高溫脅迫下陸地棉轉錄組差異性分析
2023-12-18 09:27:16董永梅郭偉鋒曹新川郭金成謝宗銘何良榮
新疆農業科學 2023年11期
王 輝,董永梅,郭偉鋒,曹新川,郭金成,謝宗銘,何良榮
(1.塔里木大學農學院,新疆阿拉爾 843300;2. 新疆農墾科學院棉花研究所,新疆石河子 832000;3.作物種質創新與基因資源利用兵團重點實驗室,新疆石河子 832000)
0 引 言
【研究意義】近年來,高溫天氣頻繁出現,高溫脅迫造成作物的經濟損失越來越嚴重[1]。高溫會導致作物的光合作用等生理活動受到嚴重影響,并影響植株的正常生長和發育,是作物減產的主要影響因素[2]。新疆棉區高溫出現時間是6~7月,35℃及以上高溫天氣天數達到9~33 d,而7月是棉花盛花期,高溫的出現造成棉花蕾鈴脫落,導致棉花質量和產量下降[3-4]。因此,高溫對棉花生產的不利影響日益受到關注。【前人研究進展】轉錄組測序技術對于探索植物在不同條件和生長狀態下,可以挖掘和發現新調控機制基因,從而為農作物、生態防護植被、優良林草植物抗逆性遺傳改良以及新品種的培育奠定基礎。尹波等[5]研究在高溫脅迫下,番茄受高溫脅迫后轉錄調控是一個極其復雜的過程,受到多種不同途徑的共同調控,篩選出了36個功能未知的基因。徐洪國[6]研究表明在葡萄中響應高溫脅迫的基因主要包括細胞滲透壓、抗氧化作用、熱激蛋白等基因,NAC轉錄因子家族、HSP20可能在強耐熱品種刺葡萄抵抗高溫脅迫中起著重要的作用。Waters等[7]認為,在高溫脅迫下植物體內會產生大量不同種類的sHSP,是植物對高溫脅迫的一種適應機制。Timperio等[8]強調,當植物受到熱害,HSP作為分子伴侶發揮作用。而Kimpel等[9]則推測sHSPs可能作為熱受損蛋白的保護屏障起作用;另外,sHSPs可以有效地結合變性的蛋白,阻止不可逆轉的蛋白質聚集, 從而提高細胞的耐脅迫能力[10]。