劃分不同類型的碳酸鹽工廠,并以此研究不同碳酸鹽工廠間的轉換特征及機制。
連續的多類型地球化學測試分析、微觀Fe形態及黃鐵礦分析的引入,利用數學分析方法對沉積速率及天文旋回等方面的定量化研究,且天文旋回與碳酸鹽工廠潛在關系的研究中采用GR、δ13C及剩余磁等多種參數結果的綜合分析等,使得碳酸鹽工廠演化及主控因素的研究更深入且更精確。
3 碳酸鹽工廠消亡主控因素
碳酸鹽工廠經過不同時間的碳酸鹽沉積后,通過長期暴露、硅質碎屑掩埋或淹沒而使碳酸鹽巖沉積終止。Schlager[26]將碳酸鹽臺地消亡定義為“海平面相對上升超過碳酸鹽沉積,從而使得臺地/生物礁淹沒在碳酸鹽主要沉積的透光層之下”。前人將主控因素歸納為生態因素、氣候/環境因素及構造因素[13,68?69]。作者通過對國內外碳酸鹽臺地/碳酸鹽工廠建造及消亡最新研究文獻的調研(表1)(表中僅選擇性展示2013后發表的較新碳酸鹽工廠研究文獻),將主控因素分為11種,且各主控因素間存在相關性。
3.1 營養水平
全新世珊瑚礁上的珊瑚生長率表明,碳酸鹽臺地能夠很容易跟上地層長期下沉和海平面變化的步伐,但淹沒的珊瑚礁和碳酸鹽臺地卻在地質記錄中很常見,這一事實構成了一個科學悖論[26]。Margalef[70]發現高營養環境中造礁石珊瑚稀少。隨后Schlager[26]提出相較于海平面的上升,通過環境壓力降低碳酸鹽臺地生長潛力可能是臺地淹沒的方式。Hallock et al.[71]指出營養物質對珊瑚礁群落起負面影響為這一悖論提供了解釋。珊瑚礁群落的主要碳酸鹽沉積物生產者高度適應了營養缺乏的環境,硝酸鹽和磷酸鹽的輸入刺激了浮游生物的生長[45],降低了海水的透光度[48,72?74],限制了蟲黃藻—珊瑚和鈣藻等造礁生物的生存深度范圍[74?78](圖4)且磷酸鹽抑制了碳酸鈣晶體的形成[79?81],降低了碳酸鹽臺地的生長潛力[82]。此外,碳酸鹽生產和生物侵蝕的速率是相似的[83?85],較高的營養濃度和浮游生物密度也刺激了底棲生物中肉質藻類和非繁殖型懸食生物的生長[86?87],改變了底棲生物的群落結構[88]。除了取代造礁石藻類和珊瑚外,許多快速生長的競爭對手也是主動破壞珊瑚礁結構的生物腐蝕劑,因此即使養分的適度增加也能使珊瑚礁群落碳酸鹽巖生產力從凈生產轉變為凈侵蝕。
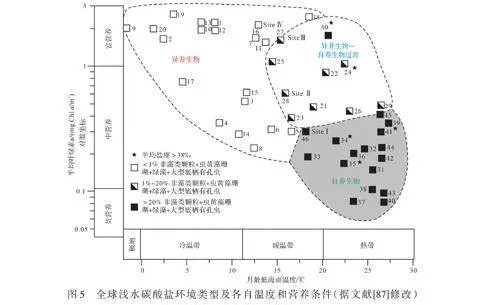
δ13Corg和δ15N反映了富含營養物質的表層水使初級生產力增加和洪水泛濫導致的有機物掩埋增加[88],將更高的有機物通量帶入海水,導致生產碳酸鹽的底棲生物死亡,并促進富含有機物的泥巖對碳酸鹽沉積物進行覆蓋[89]。地質記錄中,未沉積、生物侵蝕及氧化還原電位的降低等證據表明被淹沒的珊瑚礁與碳酸鹽臺地有過量的營養物質。營養過剩導致碳酸鹽臺地淹沒的其他機制包括局部或區域性上升流模式及大洋對流的變化[90]。Halfar et al.[91]通過統計分析全球現代碳酸鹽數據,建立了根據現有的生物組合區分高營養水平下和低營養水平下形成的碳酸鹽系統的標準。并且將全球范圍內營養和溫度條件與現代碳酸鹽組成進行比較,揭示了隨著營養增加和溫度降低,光自養碳酸鹽生產者逐漸減少[87](圖5)。Colettiet al.[46]利用磷連續提取法證明了中新世意大利北部及法國南部碳酸鹽工廠深受營養物質的影響。且當研究層位沒有明顯的陸源物質時,海水中溶解的Al3+可能被吸附到顆粒物質上形成包殼顆粒的氫氧化物[92]而使Al不適合于標準化。因此可選擇Ti作為碎屑元素,利用Ln(Cu/Ti)和Ln(Ba/Ti)作為海洋營養條件的代用指標,用于推斷營養物流入量和生產力水平[54]。
