MAPKKK參與植物生長發育及逆境脅迫調控的研究進展
2023-12-25 07:38:46陳磊清陳麗妃
福建農業科技 2023年9期
陳磊清,陳麗妃
(福建師范大學生命科學學院,福建 福州 350117)
促分裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPK或MPK)是一種廣泛存在于真核生物中,在進化過程中高度保守的生物信號轉導模塊,對植物生長發育及抵抗逆境脅迫起著至關重要的作用。典型的MAPK級聯信號通路由MAPK激酶(Mitogen-activated protein kinasekinasekinase,MAP3K或MAPKKK或MKKK)、MAPK激酶(Mitogen-activated protein kinasekinase,MAP2K或MAPKK或MKK)和MAPK組成,該信號傳導途徑可將植物感受到的環境信號放大并傳遞,從而激活下游的酶及轉錄因子,最終激活靶基因表達,對外界信號做出應答[1]。MAPKKK位于MAPK級聯通路的上游,可磷酸化激活MAPKK T環Thr/Ser基序中的兩個氨基酸,進一步磷酸化激活MAPK(圖1)。MAPKKK基因家族成員通常數量龐大,且在植物的生長發育及適應逆境脅迫方面發揮極其重要的作用。盡管MAPK級聯通路中部分MAPKKK成員的功能已被闡明,但大多數MAPKKK成員的功能仍然未知。MAPKKK在真核生物中具有普遍性和高度保守性,探索MAPKKK在植物調節生長發育以及響應外界生物或非生物脅迫信號的分子機制,發掘MAPKKK逆境脅迫調控應答功能基因并解析其分子調控通路,對提高作物抗逆性和培育具優良抗性的作物品種具有重要的理論指導意義。基于此,本文主要對MAPKKK參與調節植物生長發育及響應逆境脅迫的研究進展進行綜述,期望為MAPKKK的分子機制研究提供參考。

圖1 植物MAPKKK介導的MAPK級聯途徑作用模式圖
1 植物MAPKKK的組成及分類
MAPKKK作用于MAPK級聯通路的頂端,是一類蘇氨酸 (nr)/絲氨酸 (ser)激酶,能通過磷酸化中游MAPKK活化環S/T-XXXXX-S/T基序中S/T殘基來激活MAPKK,進而磷酸化激活下游MAPK。1993年Banno等[2]從煙草中分離出來的第一個編碼MAP3K的基因NPK1,隨后越來越多的MAPKKK基因在植物中被鑒定出來[3]。與中游MAPKK和下游MAPK相比,MAPKKK數量最為龐大,也表現出更明顯的復雜性和序列多樣性,擁有更為多樣的初級結構和二級結構。根據催化結構域的氨基酸保守基因序列的不同,MAPKKK基因家族可細分為MEKK、Raf和ZIK這3個亞家族,MEKK和Raf是其中兩個較大的亞組,ZIK亞家族成員最少[4],大多數植物都符合這一規律(表1)。MEKK亞族成員中YDA、ANP1、ANP2、和ANP3與細胞周期調控相關,Raf亞族成員中CTR1和EDR1與逆境脅迫相關。每類成員含有必需的蛋白激酶區域以及其他的功能特區,MEKK大多數具有保守的G(T/S)Px(W/Y/F)MAPEV結構域,Raf大多數具有保守的GTxx(W/Y)MAPE結構域,ZIK大多數具有保守的GTPEFMAPE(L/V)Y結構域[5]。值得注意的是,MEKK亞族的蛋白質結構保守性與ZIK亞族和Raf亞族相比相對較低,這也意味著MEKK亞族在植物生長發育及逆境響應的功能更為重要。