【本研究切入點】轉錄組學是一個發現不同差異表達基因的強有力工具,并已被廣泛應用在一些作物品種中,包括水稻[11-12]、玉米[13]、小麥[14]、油菜[15]、馬鈴薯[16]、番茄[17]。其中對水稻,玉米熱應激的反應進行了轉錄組分析。需研究陸地棉高溫脅迫下關鍵基因的響應,比較基因表達差異。【擬解決的關鍵問題】采用轉錄組測序和分析,研究高溫脅迫下棉花不同差異表達上調基因和下調基因的生物學通路,為陸地棉的高溫脅迫研究及陸地棉品種在高溫脅迫下的轉錄變化提供參考。
1 材料與方法
1.1 材 料
材料為陸地棉品系YZ1(由華中農業大學棉花育種組提供),即wide type(W)。
1.2 方 法
采用光照培養室溫度(N,26℃)播種、育苗至5~6葉期,設置正常溫度(N,26℃)處理和高溫(H,42℃)處理,分別在4、12、24、48 h取葉片迅速置于液氮中,于-80℃冰箱保存,進行RNA提取。WN4、WN12、WN24、WN48代表YZ1在常溫處理下4、12、24和48 h,WH4、WH12、WH24、WH48則代表YZ1在高溫處理下4、12、24和48 h。
1.3 數據處理
測序錯誤率用e[18]表示,Illumina平臺測得數據的堿基質量值(Quality score)表示,反映的是堿基識別出錯的概率,進而衡量測序數據的可靠性,則有:Qphred=-10log10(e)。
皮爾遜相關系數r(Pearson’s Correlation Coefficient)為生物學重復相關性的評估指標[19]。r2越接近1,2個重復樣品相關性越強。