3.2 缺氧事件
前人研究表明幾期重大的碳酸鹽臺地消亡事件都發生于海洋缺氧期間[50,93?94],這種關系對于泥盆紀[26,67]及中白堊世[26,61?62,95?100]珊瑚礁和臺地的廣泛消亡尤為明顯。北特提斯白堊系碳酸鹽臺地4次淹沒都與缺氧事件具有較好的相關性,證實了缺氧事件與碳酸鹽工廠消亡具有因果關系[101?102]。南阿爾卑斯山Trento臺地在早侏羅世缺氧事件期間消亡,隨后沉積了深水碳質泥灰巖[103];古近紀—新近紀的碳酸鹽臺地消亡事件主要集中在始新世和中新世[26],當時全球海洋同樣具有明顯的缺氧趨勢[104]。
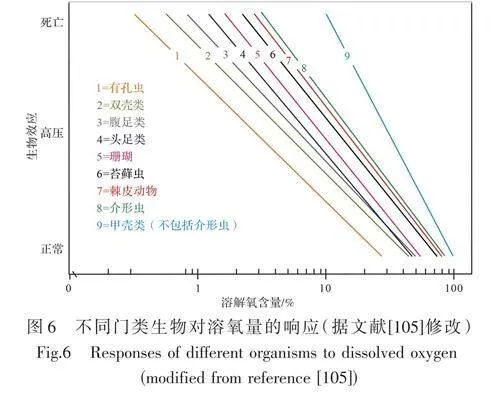
Arthur et al.[96]觀察到“快速的海侵可能將最小含氧層帶到整個淺水碳酸鹽巖區域”。但最小含氧層必須位于透光層之下,所以這種情況需要超過100~200 m的快速海侵,這很難達到,除非同時有某種機制將營養物混入表層水,從而造成浮游植物的大量繁殖并降低透光層的深度。因為大洋對流會將營養物質帶入表層水,所以大洋對流為此提供了可能。Hallock et al.[71]認為在缺氧事件期間存在海洋分層的條件下,大洋對流是可能的,并且碳酸鹽臺地的消亡可能是被大洋對流帶到地表的營養物所造成的。海洋缺氧事件對生物有著重要的影響,不同的海水含氧量將直接影響生物的生存,從而導致碳酸鹽工廠的消亡[105?106](圖6)。鉬是一種過渡金屬元素,通常富集在缺氧條件下沉積的富含有機物的沉積物中,富集程度反映了局部/全球鉬的總體可用性以及有機碳的濃度。因此,前人利用鉬在同期黑色頁巖中的富集說明了缺氧的短暫擴散[107?108]。
3.3 構造運動
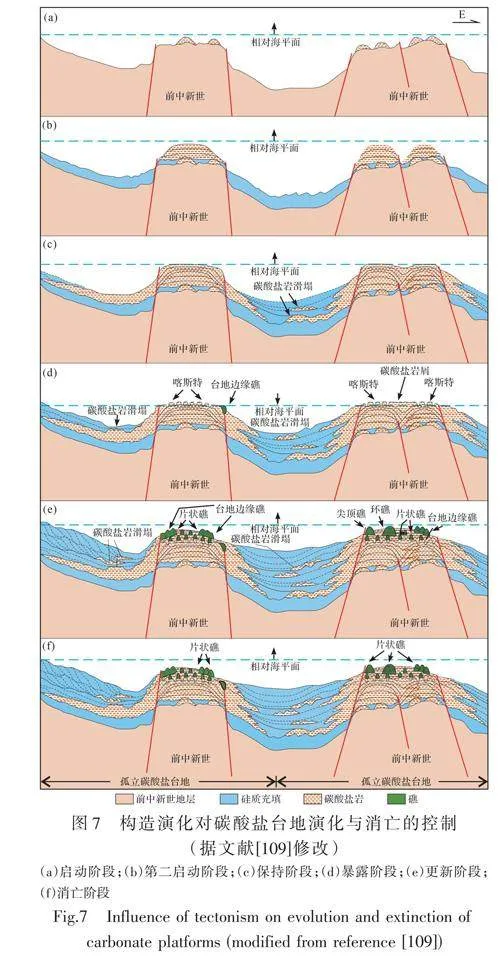
構造運動導致的碳酸鹽臺地隆起/下沉可能導致碳酸鹽巖平臺的消亡[37,40,43,48,58,88,90,109](圖7),且構造運動引起的大洋對流導致營養水平上升也為碳酸鹽臺地的消亡提供了一種可能機制。
Taira[110]引用了FairBridge的“由板塊運動觸發的海洋盆地突然運動將導致海洋中的大洋對流”假設,并通過研究證實現代相對較低水平的構造運動的確能夠影響現代海洋的熱分層,從而導致大洋對流。因此在海水分層不穩定的時期,構造運動觸發的大洋對流與快速海侵同時發生時會使碳酸鹽臺地消亡。Vogt[111]研究表明火山活動導致的缺氧、富營養的上升流是碳酸鹽巖工廠消亡的可能因素,并對不同成因上升流(圖8a)造成碳酸鹽工廠消亡的可能機制進行了分析(圖8b)。Bahamonde et al.[112]對西班牙西北部晚石炭世碳酸鹽臺地的建造和消亡研究認為區域性構造運動引起的海平面重大變化是臺地消亡的主要控制因素。Bekker et al.