表1 26種植物的MAPKKK的分類
2 MAPKKK在植物生長發育中的調控應答
植物的生長發育受多種因素調控,是一個復雜且重要的生理過程。據大量文獻報道,MAPK級聯信號通路與植物生長周期的基本發育過程相關,幾乎參與植物生長的整個過程,包括配子和胚胎發育、形成組織器官及形態建成等,而且在植物生長發育中細胞增殖和細胞分化兩個重要的進程都起著關鍵的作用,是植物生長發育中不可或缺的信號傳導和調控途徑之一。MAPKKK作為MAPK級聯通路中上游成員,在植物胚胎發育、氣孔發育、根部發育、葉片和花結構發育等過程中都起著不可或缺的調節作用(表2)。
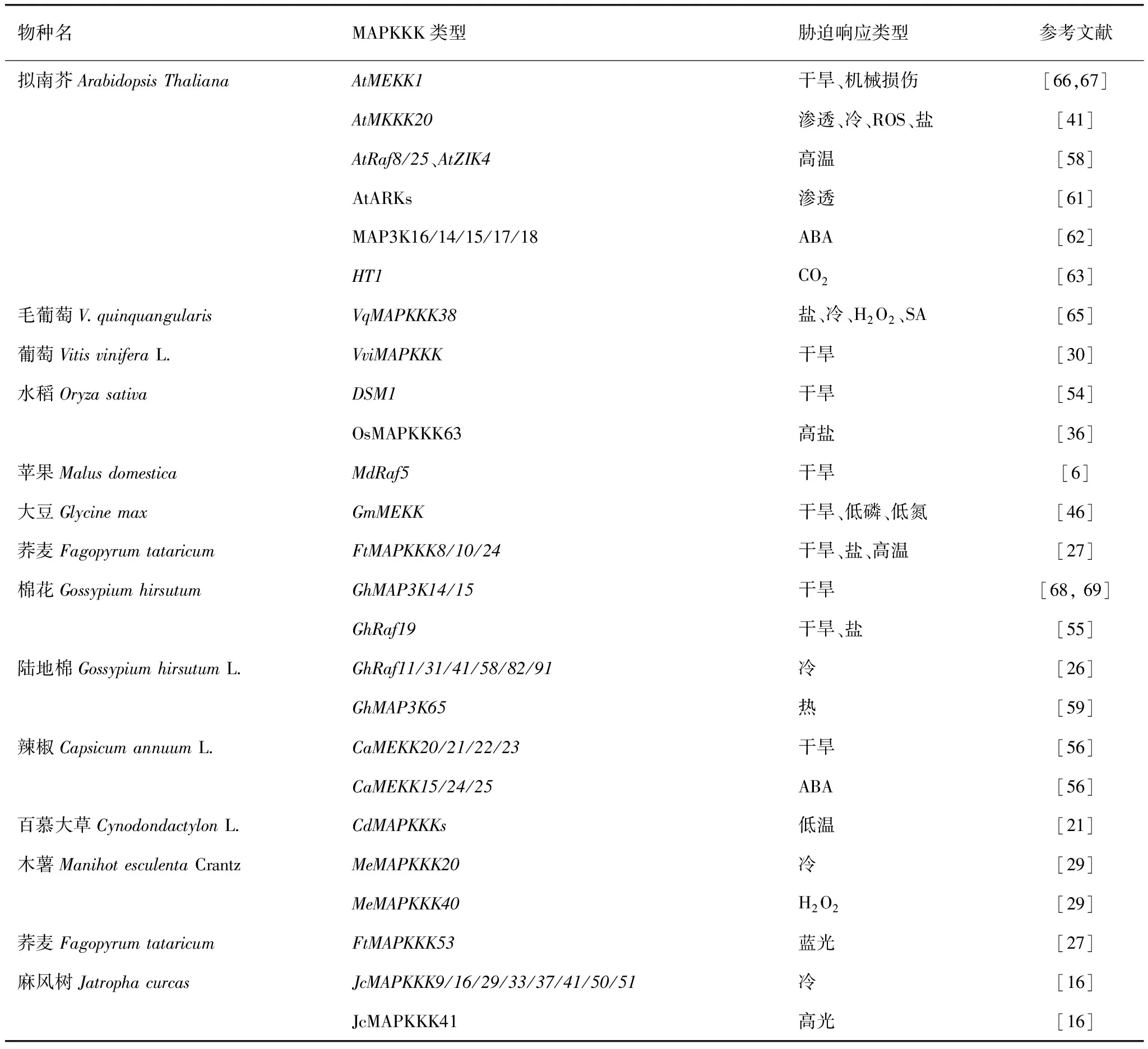
表2 MAPKKK在植物非生物脅迫中的調控應答
2.1 胚胎發育
MAPKKK參與調控植物胚胎發育在宏觀方面表現在影響植物組織、器官和胚胎的發育,在微觀方面可影響植物細胞分裂和分化。Chaiwongsar等[32]研究確定擬南芥MAP3Kε1和MAP3Kε2可在胚胎發育過程中表達,且MAPKKKε1或MAPKKKε2雙突變組合導致胚胎致死性。Xu等[33]研究表明OsMKKK10通過OsMKKK10-OsMKK4-OsMAPK6級聯通路正向調節水稻粒徑和重量,OsMKKK10功能缺失會導致小而輕的籽粒、短穗和半矮稈植株,而構成活性OsMKKK10的過表達會導致大而重的籽粒、長穗和高的植株。Tian等[34]研究發現水稻MAPKKK10-MAPKK4-MAPK6-WRKY53級聯通路在葉片角度和種子大小控制方面發揮作用。Liu等[35]研究指出水稻OsMKKK70介導的OsMKKK70-OsMKK4-OsMAPK6-OsWRKY53級聯通路在控制粒形和葉角中共同發揮作用,過表達OsMKKK70導致植物產生更長的種子,且OsMKKK70、OsMKKK62和OsMKKK55在調節種子大小和葉角方面存在冗余功能。Na等[36]研究發現OsMAPKKK63過表達株系在水稻和擬南芥中均表現出胎生表型,表明OsMAPKKK63可能參與種子休眠控制。Liu等[37]發現水稻OsMAPKKK5的截短變體OstMAPKKK5通過影響細胞大小正向調控水稻株高和產量。Yin等[26]研究發現在陸地棉GhMEKK20和GhMEKK42主要在胚珠中表達,在其他測試組織中低表達,表明二者可能在胚珠特異性發育中發揮作用。
2.2 氣孔發育
氣孔作為植物表皮上的特化氣孔,主要作用是調節植物與外界環境的氣體交換和水分蒸發。已有的研究證實,擬南芥YODA (YDA或MAPKKK4)在氣孔發育中的作用至關重要。Samakovli等[38]發現YODA與熱休克蛋白HSP90相互作用,并通過調節下游MPK3/6激酶的磷酸化來調控氣孔發育,且YODA和HSP90之間的串擾導致轉錄因子SPCH (SPEECHLESS)的不穩定和轉錄失活,從而影響氣孔發育。Lee等[39]研究表明YDA-MKK4/5-MPK3/6級聯通路通過作用于包含RLK ERECTA (ER)/ER-like 1 (ERL1)/ERL2和RLP Too Many mouth (TMM)的受體復合物下游來調控氣孔發育。此外,Guo等[40]研究證明YDA可被定位于細胞皮層的蛋白磷酸酶BSL1激活,從而進一步提高MPK3/6活性,抑制氣孔形成。
