新型全蝎寡肽在體外抗苯并芘毒性的功能及其機制的評價
2023-12-24 18:15:58鄭曼袁燕張風雷馬凱李葉婷顧磊劉杰王殿云
廣東藥科大學學報 2023年6期
關鍵詞:研究
鄭曼,袁燕,張風雷,馬凱,李葉婷,顧磊,劉杰,王殿云
(1.東營市人民醫院,山東 東營 257091;2.深圳大學過敏反應與免疫學研究所,廣東 深圳 518060)
動脈粥樣硬化(atherosclerosis,AS)是一種慢性血管炎癥性疾病,其特點是動脈內膜受損和炎性細胞浸潤,參與其中的細胞和分子眾多[1-2]。每年全球有近2 000 萬人死于心血管疾病(cardiovascular disease,簡稱CVD)。作為心肌梗死等多種CVD 的病理基礎,AS 對人類健康構成了重大的公共衛生問題[3]。目前,AS 的預防和治療主要是通過控制心血管危險因素并結合患者的臨床癥狀給予抗血小板藥物、抗缺血藥物治療等[4]。然而,長期大量服藥可能產生耐藥及藥物相關不良反應,給患者帶來巨大經濟負擔及心理負擔[5]。多環芳烴(polycyclic aromatic hydrocarbons,簡稱PAHs)是一類廣泛存在于環境中的污染物,可通過吸入、飲食和皮膚接觸等途徑攝入。研究表明,PAHs 的暴露會增加AS 形成和CVD 發病的風險[6-7]。而苯并芘(Benzopyrene,BaP)則為PAHs中含量最多且毒性最強的化合物之一,早在2010年已被列為I類致癌物[8]。
全蝎是指東亞鉗蝎(Buths martensii)的干燥體,被廣泛應用于民間與臨床治療風濕痹癥、痙攣抽搐、惡瘡腫毒等疾病[9]。現代藥理研究證實,全蝎具有抗腫瘤[10]、抗癲癇[11]、抗驚厥、抗凝、抗菌[12]等多種藥理作用,并且還有鎮痛抗炎和提高免疫力等活性[13]。蝎毒、甾體衍生物和生物堿等小分子成分是全蝎藥理活性的重要基礎[14]。作為傳統藥食同源類中藥,我國全蝎資源十分豐富,但現有研究主要集中在蝎毒等生物活性成分的鑒定和臨床應用上,如芪蝎活血通絡湯(主要成分為全蝎)對于腦血栓的形成,具有顯著的抑制效果[15];同樣以全蝎為主要成分的復方芎蝎膠囊,在臨床上可有效改善頸動脈粥樣硬化患者的腦供血不足的癥狀[16];然而對于全蝎中具有抗AS 活性的肽類單體化合物的研究卻并沒有見到相關報道。本研究旨在通過構建苯并芘(BaP)誘導的人臍靜脈血管內皮細胞(HUVEC)細胞氧化和炎癥損傷模型,追蹤分離全蝎中具有活性的肽,并研究其抗炎和抗氧化活性以及潛在的分子機制,以期為發現全蝎中潛在緩解和治療AS相關活性物質,為功能性食品或藥物的研發提供實驗基礎。
1 材料與方法
1.1 材料與試劑