[93]研究認為美國古元古代碳酸鹽臺地兩次消亡事件很可能與Kenorland斷裂有關的成熟被動邊緣的解體有關。劉采等[36]認為鄂爾多斯南緣碳酸鹽臺地淹沒的區域構造背景應與秦嶺造山帶構造演化密切相關。意大利Apennines下侏羅統碳酸鹽臺地淹沒的分析表明,構造運動導致快速沉降促使可容納空間的增加控制了碳酸鹽工廠的消亡,而非營養資源的變化或古海洋學的變化導致[38]。Alborz Basin盆地下石炭統邊緣差異斷塊機制導致盆地西部的抬升及東部的沉降,從而使西部地層頂部保存大量的水下暴露和角礫巖化[44]。Jin etal.[53]對四川漢旺卡尼期碳酸鹽臺地消亡的研究表明該地區的碳酸鹽臺地危機和強烈的陸源輸入并非由卡尼期洪水事件的開始導致,而可能是印支造山期間與前陸盆地形成有關的沉降速率加快、環境變遷和增強的陸源碎屑輸入造成。
同沉積構造活動和火山作用是控制裂谷盆地碳酸鹽基底位置、大小和形狀的兩個主要因素。碳酸鹽臺地被大量分割為斷塊,這可能是由于噴發的火山碎屑輸入隔離碳酸鹽工廠生產地區導致[88,113?115],álvaro et al.[116]通過對摩洛哥沿海寒武紀碳酸鹽工廠的研究證實了碳酸鹽工廠的最終消亡與該地區的構造活動有關。火山活動對環境和生態造成破壞從而影響碳酸鹽巖的沉積以及火山碎屑大量輸入,直接替代碳酸鹽沉積[56]。奧陶紀華南板塊運動引起的火山活動導致碳酸鹽臺地陷落形成深水斜坡[117]。以上大量研究證實構造運動直接或間接導致了碳酸鹽工廠的消亡。
3.4 陸源物質輸入
陸源物質的輸入可劃分為兩種。(1)陸地徑流中營養物質輸入[37,45,61,71,82,90,118],導致這些營養物質在硅質陸源碎屑沉積物到達之前抑制了碳酸鹽工廠的生產;如果在下沉超過臨界深度之前未能將多余的營養物質從系統中排出,碳酸鹽工廠就會消亡。(2)陸源硅質碎屑加速進入近海盆地,導致大陸邊緣從以碳酸鹽為主的沉積轉變為硅質碎屑沉積[47?49,119]。
3.5 生物大滅絕事件
碳酸鹽生產受到生物組合的影響,生物大滅絕導致碳酸鹽生產者減少從而控制了碳酸鹽工廠的消亡[5,50,120?121],生物大滅絕事件常伴隨著碳酸鹽產量的下降(圖9)。
Philip et al.[122]通過對位于西地中海白堊紀塞諾曼階—土倫階的碳酸鹽臺地進行研究發現,其消亡開始于晚白堊世和Helvetica生物帶時期。重大生物更替影響了底棲生物,使其發生了一次嚴重的滅絕事件,導致主要分泌文石的生物消失。Iba et al.[123]研究發現太平洋西北部白堊系碳酸鹽臺地生物群的滅亡很可能不是一個局部事件,而是白堊紀海洋中具有全球影響的事件的一部分,從而導致了碳酸鹽臺地的消亡。在環境變化和生物危機期間,微生物是碳酸鹽系統的重要貢獻者[124?126],并在高營養水平下發育良好[127?130]。
3.6 海平面變化