RNA樣品檢測合格后,送美因健康科技(北京)有限公司測序,采用Illumina測序平臺技術對構建好的文庫進行高通量測序。使用軟件:用fast qc軟件進行質量控制;使用Hisat2[20]軟件將質控后獲得的Clean Reads與陸地棉參考基因組進行序列比對;利用feature Counts[21]計數,得到每個基因在各個樣本中的原始記數(raw counts)。將數據導入R中,利用DESeq2[22]進行差異表達分析。研究選取|log2F C|>1(log2F C代表處理與對照表達量比值的對數值)及pad j<0.05(pad j代表校正后的P值)的基因作為差異表達基因;使用R語言cluster Profiler包對差異表達基因進行GO富集分析和KEGG富集分析。
2 結果與分析
2.1 測序質量檢查
研究表明,Q phred=-10 log10(e)。Q30的堿基正確識別率達到99.90%,Q20堿基正確識別率為99%,測序數據可靠。表1
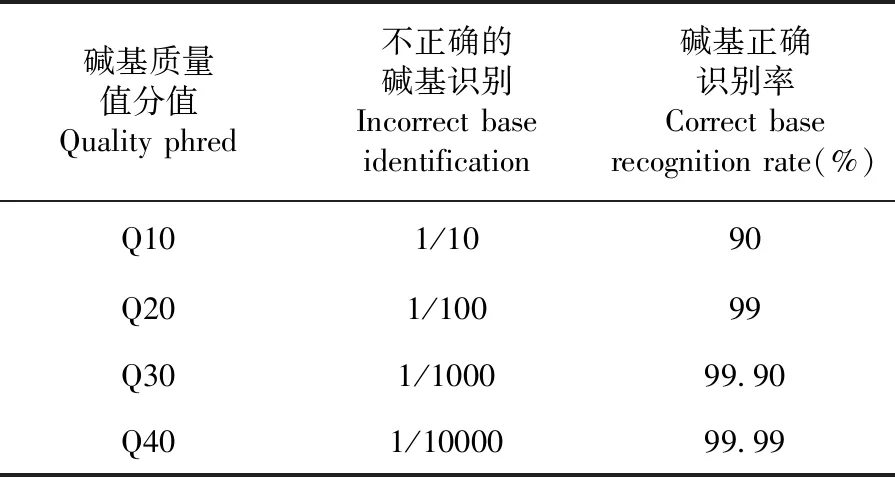
表1 堿基識別正確識別率與Phred分值之間的簡明對應關系
2.2 樣品相關性
研究表明,WN4/WH4,WN12/WH12,WN24/WH24,WN48/WH48兩兩比較后的r2均在0.85以上,兩兩比較的樣本相關性較好。圖1
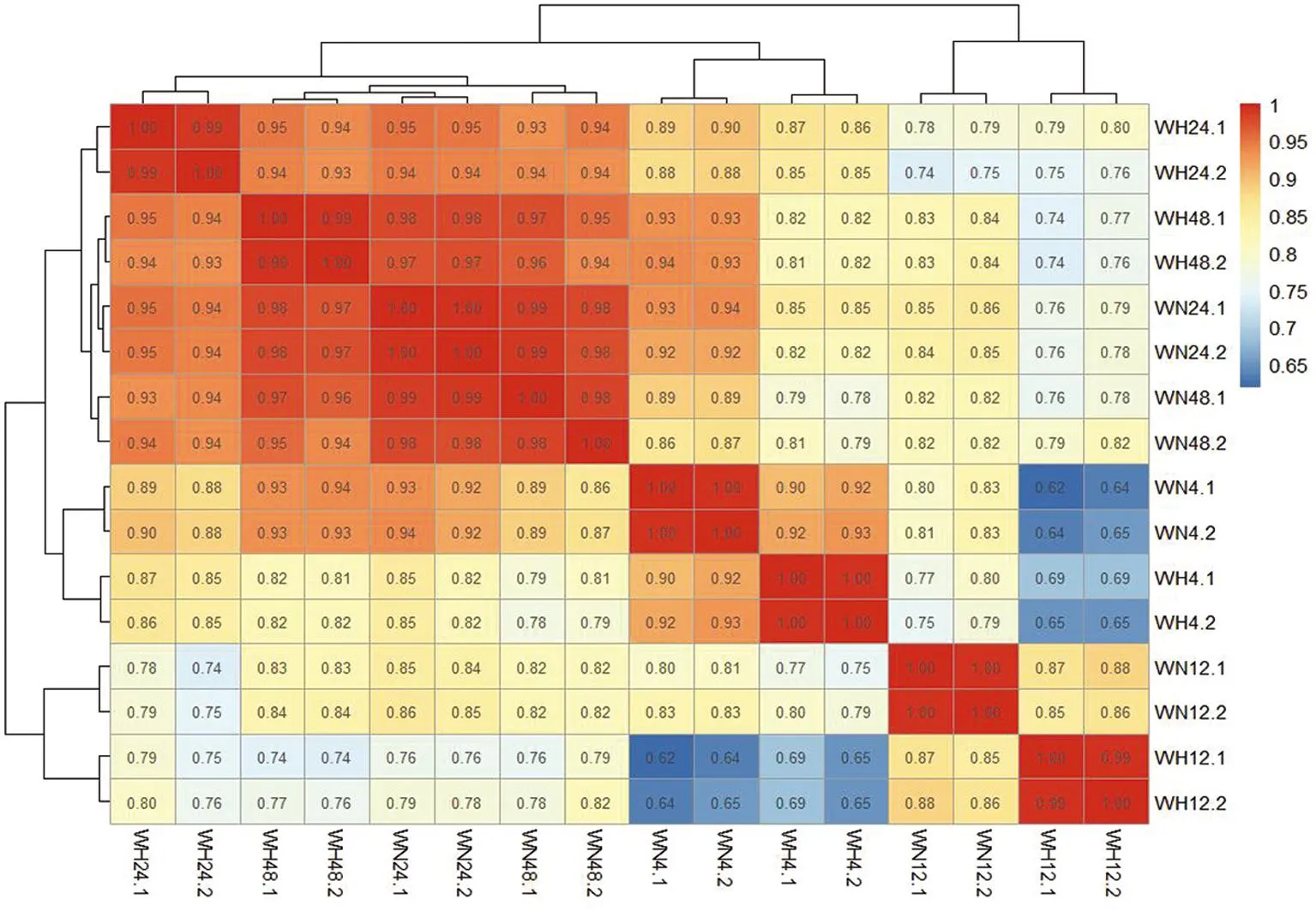
圖1 兩兩樣品的相關性熱圖
2.3 陸地棉差異表達基因
研究表明,隨著時間增加,差異表達基因數量有所下降,在4~12 h時,上調基因增加,下調基因減少,在12~48 h時,上調基因大量減少,下調基因先增加后減少。鑒定出383個普遍上調基因和234個普遍下調基因。隨著熱處理時間延長,WN4/WH4,WN12/WH12,WN24/WH24,WN48/WH48之間特異上調基因的數量呈減少趨勢,上調基因數量分別為2 359個、2 171個、1 092個、418個,下調基因的數量趨于平緩增加,下調基因數量分別為1 733個、1 356個、1 424個、1 490個,下調基因的數量趨于平緩增加。圖2,圖3