人臍靜脈血管內皮細胞(中國科學院上海細胞生物研究所);DMEM 基礎培養基、胎牛血清-FBS和青霉素-鏈霉素混合液(美國Gbico);BCA 蛋白質濃度測定試劑盒(碧云天生物技術研究所);Cell counting KIT-8(CCK-8)(日本同仁);ELISA 試劑盒(ROS、IL-1β、IL-6、IL-8、TNF-α、PGE2、ICAM-1 及VCAM-1)(北京四正柏生物科技有限公司);COX-2、ASC、Caspase-1、NLRP3、p62、LC3-Ⅱ、Pink1、Parkin和β-actin 的一抗抗體(美國CST);山羊抗兔二抗和山羊抗鼠二抗(美國CST);苯并芘(美國MCE)。
1.2 主要儀器
SW-CJ-IC 型超凈工作臺(蘇州凈化);INCO 2108 型CO2培養(Memmert,德國);2K15 型低溫高速離心機(Sigma 公司,美國);IKA-Labortechnik 渦旋振蕩器(Staufen,德國);3-30K 高速臺式冷凍離心機(Sigma 公司,美國);Multiskan ?FC 酶標儀(Thermo Fisher,美國)。
1.3 方法
1.3.1 全蝎活性肽分離、氨基酸序列鑒定 首先制備全蝎的總蛋白,后超濾分離全蝎總蛋白,利用疏水色譜法、陰離子交換色譜法、
反相高效液相色譜(RP-HPLC)分離等,最終分離純化得SOP-1(301.5 mg)、SOP-2(432.8 mg)、SOP-3(227.2 mg)、SOP-4(355.6 mg)。通過質譜儀進行ESI-MS 分析。使用Edman 降解法進一步驗證SOP-1~4的氨基酸序列[17]。
1.3.2 細胞活力檢測 將HUVEC 細胞按5 000 cells/well的密度接種于96 孔板中,并按以下方法處理細胞:①空白(Control)組不做任何處理,第2~6組,用10 μmol/L 的BaP 處理12 h,第3~6 組分別用20 μmol/mL 的SOP-1~4 孵育24 h,20 μmol/mL 的谷胱甘肽(glutathione,GSH)作為對照組;②不同劑量的SOP-1(0、10、50、250、500及1 000 μmol/L)孵育24 h;③空白組不做任何處理,第2~5 組,用10 μmol/L 的BaP處理12 h,隨后在第3~5組用20 μmol/L的SOP-1分別孵育24、48、72 h;④空白組不做任何處理,用10 μmol/L的BaP處理12 h,隨后第3~5組分別用10、50、250、1 000 μmol/L的SOP-1孵育24 h。分別檢測其細胞活力[18-19]。