普遍認為相對海平面變化是無法超過碳酸鹽臺地沉積的[26],因此海平面對碳酸鹽臺地消亡的影響可能為大規模的洪水事件、快速海侵和長期的海洋學變化導致洪水期海底生態系統壓力增加[64,112,131?132]。海平面快速上升淹沒了以前的陸地,導致泛濫平原土壤中的營養物質混入,抑制了珊瑚礁/碳酸鹽臺地的再生長也可能是碳酸鹽工廠消亡的因素[47,49,90,130]。Jarvis et al.[133]研究發現白堊紀塞諾曼階—土倫階的黏土含量異常高,這也可能與海平面的快速上升有關;Thierry et al.[134]以及Wilmsen[82]認為疊加在二級海平面上升之上的快速和高幅度的三級海平面波動可能對碳酸鹽臺地的淹沒起了很重要的作用。但海平面的下降也可能是粗粒陸源硅質碎屑巖進積的原因,并且導致碳酸鹽臺地暴露從而破壞碳酸鹽工廠的生產能力[61,86]。Sattler et al.[135]研究南海中新世珠江組碳酸鹽工廠消亡的主要原因之一是相對海平面的上升。除海平面上升之外,海退事件可導致沖刷重力河道沉積、侵蝕面、沉積間斷的形成,且全球海退的跡象可能反映了冰川作用的強烈影響[44]。海平面升降變化導致的碳酸鹽工廠消亡常伴隨著暴露剝蝕,因此可根據碳氧同位素數據及巖相學證據來解釋碳酸鹽巖內部的重復陸上暴露[135]。不整合面以下碳氧同位素值的降低通常反映了地表暴露以及土壤帶CO2和來自大氣降水的低氧同位素[136?138]。對不連續面下同位素降低的另一種解釋為來自沿流動通道運移的埋藏流體的膠結物沉淀[139],與正常的泥晶灰巖相比,埋藏泥晶灰巖的氧同位素值通常較低,因為埋藏流體的溫度較高[140]。此外,低碳同位素信號可能由地下環境中高溫下的有機物降解產生[141]。除此之外還可利用多種手段對海平面變化進行研究,從而確定海平面變化是否為所研究地層碳酸鹽工廠消亡的主控因素。
3.7 氣候變化
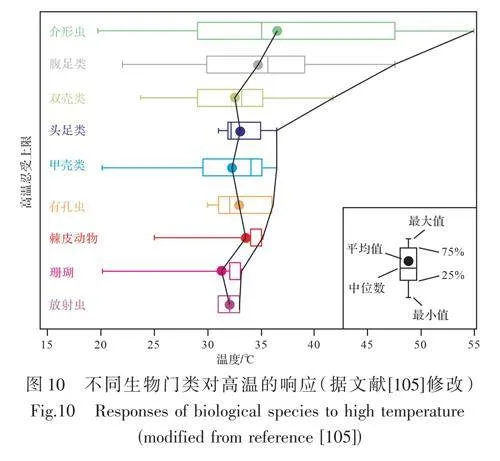
熱帶和非熱帶碳酸鹽臺地在幾何形狀、沉積相帶特征、地層結構、沉積物堆積速度和碳酸鹽生產生物群方面存在差異[142?143]。氣候決定了溫度、鹽度和養分的有效性,并在很大程度上控制著形成碳酸鹽的生物組成[37,48,52,65?66,87]。Simms et al.[144]研究假設一個“洪水事件”打破了晚三疊世普遍的干旱氣候,導致粗粒、成熟度低的硅質碎屑徑流和堆積增加,表現為黑色頁巖(或綠色頁巖、放射蟲硅質巖)覆蓋于特提斯范圍的碳酸鹽巖之上,從而導致碳酸鹽臺地的消亡。這一假設在全球多個地方被證實,稱為“卡尼期洪泛事件[49,55,60,119,145?150]。Betzler et al.[151]研究發現表層水溫抑制了昆士蘭碳酸鹽臺地生物礁的發育;Simone etal.[152]也討論了溫度對碳酸鹽臺地消亡影響的例子。較大的底棲有孔蟲繁殖的最低溫度是17 ℃~20 ℃,如此低的溫度無法支持珊瑚礁的生長[153?156],當表層水溫度再次達到足以支撐珊瑚礁成長時,其可能已經下沉到透光層以下,或者下沉到不能形成生物礁的深度。且碳酸鹽巖工廠的消亡可能會加劇海水溫度下降的速率[157?158]。西班牙白堊系塞諾曼—土倫階Altamira碳酸鹽臺地消亡研究表明臺地消亡伴隨著不斷加劇的溫室氣候,增加了河流從大陸向海洋的轉移,導致了初級生產力的提高和隨之而來的有機物埋藏的增加[81]。Sattler et al.[135]研究南海中新世珠江組碳酸鹽工廠消亡的主要原因之一是向淹沒不整合面海水溫度逐漸下降。極端的高溫氣候同樣會對海洋生物造成致命的打擊,每種生物都有一定的高溫忍受上限,常見生物多為30 ℃~40 ℃,僅個別生物能超過40 ℃。因此當高溫氣候產生時,生物就會出現死亡乃至滅絕[105?106](圖10)。同時季風活躍導致洋流增強同樣是碳酸鹽工廠消亡的原因之一[63]。