2.3 根部發育
根的作用主要體現在固定位置和為植株生長提供養分,MAPKKK參與調控植物根部發育在部分植物中得到證實。在擬南芥中,MAP3K20通過磷酸化下游MKK3激酶和MPK18激酶調節根微管功能,MAP3K20突變體也存在微管組織缺陷。值得注意的是,MAP3K20突變體在正常生長條件下與野生型植物相比發育無明顯缺陷,而經二硝基苯胺除草劑(Oryzalin)處理后的MAP3K20突變體幼苗根明顯短于野生型[41-42]。擬南芥AtMEKK1可通過參與谷氨酸信號通路調控根系發育,AtMEKK1突變體與野生型相比,植株整體矮化,根毛短,側根少[43]。AtMAP3K4作為MAPK6的上游蛋白,在根的發育和有絲分裂過程中微管的調控中發揮重要作用,AtMAP3K4突變體中主根和側根的細胞分裂紊亂,導致細胞質分裂終止[44]。擬南芥MAP3Kε1和MAP3Kε2在根中均有表達,且MAP3Kε1均勻地分布在所有類型的組織中,而MAP3Kε2的表達主要集中在根管組織區域[32]。麻風樹JcMAPKKK64主要在根中表達,JcMAPKKK27在根中高表達,JcMAPKKK9/35/55/58/59在所有測試組織中都高表達,其中JcMAPKKK9在根中表現出更高的表達[16]。棉花GhMAP3K40在本氏煙草N.benthamiana中的過表達抑制了根的生長[45]。
2.4 葉片和花結構發育
已有的文獻報道,MAPKKK在調控植物葉片和花結構發育發揮重要作用。麻風樹JcMAPKKK51在葉片中高表達,JcMAPKKK56在葉片中特異性表達,JcMAPKKK59和JcMAPKKK65在葉片中具有更高的表達水平,表明其在葉片特異性發育中發揮作用[16]。GmMEKK在大豆葉片中的表達水平最高,其次是根、莖、花、莢,在種子中沒有表達,這表明GmMEKK在大豆中可能具有重要的生物學功能[46]。水稻中一個新MAPKKK基因OsCSL1的破壞導致含有積累的淀粉顆粒的葉綠體和脫氯幼苗的嚴重破壞,且OsCSL1的突變可導致能量缺乏和水稻幼苗過早死亡,表明OsCSL1對水稻葉綠體的發育至關重要[47]。水稻OsMKKK10介導的OsMKKK10-OsMKK4-OsMPK6級聯通路參與調控水稻花序結構發育[48]。擬南芥YODA-MKK4/5-MPK3/6級聯通路在受體樣蛋白激酶ERECTA的下游調節局部細胞的增殖,從而使花序結構發生改變[49]。龍葵MAPKKK基因ScFRK1參與調控胚囊和花粉發育,ScFRK1的下調嚴重影響胚囊和花粉的發育,并導致授粉后產生部分孤雌果實[50]。MAP3Kε1、MAP3Kε2和MAP3K20在擬南芥不同組織中均有表達,尤其是在花粉粒中[42,51]。MAPKKKε1或MAPKKKε2純合子的植株沒有明顯的突變表型,而MAPKKKε1和MAPKKKε2雙突變組合引起花粉致死,在MAP3Kε1/2背景中使用誘導的MAP3Kε1結構可挽救花粉活性,表明MAP3Kε1是擬南芥花粉發育過程中質膜正常功能所必需的[52]。值得注意的是,MAP3Kε1和MAP3Kε2在參與花粉粒落在柱頭前的花粉萌發過程存在功能冗余,且MAP3Kε1/2與MOB1A/1B相互作用,可能通過茉莉酸 (JA)信號串擾影響擬南芥胼胝體積累,在限制花粉萌發中發揮重要作用[51]。