1.3.3 ELISA 法檢測細胞炎性因子及ICAM-1、VCAM-1 含量 將HUVEC 細胞按1×106cells/well的密度接種于6 孔板中,Control 組不做任何處理,BaP組、10 μmol/L組和50 μmol/L組首先用10 μmol/L 的BaP 處理12 h;隨后10 μmol/L 組和50 μmol/L組再分別用10 μmol/L 和50 μmol/L 的SOP-1 處理24 h;收集細胞培養上清,按照ELISA試劑盒的操作說明檢測ROS、TNF-α、IL-1β、IL-6、IL-8、PGE2、ICAM-1及VCAM-1含量。
1.3.4 Western blot檢測相關蛋白的表達 按照方法“1.3.3”處理細胞,胰蛋白酶消化并收集細胞,提取細胞總蛋白;在SDS-PAGE凝膠上分離蛋白;隨后將分離的蛋白轉移到硝酸纖維素膜上;將一抗(COX-2、ASC、Caspase-1、NLRP3、p62、LC3-Ⅱ、Pink1、Parkin 和β-actin)用一抗稀釋液按1∶1000 稀釋,在4 ℃條件下孵育過夜;將二抗同樣按照按1∶1 000稀釋,用相應的二抗室溫孵育1 h;加入顯色液后在WB顯影儀中顯影。
1.3.5 數據處理 采用SPSS 18 軟件統計,實驗結果均以表示,n=3。采用獨立樣本t檢驗,對SOP-1 處理效應進行回歸分析。P<0.05 為差異有統計學意義。
2 結果與分析
2.1 GSPs鑒定
基于MS/MS 數據解析及結合Edman 降解法驗證,共分離并鑒定出4 種活性肽,見圖1。SOP-1 為氨基酸序列為Pro-Met-Tyr-His-Glu-Pro 的寡肽,SOP-2 為氨基酸序列為Phe-Thr-Glu-Gly-Pro 的寡肽,SOP-3為氨基酸序列為Phe-Arg-His-Ala-Leu-Ser的寡肽,SOP-4 為氨基酸序列為Trp-Glu-His-Lys-His-Ala的寡肽。

圖1 SOP-1~4 MS圖譜及結構Figure 1 SOP-1~4 MS and structure
2.2 SOPs抗BaP損傷的活性評價
實驗結果如圖2A 所示,BaP 暴露損傷HUVEC細胞,使細胞活力顯著降低;與Control 組相比,BaP暴露HUVEC 細胞12 h 后,細胞活力降低21.6%(P<0.01)。而SOPs及GSH 能夠緩解BaP暴露導致的HUVEC 細胞損傷,提高細胞活力;相比于BaP 處理組,SOP-1、SOP-2、SOP-3 及SOP-4 組細胞活力分別提高19.3%(P<0.01)、13.4%(P<0.05)、13.7%(P<0.05)及15.2%(P<0.05);其中,SOP-1 保護活性最佳,且SOP-1 減輕BaP 暴露導致HUVEC 細胞損傷具有時間和劑量依賴性(圖2C、2D),因此后續以SOP-1為代表化合物對全蝎寡肽發揮保護活性的潛在分子機制進行研究;同時,結果顯示,濃度不高于1 000 μmol/L 時,SOP-1 對HUVEC 細胞無顯著細胞毒性(圖2B)。
