植物SPL轉錄因子研究進展
2023-12-14 14:54:38葛奇席會鵬
安徽農業科學 2023年23期
葛奇 席會鵬
摘要 SPL(squamosa promoter-binding protein-like)基因編碼的綠色植物所特有的轉錄因子,參與植物形態建成、花發育、根發育等過程,調控植物不同發育時期的轉變,維持植物育性,響應植物的外界脅迫,在植物的整個生長發育過程中發揮著重要的作用。對SPL轉錄因子的研究概況、結構特征、同源基因克隆情況、表達調控模式以及生物學功能進行了綜述,以期為進一步進行SPL轉錄因子的相關研究奠定基礎。
關鍵詞 SPL;轉錄因子;基因克隆;表達調控;生物學功能
中圖分類號 Q943.2? 文獻標識碼 A? 文章編號 0517-6611(2023)23-0025-05
doi:10.3969/j.issn.0517-6611.2023.23.006
Research Progress of SPL Transcription Factors in Plants
GE Qi1,XI Hui-peng2
(1.College of Landscape Gardening,Southwest Forestry University,Kunming,Yunnan 650224;2.Xishuangbanna Tropical Botanical Garden,Chinese Academy of Sciences,Mengla,Yunnan 666303)
Abstract SPL(squamosa promoter-binding protein-like) gene encodes a transcription factor unique to green plants,which participates in plant morphogenesis and flower development, root development and other processes, regulates the transformation of plants at different developmental stages,maintains plant fertility,responds to external stresses of plants,and plays an important role in the whole process of plant growth and development. The research situation, structural characteristics, homologous gene cloning, expression regulation pattern and biological functions of SPL transcription factors were reviewed in this paper,in order to lay the foundation for further research on SPL transcription factors.
Key words SPL;Transcription factor;Gene cloning;Expression regulation;Biological function
基金項目 國際合作課題(Y9HX111B02)。
作者簡介 葛奇(1996—),女,云南保山人,碩士研究生,研究方向:風景園林植物資源及應用。*通信作者,高級工程師,碩士,從事植物資源保護及利用研究。
收稿日期 2022-11-07