2.5 其他發育過程的調控
MAP3Ks除直接作用于生長發育外,還影響植物的晝夜節律和一些代謝過程,間接影響植物的生長發育[3]。如擬南芥AtZIK4蛋白WNK1 (At3g04910)被證實在體外磷酸化假定的晝夜節律時鐘成分APRR3,并可能通過調節其生物活性參與調控晝夜節律[53]。過表達OsMKKK70水稻植株的粒徑和葉角增大情況類似于增強的油菜素甾醇 (BR)信號突變體的表型,表明OsMKKK70可能在水稻BR信號傳導調控中發揮積極作用[35]。水稻OstMAPKKK5的過表達可導致各種形式的內源性赤霉素(GA)含量增加,尤其是最常見的活性形式GA1、GA3、GA4,進而導致細胞增大,并最終導致株高和產量增長[37]。麻風樹JcMAPKKK51和JcMAPKKK55與乙酰輔酶A羧化酶ACC1和ACC3相互作用,表明其可能在脂肪酸合成和延伸中發揮作用[16]。
3 MAPKKK在植物非生物脅迫中的調控應答
植物作為固著生物難免面臨各種生物和非生物脅迫,并進化出大量復雜的信號轉導機制,以調節其發育并增強其對這些脅迫的抵抗力。植物已經進化出各種環境反應機制,通過激活多種信號轉導途徑來適應環境壓力,將自身損害降至最低。應激激活的分子通路包括多個相互連接的調節網絡,如蛋白激酶信號級聯通路,可有效地將輸入信號轉換為合適的輸出。MAPK級聯途徑是植物響應生物及非生物脅迫最常見的信號通路之一,MAPKKK作為該信號通路的第一接收者和傳遞者,負責將信號放大后依次傳遞給MAPKK和MAPK,最終介導植物對外界信號的應答,在植物響應非生物脅迫中起著不可或缺的調節作用,包括干旱脅迫、鹽脅迫、極端溫度脅迫、滲透脅迫等。
3.1 干旱脅迫
植物對干旱脅迫的生理響應,是一個非常復雜而精妙的過程。干旱脅迫是限制全球植物生產力和分布的主要環境因素之一,研究證實MAPKKK參與植物對干旱脅迫的防御反應。Ning等[54]研究表明水稻Raf 亞家族的DSM1(DROUGHT-HYPERSENSITIVEMUTANT1)可通過清除活性氧(ROS)來調節水稻的耐旱性。Sun等[6]研究認為過表達蘋果MAPKKK基因MdRaf5可導致葉片對干旱脅迫的敏感性降低,這與調控氣孔關閉和蒸騰速率有關。Jia等[55]運用病毒誘導基因沉默 (VIGS)技術獲得GhRaf19轉基因棉花植株,發現其對干旱和鹽脅迫的耐受性增強,表明棉花GhRaf19可負向調節干旱和鹽脅迫。Lim等[56]認為辣椒MEKK基因可能參與干旱脅迫反應,研究發現在干旱脅迫下,除CaMEKK16和CaMEKK17外,辣椒CaMEKK基因表達量在處理2 h后顯著增加,并且CaMEKK20/21/22/23基因在6 h后持續高表達。