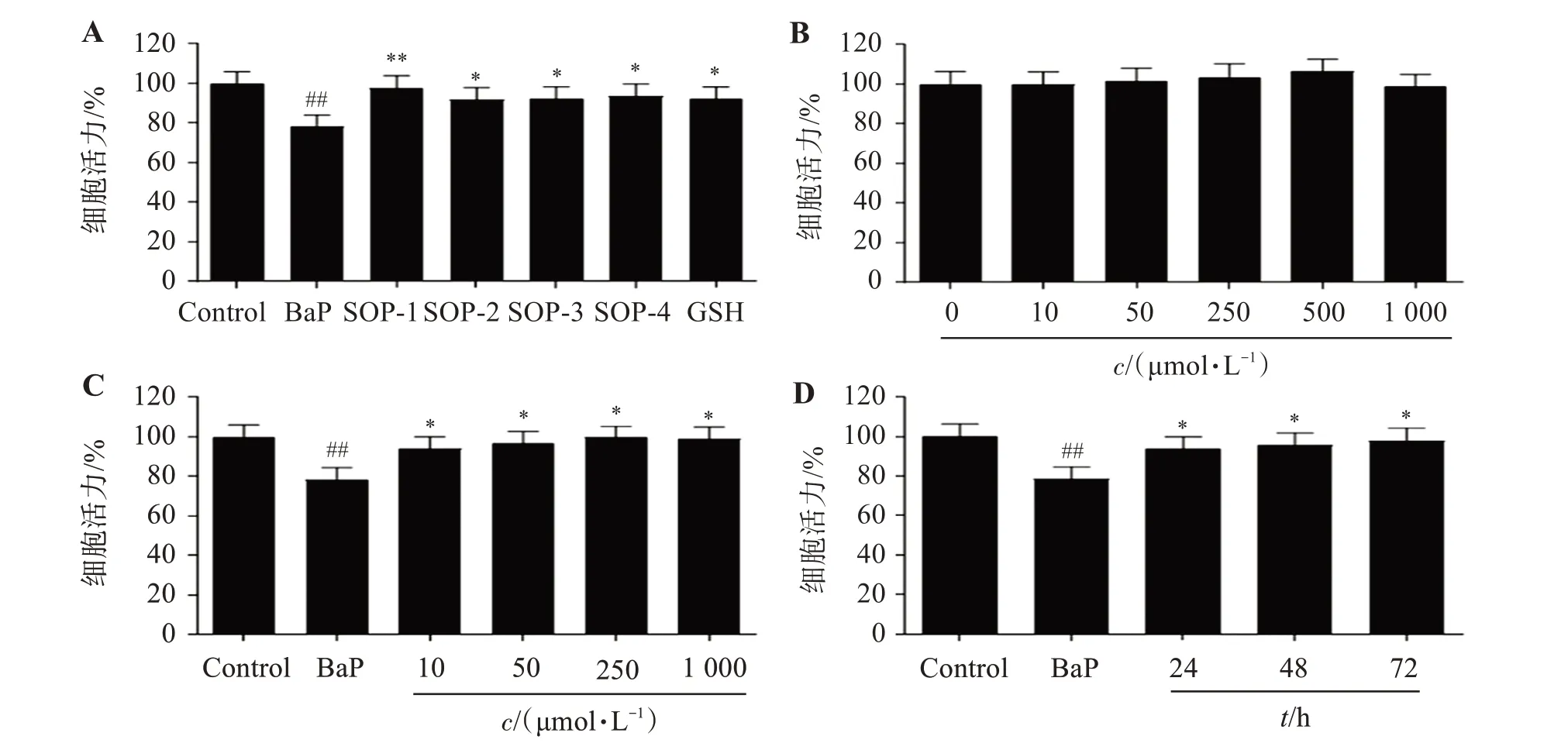
圖2 SOP-1抑制BaP的細胞毒性Figure 2 Inhibition of SOP-1 on the cytotoxicity induced by BaP(n=3)
2.3 SOP-1對BaP誘導的HUVEC細胞ROS及炎性因子過量分泌的影響
研究結果顯示,與Control組相比,BaP處理組的細胞ROS水平顯著增加了451.3%(P<0.01);而相比于BaP組,不同濃度的SOP-1治療后,ROS含量分別減少33.6%(10 μmol/L,P<0.01)、63.5%(50 μmol/L,P<0.01)。表明SOP-1 可能具有抗氧化活性,能夠通過抑制BaP暴露誘導的HUVEC 細胞ROS過量產生,抑制細胞的氧化損傷(圖3A)。
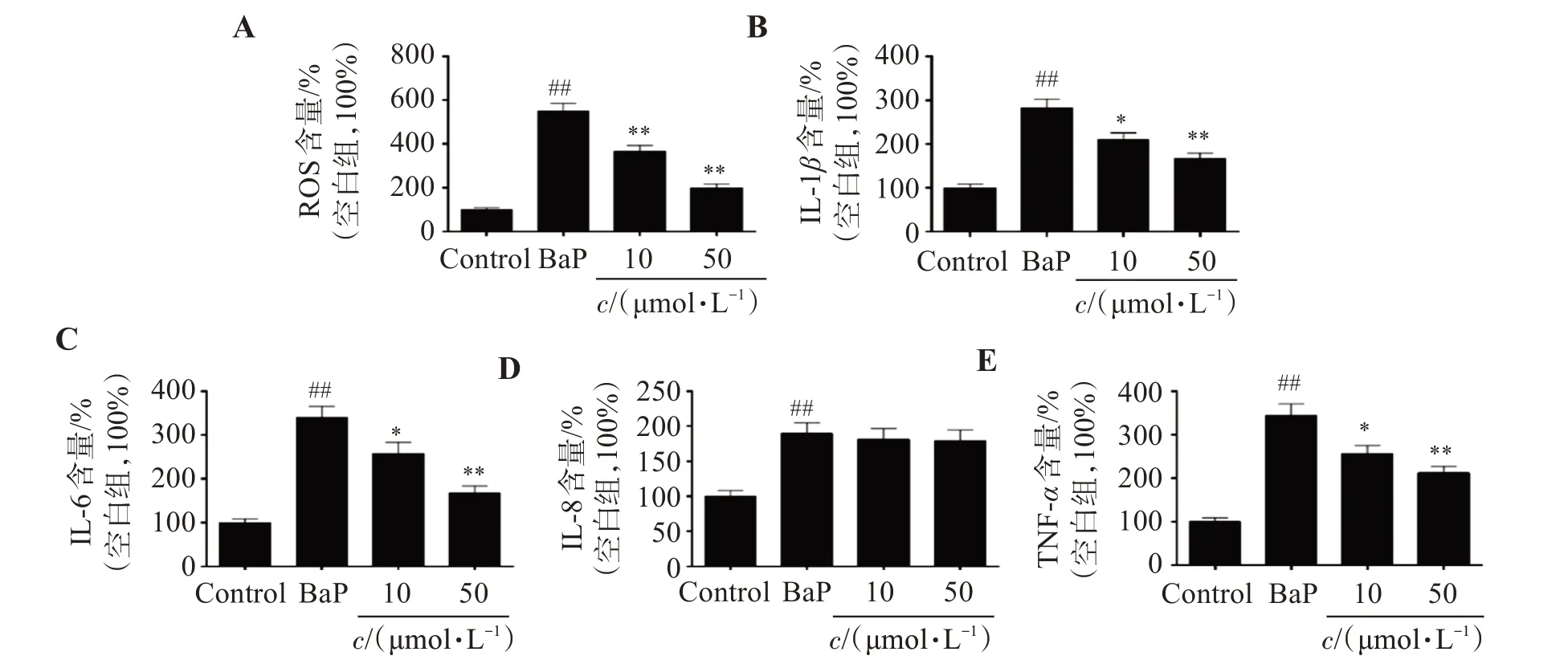
圖3 SOP-1抑制BaP誘導的HUVEC細胞ROS及炎性因子過量分泌Figure 3 Inhibition of SOP-1 on the excessive expression of ROS and inflammatory factors in HUVEC cells induced by BaP
ELISA 的結果顯示,與Control 組相比,BaP 組的IL-1β、IL-6、IL-8 及TNF-α含量分別增加183.4%(P<0.01)、239.7%(P<0.01)、89.7%(P<0.01)和222.2%(P<0.01);而50 μmol/L SOP-1 可顯著抑制BaP 導致的炎性損傷,IL-1β、IL-6、IL-8 及TNF-α的含量分別減少了41.3%(P<0.01)、50.7%(P<0.01)、5.29%及38.4%(P<0.01)。結果表明,SOP-1能夠通過抑制BaP 暴露誘導的HUVEC 細胞炎性因子過量表達、分泌發揮抗炎活性。
2.4 SOP-1對BaP誘導的HUVEC細胞PGE2、COX-2、ICAM-1和VCAM-1蛋白高表達的影響
與Control 組相比,BaP 組PGE2含量增加188.6%(P<0.01);而與BaP組相比,50 μmol/L SOP-1 組PGE2含量減少42.1%(P<0.01,圖4A);說明SOP-1 能夠有效抑制BaP 暴露誘導的PGE2過量分泌。COX-2 是PGE2的合成酶,為了探討SOP-1 對PGE2上游通路的影響,本研究還檢測了COX-2蛋白表達量的變化,結果顯示,BaP 暴露顯著誘導HUVEC 細胞COX-2 蛋白高表達,而SOP-1 則有效抑制BaP暴露誘導的HUVEC 細胞COX-2蛋白表達增加(圖4B)。