圖2 上調和下調基因總數

圖3 上調和下調基因的韋恩圖
2.4 共有差異表達基因GO功能富集
研究表明,差異表達基因主要富集在L-苯丙氨酸分解代謝過程、應激反應、蛋白質復性、類固醇生物合成的過程、超氧化物歧化酶活性的正調節、鋅離子跨膜轉運、蛋白質折疊、系統獲得性抵抗力的調節、金屬離子傳輸等生物學過程,苯丙氨酸轉氨酶活性、催化活性、3-β-羥基-δ5-類固醇脫氫酶活性、過渡金屬離子結合、鋅離子跨膜轉運體活性、金屬離子跨膜轉運體活性、轉移酶活性,轉移酰基、ATP結合、鈣調素結合等分子功能,細胞質細胞組分。圖4

圖4 DEGs基因GO功能富集
2.5 共有差異表達基因的KEGG
研究表明,有241個(P-value<0.000 1)差異基因被注釋。差異基因主要富集在類黃酮生物合成、次級代謝產物的生物合成、苯丙烷代謝、晝夜節律-植物、代謝途徑、酮體的合成與降解、內質網蛋白加工等12個途徑。內質網蛋白質加工富集到17個差異基因都表達下調,17個DEGs與熱激反映有關,高溫脅迫下,熱激蛋白基因下調,受到抑制。表2,表3

表2 17個差異基因熱激蛋白(HSP)
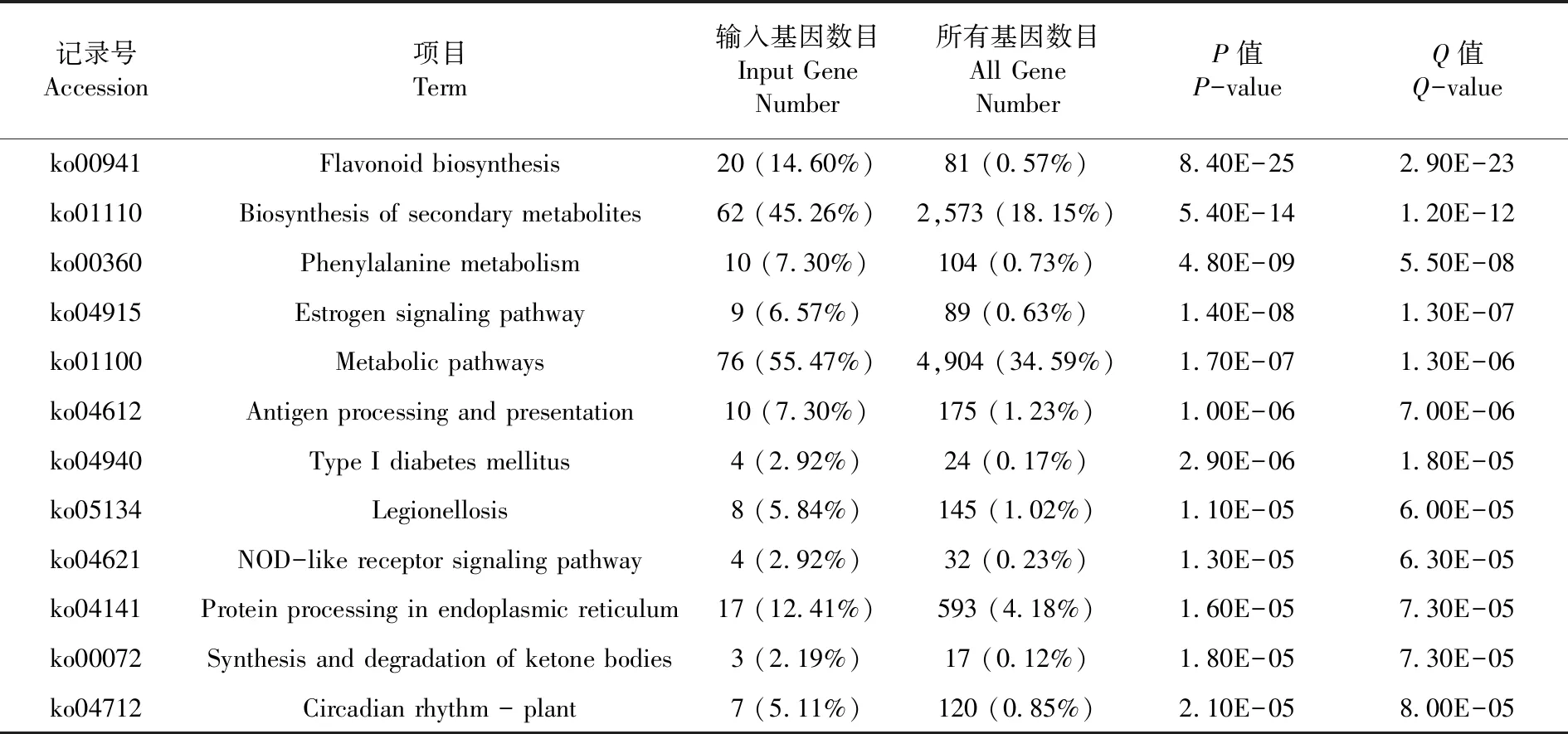
表3 差異基因KEGG富集變化
2.6 轉錄因子表達
研究表明,在差異基因中,鑒定出39個轉錄因子,其主要富集在MYB、WRKY、ERF、NAC等轉錄因子家族,其中WRKY和MYB兩大家族具有最多差異基因,15個和9個差異基因,兩大家族差異基因主要參與高溫脅迫表達,且大部分基因都是上調表達,起著重要作用。表4
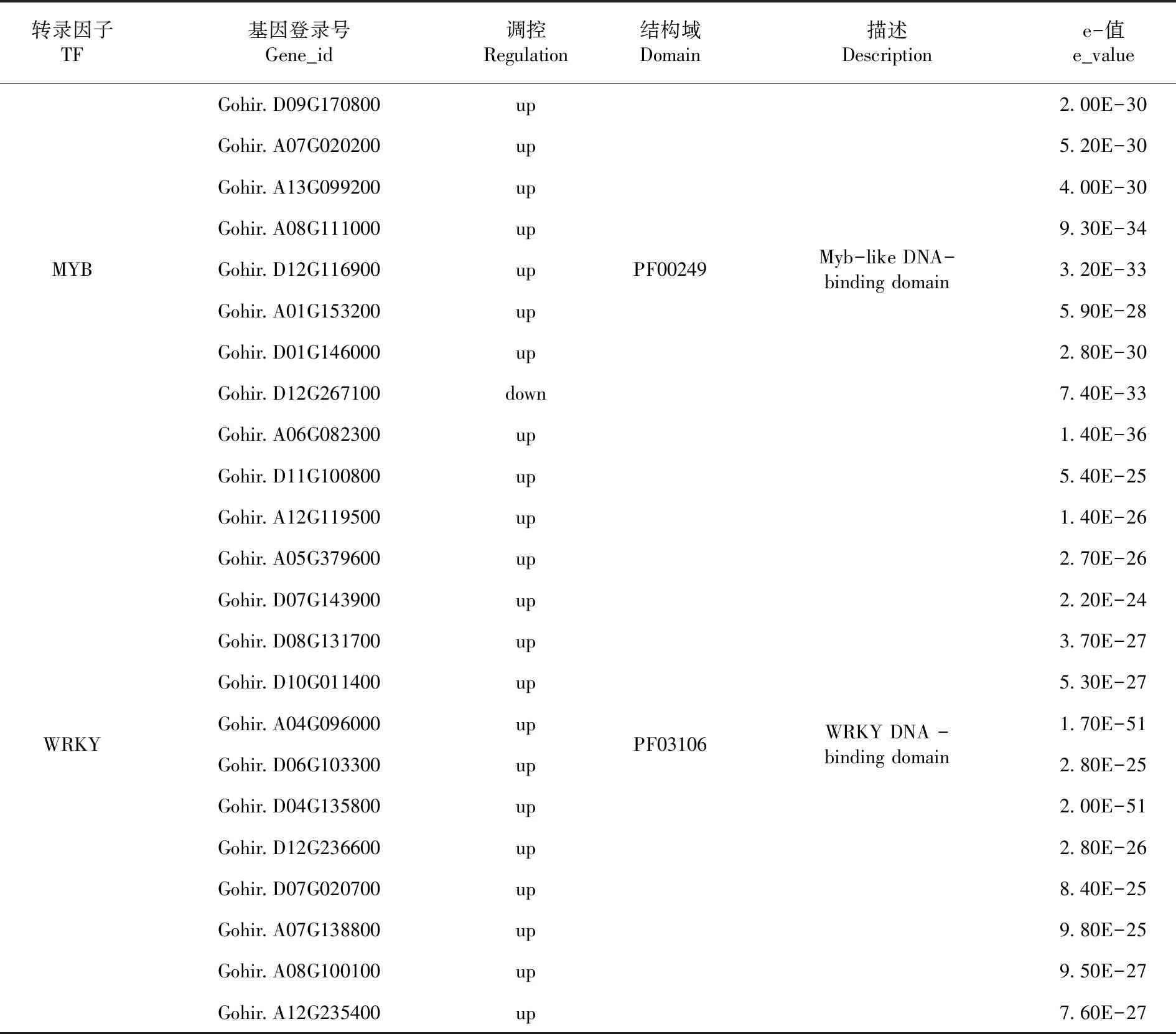