Wang等[30]研究表明葡萄MAPKKK基因家族成員積極響應干旱脅迫,葡萄VviMAPKKK基因在植株受到干旱脅迫時的表達水平幾乎都呈顯著增加的趨勢,許多VviMAPKKK轉錄物增加了10倍以上,其中有4個轉錄物(VviMAPKKK22/23/51/54)的表達增加了20倍以上。Sha等[46]發現大豆根中屬于MEKK1(STE11/BCK1)類MAPKKK基因的GmMEKK受干旱脅迫后上調表達。Ye等[29]研究發現木薯中MeMAPKKK16/18/19/20/23/30/40在干旱處理2~14 d的表達明顯上調,表明它們可能在木薯響應干旱脅迫中起作用。
3.2 鹽脅迫
鹽脅迫是植物在高鹽環境下面臨的一種生理壓力,可導致植物生長不良、凋零甚至死亡。研究發現,部分MAPKKK參與調控植物對鹽脅迫的響應過程,Yang等[57]研究發現MAPKKK18在鹽脅迫期間持續上調,這表明它在耐鹽性中起著至關重要的作用。Kim等[41]研究表明擬南芥mkkk20突變體(SALK_124389)對高鹽脅迫表現出顯著的敏感性,進一步試驗表明MKKK20通過調節MPK6活性來參與響應高鹽脅迫[41]。Na等[36]揭示了水稻OsMAPKKK63具有體外激酶活性,并且它與水稻MAPKK中鹽脅迫響應的中介OsMKK1和OsMKK6相互作用,提示OsMAPKKK63可能參與了高鹽響應。Chen等[45]研究發現棉花GhMAP3K40正向調節耐鹽性且可受鹽脅迫誘導,高鹽脅迫下過表達GhMAP3K40可提高植物發芽率。Jia等[55]研究表明棉花Raf19在鹽脅迫下抑制ROS相關抗氧化基因的表達,負調控棉花耐鹽性,導致ROS的積累。
3.3 極端溫度脅迫
極端溫度脅迫是限制植物生長發育的主要環境脅迫,主要包括高溫脅迫和低溫脅迫。隨著地球氣候環境的變化,高溫已成為影響主要糧食作物生長發育的重要因素。擬南芥AtRaf8、AtRaf25和AtZIK4在高溫脅迫下表達水平上調[58]。擬南芥YODA與HSP90相互作用并通過激活下游MKK4/5-MPK3/6級聯通路來響應熱脅迫[38]。棉花Raf亞族成員GhMAP3K65可受熱脅迫誘導,沉默GhMAP3K65可提高棉花植株的耐熱性,過表達GhMAP3K65使棉花對熱應激的敏感性增強[59]。低溫脅迫對植物的正常生長代謝產生不利影響,嚴重時可造成凍害現象。棉花MAP3K基因GhRaf19正向調控低溫脅迫,過表達棉花GhRaf19可增加植株耐低溫能力,而GhRaf19的沉默使棉花植株抗寒能力下降[55]。在陸地棉中,GhRaf31/58/82僅在12 h的冷脅迫下高表達,GhRaf41和GhRaf91在冷脅迫下的表達水平較低,GhRaf11的表達水平在冷脅迫下增加[26]。在低溫脅迫下,百慕大草12個CdMAPKKKs和13個CdMAPKKKs分別顯著上調和下調;其中有6個CdMAPKKKs至少在一個時間點受到低溫脅迫的顯著誘導[21]。在冷脅迫下,木薯MeMAPKKK20轉錄物在所有測試時間點都被顯著誘導[29]。麻風樹在冷脅迫處理12、24、48 h后,有8個JcMAPKKKs(JcMAPKKK9/16/29/33/37/41/50/51)的表達均顯著上調,尤其是JcMAPKKK16和JcMAPKKK41在冷脅迫12 h后顯著上調,這表明二者在麻風樹低溫反應中具有潛在的重要功能[16]。