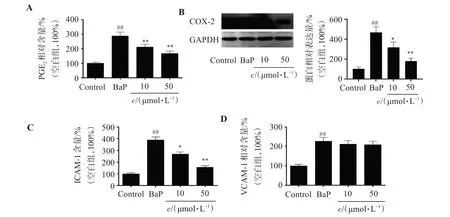
圖4 SOP-1對BaP誘導的HUVEC細胞PGE2、COX-2、ICAM-1和VCAM-1蛋白高表達的影響Figure 4 Effect of SOP-1 on the overexpression of PGE2,COX-2,ICAM-1 and VCAM-1 in HUVEC cells induced by BaP (n=3)
同樣的,對于VCAM-1 和ICAM-1,BaP 暴露會導致其顯著升高,與Control組相比,BaP組ICAM-1、VCAM-1 含量分別增加289.5%(P<0.01)、126.7%(P<0.01);而經過50 μmol/L 的SOP-1 治療后,ICAM-1、VCAM-1 含量分別減少了59.8%(P<0.01)、5.87%(圖4C、D)。
2.5 SOP-1 對BaP 誘導的HUVEC 細胞NLRP3 炎癥小體激活的影響
為了進一步探討SOP-1 抗BaP 誘導的HUVEC細胞炎癥的機制,本研究對NLRP3 炎癥小體相關蛋白的表達進行了檢測。結果顯示,BaP 處理后,NLRP3、ASC 和Caspase-1 的蛋白表達量顯著升高,NLRP3 炎癥小體被激活;而SOP-1 能夠有效抑制BaP 暴露誘導的HUVEC 細胞NLRP3、ASC和Caspase-1 蛋白表達增加,抑制NLRP3 炎性小體激活,SOP-1 的濃度越高,抑制效果越強(如圖5)。說明SOP-1 發揮抗炎活性與抑制BaP 誘導的HUVEC 細胞NLRP3 炎癥小體激活密切相關。
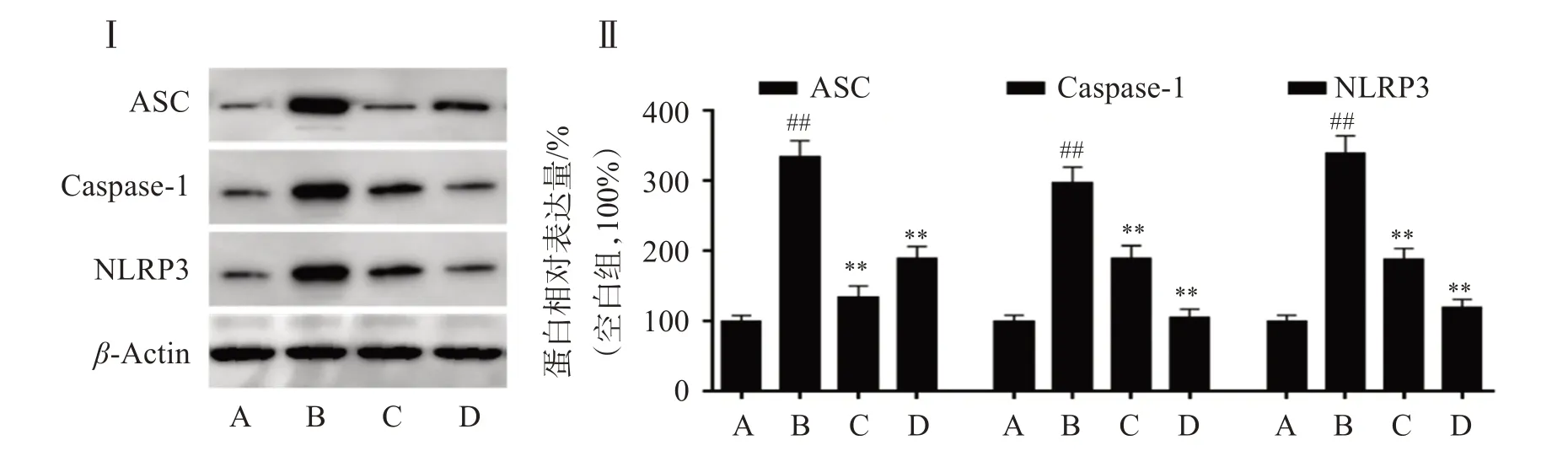
圖5 SOP-1抑制BaP誘導的HUVEC細胞NLRP3炎癥小體相關蛋白表達Figure 5 Inhibition of SOP-1 on the expression of NLRP3 inflammasome associated proteins in HUVEC cells induced by BaP(n=3)
2.6 SOP-1 對BaP 誘導的HUVEC 細胞Pink1/Parkin信號通路激活的影響