表4 差異基因的轉錄因子
2.7 高溫脅迫相關基因
研究表明,在整個時期表達量差異倍數較為顯著的基因Gohir.A08G104100(Small heat shock protein, chloroplastic,HSP22),熱休克蛋白家族成員,是高溫脅迫主要的基因,對植物的耐熱性至關重要。Gohir.D08G033300(Probable 1-deoxy-D-xylulose-5-phosphate synthase 2, chloroplastic,Os07g0190000),催化丙酮酸的C原子2和3與3-磷酸甘油醛之間的酰基縮合反應,生成1-脫氧-D-木酮糖-5-磷酸(DXP),是質體類異戊二烯生物合成的限制酶,對葉綠體發育至關重要。

續表4 差異基因的轉錄因子
3 討 論
3.1高溫是影響植物生長發育的不利因素,造成大量的產量損失[23]。盡管高溫脅迫對作物的生理效應已被廣泛報道,但對其潛在的分子機制的了解仍然有限。研究對正常溫度和高溫脅迫下的陸地棉進行RNA-Seq測序,鑒定出陸地棉轉錄組上調差異表達基因383個,下調差異表達基因234個;可見,抗熱基因為上調表達,高溫脅迫限制了陸地棉的某些代謝活動;下調表達低于上調表達;與李川等[23]、翟秀明等[24]對植物高溫脅迫下的基因表達上下調結果一致。
3.2差異表達基因GO功能富集分析,陸地棉轉錄組特性與生物過程、細胞成分、分子功能相關;富集到差異基因主要是L-苯丙氨酸分解代謝過程、應激反應、蛋白質復性等生物學過程,苯丙氨酸轉氨酶活性、催化活性等分子功能,細胞質細胞組分。進一步對KEGG代謝通路富集,上調差異基因和下調差異基因共241個被富集到12個KEGG代謝通路,這些基因主要參與類黃酮生物合成,次級代謝產物的生物合成,晝夜節律-植物,苯丙烷代謝,酮體的合成與降解,內質網蛋白加工等途徑。