3.4 滲透脅迫
環境干旱和高鹽會造成滲透脅迫,抑制植物生長和產量。植物對滲透脅迫有多種信號通路,其中植物激素脫落酸(ABA)起著重要作用[60]。Lim等[56]研究發現辣椒經外源ABA處理后,CaMEKK15/24/25基因表現出不同程度的誘導,CaMEKK16/20/22基因表達量在處理6 h和12 h后均顯著增加。Katsuta等[61]研究證明擬南芥ABA和MAPKKK家族B3支的非生物脅迫應答Raf-like激酶AtARKs在氣孔關閉應對干旱的核心因子SnRK2介導的滲透脅迫應答中起著關鍵作用。Choi等[62]研究指出擬南芥MAP3K16和MAP3K14/15/17/18參與ABA反應,根據發育階段發揮負或正作用,并且MAP3K16可能通過MKK3和ABR1(ABA REPRESSOR1)發揮作用。Kim等[41]發現擬南芥MKKK20在NaCl、H2O2和冷處理下調控MPK6的活性,結果表明MKKK20可能通過調控MPK6活性參與了對各種非生物脅迫,特別是滲透脅迫的響應。
3.5 其他脅迫
擬南芥Raf亞家族基因HT1(HIGHLEAFTEMPERATURE1)是CO2信號傳導的關鍵調節因子,HT1正向調節CO2反應,并部分參與光誘導的氣孔開放途徑[63]。H2O2是一種重要的ROS,也是導致植物對病原體和非生物脅迫因子反應的級聯反應中的關鍵信號分子[64]。在木薯中,MeMAPKKK40經H2O2處理后在所有時間點都表現出顯著的下調[29]。蕎麥15個MAPKKK基因在藍光處理下表達上調,其中FtMAPKKK53基因的表達上調最明顯[27]。毛葡萄幼葉暴露于外源或H2O2會增加VqMAPKKK38的表達,且VqMAPKKK38在毛葡萄葉片中的過表達可顯著增強水楊酸(SA)誘導的二苯乙烯的積累,同時強烈誘導STS(stilbene synthases)和MYB14的表達[65]。棉花GhRaf19的表達可被H2O2誘導,過表達GhRaf19可激活ROS相關抗氧化基因或酶的表達來增強棉花對冷脅迫的抗性,而沉默GhRaf19使棉花植株抗寒能力下降,產生更大的ROS積累[55]。麻風樹JcMAPKKK41與ABI2(ABA insensitive 2)結合,通過抑制ABA信號通路,在調控氣孔關閉、高光脅迫、對葡萄糖的響應、種子萌發、抑制營養生長等方面發揮重要作用[16]。
4 MAPKKK在植物生物脅迫中的調控應答
4.1 細菌、真菌和病毒侵害
植物有一個復雜的免疫系統,也稱為效應器,可以識別來自不同病原體的毒力蛋白,包括細菌、真菌、卵菌和病毒。MAPKKK作為MAPK級聯通路中最大的家族,在植物對細菌、真菌和病毒侵害的響應過程中發揮重要作用。擬南芥Raf成員ILK5(INTEGRIN-LINKEDKINASE)突變體植株對丁香假單胞菌病原體感染表現出更高的易感性[70]。擬南芥MAPKKKδ-1(MKD1)是完全免疫抵御細菌和真菌感染所必需的,它參與了赤霉素的植物毒素應答和對紫丁香假單胞菌的抗性[71]。擬南芥MAPKKK5基因敲除突變體破壞了幾丁質誘導的MAPK激活和對油菜鏈格孢(Alternariabrassicicola)的抗病能力[72]。