為了研究SOP-1 抗BaP 誘導的HUVEC 細胞氧化損傷的機制,對Pink1/Parkin 信號相關蛋白表達進行檢測。如圖6 所示,BaP 暴露導致Pink 1、LC 3-Ⅱ及Parkin 蛋白的表達顯著增加,同時p62 的表達量顯著降低;表明BaP 暴露激活HUVEC 細胞的Pink1/Parkin 通路,可能通過HUVEC 細胞線粒體自噬的途徑導致細胞損傷。而SOP-1 處理后,BaP 暴露導致的Pink 1、LC 3-Ⅱ和Parkin 蛋白表達增加及p62蛋白表達減少均被逆轉,其中50 μmol/L的SOP-1 甚至可以將Pink1/Parkin 通路相關蛋白恢復至接近正常水平。結果表明,SOP-1 發揮抗氧化活性可能與抑制BaP 誘導的HUVEC 細胞Pink1/Parkin 信號通路的激活從而抑制線粒體自噬密切相關。
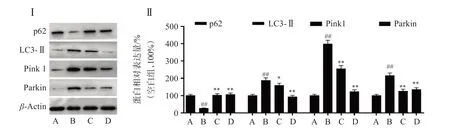
圖6 SOP-1調控BaP誘導的HUVEC細胞Pink1/Parkin通路相關蛋白表達Figure 6 Effect of SOP-1 on the expression of Pink1/Parkin pathway related proteins in HUVEC cells induced by BaP(n=3)
3 討論
PAHs 是重要的環境和食品污染物,它們是煤、煙草等不完全燃燒產生含有苯環的稠環芳烴化合物。研究表明,AS 患者血清中PAHs 的含量明顯高于健康人群,同時在與PAHs 相關的AS 病理反應中,發生了氧化應激和炎癥反應[7]。中藥全蝎具有多種藥理活性,然而,尚未發現針對全蝎中的肽類成分的抗氧化、抗炎和緩解AS 功效方面的研究。本研究使用活性追蹤方法分離了全蝎活性肽,并研究了其在體外的抗AS活性。結果顯示,SOP-1能夠有效抑制由BaP暴露引起的HUVEC細胞損傷。
AS是由多種外界刺激和其他因素誘導形成的,其中氧化應激和炎癥損傷是AS發生的主要機制[20]。COX-2 是一種酶,它在炎癥和傷害時被激活,從而合成包括PGE2前列腺素類物質,已有證據表明,COX-2 的缺失會抑制AS 的發生和發展,因此COX-2/PGE2在AS 中起到重要作用[21]。在本研究中,SOP-1可以通過抑制BaP暴露引發的細胞內ROS和炎癥因子(TNF-α、IL-1β、IL-6 和IL-8)的過量產生,同時抑制COX-2/PGE2的表達,從而緩解了細胞的氧化應激和炎癥損傷,提高了HUVEC細胞的活力。
VCAM-1 和ICAM-1 是表面黏附分子,在AS 模型中介導血管內皮細胞與白細胞的黏附。它們在內皮細胞的表達升高,會導致白細胞募集和炎癥反應[22]。本研究中,BaP的暴露引發了HUVEC細胞中ICAM-1、VCAM-1 以及炎性因子的表達升高,ICAM-1、VCAM-1 以及炎性因子進一步相互作用,加劇了炎癥反應,并對HUVEC 細胞造成損傷。SOP-1 可以通過抑制BaP 暴露所誘導的HUVEC 細胞中ICAM-1、VCAM-1 以及炎性因子的高表達,提高HUVEC細胞的活力,并發揮了保護作用。