與許小芳[25]富集到顯著通路一致。其中富集到內質網加工途徑的17個熱激蛋白差異基因,表達均下調,為后續研究作為參考。黃酮和類黃酮物質具有自由基清除能力和抗氧化能力,黃酮醇具有保護植物抵抗環境的各種刺激能力[26],在陸地棉中有大量差異表達基因注釋到黃酮和黃酮醇生物合成路徑,這些基因可能與陸地棉響應高溫脅迫有關。上述代謝通路都與植物的抗逆性相關,但個別代謝通路在陸地棉中的調控機理暫不清楚,也為進一步了解陸地棉抗熱機理指明了研究方向。
3.3經過高溫脅迫處理后,MYB、WRKY、NAC、ERF家族大多數成員表達上調,上調應答的轉錄因子在非生物脅迫下能提高植物逆境適應能力,推測這4個家族的大多數成員上調表達能提高植株對熱的適應性。與李鑫雨等[27]結論一致。RNA-Seq是一種在轉錄水平上更為精確的測定分析方法,其通量高、成本低、分辨率高、靈敏度高且不受物種限制[28-30]。
4 結 論
共鑒定出383個共有上調基因和234個共有下調基因,上調差異基因大于下調差異基因。將差異基因按GO功能分類,主要富集到上調差異基因和下調差異基因20個類別,富集到內質網加工途徑的17個熱激蛋白,均表現為下調,鑒定出差異基因主要集中在MYB、WRKY、NAC、ERF這4大家族成員。差異基因顯著富集到類黃酮生物合成途徑、苯丙烷代謝、晝夜節律-植物、內質網加工等通路;鑒定出39個轉錄因子,主要是WRKY和MYB兩大家族;Gohir.A08G104100是熱脅迫主要的基因,對植物的耐熱性至關重要,Gohir.D08G033300是質體類異戊二烯生物合成的限制酶,對葉綠體發育至關重要,2個差異基因在整個時期表達量差異倍數較為顯著。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