SPL(squamosa promoter-binding protein-like)基因所編碼的轉錄因子,是綠色植物特有的一類轉錄因子,稱為SPL轉錄因子,廣泛存在于單細胞綠藻、苔蘚、裸子植物以及被子植物中。最早的2個SPL基因由Huijser 等[1]從金魚草(Antirrhinum majus)花序中得到,因其具有能夠識別并結合SQUAMOSA啟動子的活性而被命名為SBP1和SBP2[2-3],即SQUAMOSA啟動子結合蛋白(squamosa promoter binding protein,SBP)。其后又有多名研究人員在各植物中相繼發現多個SPL轉錄因子,并證實該類轉錄因子在植物的形態建成、花器官發育及開花、果實發育和成熟、調控植物次生代謝、影響植物激素信號轉導、參與外界脅迫應答等方面發揮著重要作用。
該研究在前人研究的基礎上,搜集整理SPL轉錄因子近年來的研究成果,概括了當前的研究概況,闡述了SPL轉錄因子的結構特征,說明了SPL基因家族的克隆情況,分析了SPL轉錄因子的表達模式和生物學功能,并對其研究發展趨勢進行了展望,以期為植物SPL轉錄因子的研究發展提供有價值的參考依據。
1 SPL轉錄因子研究概況
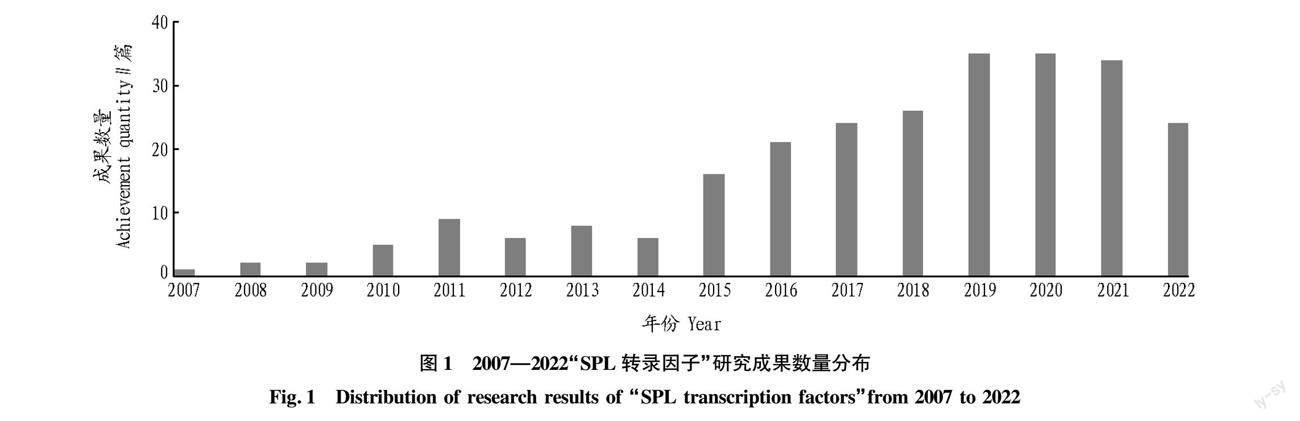
近年來,隨著國內外對于SPL轉錄因子的研究逐漸深入,相關研究文獻資源不斷遞增。對國內外研究成果進行簡要分析,可以促進對SPL轉錄因子研究情況的全面了解,從而準確把握SPL轉錄因子相關的研究動向,促進SPL轉錄因子的研究發展。以中國知網(CNKI)為數據源,設定主題詞為“SPL轉錄因子”,限定學科范圍與植物有關,共檢索到研究論文254篇,其中學術期刊53篇,學位論文196篇,國內會議5篇。相關報道顯示,國內SPL轉錄因子的研究起始于2007年,并在近年快速發展(圖1),內容大都集中在基因功能的研究方面,對于其上游調控基因及下游靶基因的研究相對較少。而國外早在1986年就開始了關于SPL轉錄因子特異性啟動子的研究[4],且內容范圍不僅僅局限于基因功能的研究,在基因的表達調控模式等方面的研究成果也十分豐碩。由此可見,國內對于SPL轉錄因子的研究還應進行更加深入全面地探討,以準確詳盡地掌握其基因表達調控的具體模式和內在的分子機理。
2 SPL轉錄因子的結構特征
SPL轉錄因子帶有一個約由79個氨基酸殘基組成的高度保守的DNA結合域,即SBP結構域(SQUAMOSA promoter-binding protein domain)[5]。SBP結構域是典型的鋅指結構,由8個半胱氨酸(Cys)和組氨酸(His)殘基組成,其中前4個氨基酸殘基結合一個鋅離子,后4個氨基酸殘基結合另外一個鋅離子[6]。目前發現的SBP蛋白中還存在一個位于該結構域C端的保守核定位信號。該信號能夠與第2個鋅指結構發生部分重疊,從而引導SBP蛋白進入植物細胞的細胞核,并進行表達[7]。
目前,在綠色植物中,被鑒定出來并根據基因結構差異分類SPL基因已有許多報道。在擬南芥(Arabidopsis thaliana)中,SPL基因家族共有17個成員。其中第一類成員含有10個及以上的外顯子,所編碼的蛋白質含有超過800個氨基酸殘基,主要包括AtSPL1、AtSPL7、AtSPL12、AtSPL14、AtSPL16;第二類含有2~4個外顯子,其SBP蛋白質不超過400個氨基酸,主要包括AtSPL2、AtSPL3、AtSPL4、AtSPL5、AtSPL6、AtSPL8、AtSPL9、AtSPL10、AtSPL11、AtSPL13、AtSPL15、AtSPL17[8]。在葡萄(Vitis vinifera)等植物中,陳文文等[9]根據miR156靶點的分布,將SPL基因家族分為9個主要分支,其中第一、二、三分支不包含miR156靶基因,第四、五、七、八、九分支其基因的CDS區內含有miR156靶點,第六分支其基因的3UTR區含有miR156靶點,由此可見miR156位點在不同植物中具有較高的保守性。
3 SPL同源基因的克隆分析