NLRP3 炎性小體信號通路是AS 炎癥反應的核心環節,NLPR3 受體識別細胞內積累的脂肪酸、膽固醇結晶等物質,形成激活的NLPR3 二聚體,進而組成炎性小體結構NLPR3/ASC/Caspase-1。該結構能夠裂解炎癥因子前體并增強炎癥反應活性[23]。本研究結果表明,SOP-1 能有效抑制BaP 引發的HUVEC 細胞NLRP3 炎癥小體激活,從而進一步抑制BaP 引起的HUVEC 細胞炎性細胞因子的分泌,并發揮保護作用。
線粒體功能障礙同樣與AS 的發生和發展密切相關。自噬是細胞自我修復的重要途徑之一,其中線粒體自噬通過細胞自噬來清除受損的線粒體,維持線粒體數量和質量的動態平衡。線粒體自噬可以降解線粒體小碎片,從而減少ROS 釋放,減輕氧化應激損傷[20]。Pink1 和parkin 是線粒體自噬關鍵因子。在正常情況下,Pink1 被轉運到線粒體內膜并被泛素-蛋白酶系統剪切和降解。當線粒體受損后,Pink1 在線粒體表面累積和激活,激活的Pink1會招募parkin到線粒體表面,并將其磷酸化,進一步與LC3 適配體蛋白相互作用,最終降解受損的線粒體[24]。p62 是一個重要的蛋白負載體,可以促進受損蛋白通過蛋白酶體和自噬體的清除。自噬激活后,p62 也會被大量清除,因此p62 的減少是自噬活躍的標志[25]。本研究發現,暴露于BaP 的HUVEC細胞,其細胞內ROS 水平明顯上升,相應的線粒體自噬相關蛋白Pink1和parkin的表達上調,而與自噬小體和溶酶體相關的LC3 蛋白的表達增加,p62 蛋白減少。推測BaP 的暴露通過激活Pink1/parkin 通路誘導線粒體自噬,從而對HUVEC 細胞造成氧化應激和嚴重的氧化損傷。本研究結果提示SOP-1可以逆轉這個現象,這表明SOP-1能夠抑制Pink1/parkin 通路的激活,減少線粒體自噬,并減輕BaP 對HUVEC細胞的損傷。
本研究結果表明全蝎中存在的寡肽成分具有抑制由環境污染物BaP 暴露引起的血管內皮細胞NLRP3 炎癥小體和Pink1/Parkin 信號通路激活的能力,從而減輕BaP 暴露引起的血管內皮細胞的氧化和炎性損傷,并發揮保護作用。全蝎中的寡肽類活性物質具有抗氧化和抗炎活性,可以用于預防和治療AS,對功能性食品和藥物研發具有重要的指導意義。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