3.8 天文旋回
Martinez et al.[159]在綜合了西歐盆地侏羅紀時期保存完好的箭石中測得的δ13C值的演化后,確定了碳酸鹽產量的變化與穩定的δ13C值相關,因為長期的δ13C值最大值與低碳酸鹽生產同步,而長期δ13C最小值對應于高碳酸鹽產量的時期,并將這些長期的δ13C值變化解釋為9.1 My偏心率軌道周期的結果。且對不同偏心率進行了分析。在高偏心率時期,盡管季風事件加強了營養物質、有機碳和碳酸根離子向海洋的輸入,但因為干燥氣候和海水中的有效氧化條件[159],有機碳的掩埋被阻止。這導致了δ12C在海水中的富集及海洋碳酸鹽沉積物中δ13C值隨之降低。低偏心率間隔導致潮濕條件,這促進了高風化率、養分輸入、生產力水平和海洋中的有機碳埋藏,并導致了δ13C值的增加[157](圖11)。除此之外,高頻旋回受天文軌道影響的海平面變化控制,因此在高頻層序劃分的基礎上研究軌道周期對碳酸鹽巖工廠的控制同樣具有重要意義[41,160]。
3.9 海水酸堿度
大氣中CO2濃度上升時,海氣交換導致CO2混入海洋,因此DIC(溶解無機碳)增加,但海水中和酸的能力不變,導致碳酸根離子濃度與海水碳酸鹽飽和度降低,從而文石/方解石不沉淀[108,161]。碳酸根離子濃度的降低對鈣化生物非常重要,因為它不僅與PH值的降低有關,還降低了鈣離子的飽和狀態,從而影響了海洋中碳酸鈣礦物的穩定性。海洋酸化被用來解釋過去世界上多個地方深水沉積物中CaCO3含量快速且顯著下降的地質現象,表明了溶躍層(ACD)和補償深度(CCD)的大幅上升,海洋酸化導致碳酸鹽溶解、高度鈣化的生物化石的豐度急劇下降以及一些物種尺寸的減小[162],從而導致碳酸鹽工廠消亡[51,163?175]。火山活動停止后,大氣中CO2過量會由于高速率的硅酸鹽風化和碳酸鹽埋藏而在幾萬—幾十萬年后恢復到原來的狀態[176?178]。一旦海洋堿度恢復,由于較高的碳酸鹽飽和度,化學沉淀就替代了大部分由生物主導的碳酸鹽工廠。因此生物鈣化危機后海水碳酸鹽過飽和可能是短期內鮞粒廣泛出現的控制因素[179]。例如:二疊紀—三疊紀危機,記錄了碳酸鹽工廠從生物碳酸鹽巖到鮞粒碳酸鹽巖的突然轉變伴隨著生物危機、大的碳循環擾動,同時過高的碳酸鹽飽和度可能導致了巨鮞的出現[34,108,180?182]。
3.10 海水鹽度
鹽度是控制碳酸鹽生產環境的主要因素[183]。Allison et al.[184]研究發現鹽度差異影響著碳酸鹽工廠的產量,且鹽度降低的楔狀體在靠近陸地的克拉通環境中可能很常見(圖12),鹽度的降低會抑制顆石藻的鈣化[185]。但在超鹽度海水中文石是不飽和的,文石質骨骼被廣泛溶解及交代[186?187]。地中海墨西拿鹽度危機因在短時間間隔內形成了巨厚的巖鹽和其他蒸發巖礦物被描述為一場生態危機[188?193]。前人對其研究發現高鹽度環境是大型珊瑚礁堆積及半深海動物群消失的原因[194?196]。封閉海水環境中受強烈的蒸發作用,鹽度逐漸升高,形成碳酸鹽—碳酸鹽/蒸發巖—蒸發巖沉積的演化體系[197?198],且碳酸鹽產量受蒸發巖變形和溶解的局部影響[199?200]。隨后贊克爾期洪水使海平面上升,鹽度降低,環境壓力變小后,出現兩種微生物主導的碳酸鹽臺地沉積[201?202]。