水稻Raf亞家族基因OsEDR1通過物理相互作用抑制OsMPKK10.2的活性,從而使植株對稻黃單胞菌致病變種Xoc(Xanthomonasoryzaepv.oryzicola)的抗性減弱[73]。水稻Raf亞族成員OsILA1通過抑制OsMAPKK4-OsMAPK6級聯通路,對白葉枯病(xanthomonasoryzaepv.oryzae)具有廣譜抗性[74]。蘋果MdMAPKKK1在調節對葡萄座腔菌(Botryosphaeriadothidea)的抗性中發揮正向調控作用,過表達MdMAPKKK1誘導病原體非依賴性細胞死亡,沉默MdMAPKKK1降低了蘋果愈傷組織和果實對葡萄座腔菌的抗性[75]。PbrMAPKKK基因與梨抗黑斑病的生命過程呈正相關,VIGS表明PbrMAPKKK82基因增強了梨對黑斑病的抗性,qRT-PCR結果表明梨PbrMAPKKK12/13/53/60/65/82/83/96與黑斑病抗性相關,PbrMAPKKK3/9/11/34/80/81/99/100與黑斑病易感性相關[10]。番茄SlMAPKKKε是與植物免疫相關的細胞死亡信號網絡的正調節因子,SlMAPKKKε的沉默降低了番茄對油菜黃單胞菌和丁香假單胞菌的抗性,導致疾病癥狀的出現和細菌生長的增強[76]。番茄SlM3Kα通過激活MAPK信號,導致與胞內受體抗病蛋白(Nucleotide-binding leucine-rich repeat Receptor,NLR)觸發的免疫相關的程序性細胞死亡 (PCD),且SlM3Kα與SlMai1在本氏煙草葉片中的共表達增強了MAPK的磷酸化,加速了PCD的發生[77]。
MAPK級聯通路是植物免疫信號網絡中的重要節點,通過傳輸來自不同刺激的信號來調節下游防御反應,同時也受病原菌和植物因子的調控。Wan等[78]研究表明TaRaf46介導了條形柄銹菌小麥專化型Pst(Puccinastriiformisf.sp.tritici)中一種富含絲氨酸的效應因子Pst27791的毒力,在小麥重要病原菌Pst感染過程中發揮負調節植物免疫的作用。Zhai等[59]研究認為棉花GhMAP3K65可能通過SA、JA、ET (乙烯)和ROS信號通路響應病原菌侵染,沉默GhMAP3K65可增強棉花對青枯病(R.solanacearum)的抗性,過表達GhMAP3K65可增強植株對青枯病的敏感性。Murphy等[79]報道了與馬鈴薯MAP3K StVIK與RXLR效應子Pi17316互作,且StVIK作為一種易感性因子,可增強感染晚疫病菌(P.infestans)的定植。MAPKKKε與一種寄生菌RXLR-WY型效應子PexRD2相互作用,并干擾依賴于該激酶的植物免疫相關信號通路[80]。最近,REN等[81]發現馬鈴薯StMAP3Kβ2與StMAP3Kε共享相同的下游MEK2-SIPK/WIPK級聯通路,且StMAP3Kβ2同源基因NbMAP3Kβ2的VIGS增強了馬鈴薯感染晚疫病菌的定植,并減輕了番茄抗性蛋白Cf4或黃枝孢無毒蛋白Avr4誘導的細胞死亡。除此之外,病原菌產生的MAPKKK也能誘導植物細胞死亡和ROS爆發。PbMAKKK7-PbMAKK3-PbMAK1/3級聯通路中的PbMAPKKK7是甘藍根腫菌(Plasmodiophorabrassicae)侵染植物的調控因子,可誘導ROS積累和細胞死亡,增強植物對煙草花葉病毒 (Tobacoomosaicvirus,TMV)和灰霉病(B.cinerea)的易感性[82-83]。至今,仍然有許多植物與病原菌相互作用的機制尚不清楚,需要投入更多的研究來探索MAPKKK在MAPK級聯通路中如何調控植物侵染期間宿主細胞的死亡。