已有研究證實SPL基因從簡單的單細胞藻類植物到復雜的高等植物均有分布,如從衣藻(Chlamyydomonas reinhardtii)到小立碗蘚(Physcomitrella patens)再到高等植物,其存在數量不一[3,10]。隨著科技的進步,近年來已有大量SPLs基因被鑒定,如在擬南芥(Arabidopsis thaliana)中有17個,玉米(Zea mays)中有42個,水稻(Oryza sativa)中有19個,番木瓜(Carica papaya)中有10個,葡萄(Vitis vinifera)中有18個,菠蘿[Ananas comosus (L.) Merr.]中有16個,海島棉(Gossypium barbadense)和陸地棉 (G.hirsutum)中各有59 個,而人參(Panax ginseng C.A.Meyer)中則多達106個SPL家族成員[5,9,11-15]。
植物基因克隆是進行生命科學研究的關鍵組成,是進行生命科學研究時較為關鍵的部分[16],對于研究植物基因的表達調控模式和功能至關重要。對于已被鑒定的SPL基因家族成員,在許多植物中已被成功克隆。曾東琳等[17]以“改良香菇”芥藍(Brassica oleracea var.alboglabra)葉片cDNA為模板,克隆得到了“改良香菇”芥藍與葉片發育有關的SPL基因,BoSPL3-1、BoSPL10-2和BoSPL11-2;任子政等[18]以森林草莓(Fragaria vesca)的cDNA為模板,特異擴增出了森林草莓FvSPL基因;張紅雨等[19]利用枸杞(Lycium barbarum L.)花藥cDNA,進行克隆得到枸杞LbSPL6基因;王俊文等[20]利用“寧杞1號”枸杞為研究材料,克隆得到了枸杞花發育有關的LbSPL12基因;吳鴻飛等[21]在花芽分化期的“堰虹桂”桂花(Osmanthus fragrans)中,克隆得到了10個桂花SPL基因,并命名為OfSPL1A、OfSPL1B、OfSPL5、OfSPL6、OfSPL7、OfSPL8、OfSPL10、OfSPL11、OfSPL12、OfSPL13。
4 SPL轉錄因子的表達調控研究
研究表明,SPL轉錄因子一方面受其上游miRNA156/157調控,另一方面通過結合下游基因啟動子區域的順式作用元件,調控下游基因的表達[22]。
miRNA156/157- SPL為植物保守的基因途徑,調控著植物生長發育的許多過程和性狀, miR156/157 是小分子核糖核酸(microRNA),可通過介導SPL 靶基因 mRNA的切割或翻譯抑制,負調控 SPL 基因功能[23]。例如對在黃瓜(Cucumis sativus L.)營養生長時期miR156/157-SPL可調控相關表型的發生,并可能在葫蘆科(Cucurbitaceae)植物營養生長的早期發育中行使特殊功能[24]。在大豆[Glycine max (Linn.) Merr.]的17個含有miRNA156/157識別位點的SPL基因中,有2個能夠被miRNA156通過轉錄剪切調控,有15個能夠被miRNA156通過翻譯修飾調控[25]。在番茄(Solanum lycopersicum)中,SPL基因在莖尖、花序和果實中均高表達,在幼苗、根和葉中低表達,而miR156/ 157則在幼苗、根和葉中高表達,在莖尖、花序和果實中低表達[26]。在馬鈴薯(Solanum tuberosum L.)Stu-miR156過表達植株中,Stu-miR156的表達量在根、莖、葉中均上調,StSPL9均下調;在其Stu-miR156STTM沉默表達植株中,Stu-miR156表達量均下調,StSPL9均上調,且二者共同調控著馬鈴薯的植株高度和側根表型[27]。總而言之,miRNA156/157在SPL的表達調控中具有重要作用,且在大多數含有miRNA156/157識別位點的SPL轉錄因子中, miRNA156/157與SPL的表達水平在植物的生長發育過程中基本都呈負相關[8]。