3.11 碳酸鹽沉積速率/碳酸鹽臺地生長潛力
海洋中大多數碳酸鹽的沉淀都被限制在透光層(大約海洋上層100 m)以上,當碳酸鹽臺地被淹沒到碳酸鹽巖生產幾乎停止的海底深度時,就會發生碳酸鹽工廠的消亡。因此淺水碳酸鹽巖沉積速率可能決定一個礁/碳酸鹽臺地的生死[1]。Schlager[203]通過避免了比率相關性問題[204]影響的沉降速率分析實驗證實,碳酸鹽沉積速率隨著時間的增加而降低,其變化大約與時間變化的平方根成反比,碳酸鹽沉積速率決定著生物礁/碳酸鹽臺地的生長潛力。碳酸鹽臺地生長潛力的降低主要是由于環境因素,而不僅僅是由于陸上暴露時間的增加。這一假設得到了以下事實的證實:碳酸鹽臺地消亡和向陸地退積可能會持續數千萬年,并與長期的地球動力學過程相聯系。同樣,碳酸鹽臺地生物在數百萬—數千萬年內未能適應海平面,表明長期過程帶來了壓力。
4 結語
前人為碳酸鹽工廠的研究奠定了堅實的基礎,已對多個年代地層或特殊的碳酸鹽工廠進行過研究。筆者在前人研究的基礎上,對碳酸鹽工廠分類方案、研究方法及消亡主控因進行了系統的整理和分析。將碳酸鹽工廠歸納為依據碳酸鹽巖類型、主要沉積環境、碳酸鹽工廠建造的主導者、沉積環境和沉淀方式及海洋環境學條件與海洋地理參數進行類型劃分的5種分類方案,并介紹了不同劃分方案所劃分的碳酸鹽工廠類型及特征。歸納了碳酸鹽工廠的研究方法,并將消亡的主控因素總結為營養水平、缺氧事件、構造運動、陸源物質輸入、生物大滅絕事件、海平面變化、氣候變化、天文旋回、海水酸堿度、海水鹽度及碳酸鹽沉積速率/碳酸鹽臺地生長潛力,并分析了每種主控因素導致碳酸鹽工廠消亡的原因及過程。碳酸鹽工廠特征多樣,類型復雜且隨著環境變化可發生相互轉化。在碳酸鹽研究日趨高精度及高分辨率的要求下,學者需要依據研究目的有針對性地選擇碳酸鹽工廠分類方案及研究方法。進行系統、密集的取樣分析,綜合多因素進行碳酸鹽工廠類型的劃分,從而達到碳酸鹽工廠演化的高分辨率識別。以及運用多學科知識方法進行碳酸鹽工廠研究,從多方面認識其運行機制、演化過程及主控因素,使研究結論更準確,并發掘其中蘊藏的生物學及海洋學意義,可能是未來碳酸鹽工廠研究的發展方向。
猜你喜歡
教育界·下旬(2016年8期)2016-12-14 10:03:43
戲劇之家(2016年21期)2016-11-23 20:07:49
儷人·教師版(2016年14期)2016-11-22 01:13:04
商情(2016年39期)2016-11-21 09:22:07
中國集體經濟(2016年27期)2016-11-19 12:49:59
財經界·學術版(2016年19期)2016-11-16 15:41:54
大經貿(2016年9期)2016-11-16 15:38:49
化學教與學(2016年10期)2016-11-16 13:12:52
人間(2016年28期)2016-11-10 21:39:41
人間(2016年28期)2016-11-10 21:23:59