4.2 植食性昆蟲侵害
根據已有的研究,MAPK級聯通路中的部分MAPKKK成員參與了植物對植食性昆蟲的防御反應。Guo等[84]研究證明桃蚜(Myzuspersicae)半胱氨酸蛋白酶組織蛋白酶B3(CathB3)的propeptide domain與煙草EDR1-like蛋白互作,進一步揭示了蚜蟲唾液蛋白CathB3是通過穩定EDR1激活維管束中ROS通路,從而抑制蚜蟲對植物韌皮部的攝食,表明EDR1-like在介導煙草對蚜蟲的有效防御中發揮作用。Li等[85]研究揭示了一種新型的植物敏感因子Raf36在MKK2的上游起作用并直接靶向MKK2,負向調節擬南芥對寄生蜂的抗性。Guo等[86]研究揭示了MAP4K4-Raf-MAP2K1-ERK、MAP4K4-MAP3K7-MAP2K4-JNK和MAP4K4-MAP3K7-MAP2K6-p38的3個潛在激活途徑,在克服蘇云金芽孢桿菌(Bacillusthuringiensis,Bt)的毒性中起著關鍵作用,從而使十字花科蔬菜作物“超級害蟲”小菜蛾(PlutellaxylostellaL.)對BtCry1Ac殺蟲蛋白產生高抗性。S?zen等[87]研究表明擬南芥AtMAP3K14-MKK3-MPK1/2/7信號模塊是植物基于JA激活從而抑制海灰翅蛾(Spodopteralittoralis)生長的一個緩慢過程,這一個新發現的MAPK級聯通路對于保護植物不受植食性昆蟲侵害具有重要作用。
5 結語
MAPKKK在調節植物生長發育及對環境的適應過程中起著重要作用,這在許多植物中得到證實。盡管植物中越來越多MAPK級聯通路的作用機制被揭示,但MAPKKK在調控植物株高與產量、花粉發育、金屬離子脅迫、光脅迫、病原菌侵染、植食性害蟲侵害等方面的分子機制有待進一步闡明。利用基因編輯技術敲除或過表達作物中的關鍵MAPKKK來研究它們在植物生長發育與抗逆功能中的分子機制,運用酵母雙雜交、融合蛋白沉降、免疫共沉淀等技術篩選驗證MAPKKK互作蛋白并尋找MAPKKK上游的調控因子,鑒定MAPKKK介導的MAPK模塊下游靶標將有助于更好地理解植物如何響應逆境脅迫,同時培養農藝性狀更好和抗逆性更強的作物也是未來工作的重點。因此,仍需要對更多植物MAPKKK基因的功能進行分析,探究它們在調節植物生長發育和響應逆境脅迫信號的分子機制,進一步更新和完善MAPK級聯通路的調控網絡,從而優化植物育種策略來提高作物對逆境脅迫的適應能力,使病原體和宿主之間的協同進化向有利于植物的方向傾斜。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:25:42
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