在SPL轉錄因子調控下游基因的研究中,荔枝(Litchi chinensis Sonn.)LcFT1在不同成熟度末次梢葉和芽中均能表達,且隨成花進程的不斷推進其表達量也不斷降低,LcSPL3、LcSPL10能夠結合LcFT1啟動子,并激活其表達,促進LcFT1與LcFD結合,從而調控荔枝開花[28]。在大豆中,GmSPL9能夠調控下游GmWUS的表達,從而調控腋芽和分枝的形成,提高大豆產量[25]。另外,在水稻中,OsSPL16能夠直接結合GW7的核心順式作用元件,抑制GW7表達,從而調控水稻籽粒的外形[29-30]。由此可見,SPL轉錄因子具有調控植物表型的重要作用。
5 SPL轉錄因子的生物學功能研究
5.1 SPL轉錄因子參與植物的生長發育
5.1.1 參與植物的胚發育。
植物胚胎發育是一個非常復雜的過程,從受精卵到魚雷胚再到器官形成,任何一個階段發生變化都會影響植物正常的生長發育[8,31]。植物DCL1基因編碼的DCL1蛋白與miRNA的合成有關,并參與pre-miRNA剪切[32]。在擬南芥的胚胎發生中,DCL1發生突變時miRNA的合成途徑遭到破壞,致使miRNA合成受阻,進而導致miRNA的靶基因大量表達,其中AtSPL10/11表達水平顯著提高,且當AtSPL10/11發生突變時則可恢復dcl1的部分表型[33]。另外,在柑橘(Citrus reticulata Blanco)體細胞胚胎發生過程中,CsSPL3、CsSPL14下調表達或者csimiR156 上調表達,都能夠顯著促進柑橘體細胞胚胎的發生[34]。因此,植物胚胎發育和體細胞胚胎發生的過程可能有SPL轉錄因子的參與[35]。
5.1.2 參與植物的營養生長。
植物的營養生長是指植物從種子萌發到幼苗形成再到根、莖、葉等營養器官的形成的過程。研究表明,SPL轉錄因子不僅可以調控植物第1個葉原基形成至第2個葉原基形成的時間,調控植物葉片的數量,調控植物葉片的形狀,調控葉片的大小,還可以調控植株的高度,調控植株主根和側根的形成,調控植株側根的長度,調控植株側根的數量等。
在擬南芥中,使AtSPL9/10過表達可以延長植物第1個葉原基形成至第2個葉原基形成的時間,從而降低葉片生成速率;而葉原基的發生則可通過過表達AtSPL13被抑制,進而延緩真葉形成的時間[36-37]。過量表達擬南芥AtSPL2、AtSPL10或者AtSPL11,轉基因植株上最先出現的2片葉會變為橢圓形,部分蓮座葉也會表現出莖生葉的性狀[38]。將牡丹(Paeonia × suffruticosa Andr.)PsSPL基因轉入擬南芥進行異源過表達時,野生型蓮座葉數目平均比轉基因植株多2~3片[39]。此外,水稻OsSPL8基因的突變會導致水稻葉片異常發育,葉耳和葉舌也將無法正常形成[40]。在核桃(Juglans regia L.)中過表達JrSPL1.1時,過表達株系植株葉片大小顯著大于野生型植株[41]。
頂端優勢與植株株型以及植株高度等聯系緊密,Shikata等[38]發現SPL能夠維持植株的頂端優勢。水稻OsSPL4、OsSPL14 2個基因能夠調控水稻株型,與野生型相比,spl4突變體植株具有株高變高、葉片變長、穗分支和籽粒變多、產量增高的表型,過表達OsSPL14植株則會導致水稻分蘗數減少,花序分枝增加,而在SPL4過表達植株中則只能夠觀察到與二者相反的表型[42-43]。
5.1.3 參與植物的花發育。
SQUAMOSA是最早發現的影響花器官發育的因子之一,屬于MIKC類MADS-box基因家族[1,44-45],因此,作為其上游調控因子的SPL在花器官發育過程中發揮著重要的調控作用。
在金魚草中,最早分離得到的SBP1和SBP2在花器官形成便開始表達,從而激活SQUAMOSA基因表達,啟動花器官發育,并在花器官形成后期繼續表達,以維持SQUAMOSA基因的表達活性[2]。在番茄LeSPL3過表達轉基因擬南芥和煙草(Nicotiana tabacum L.)植株中,植株花易脫落,花柄變細,花柄細胞變小,并且能夠提早開花。擬南芥AtSPL具有調節開花時期的作用,白樺(Betula platyphylla Suk.)中BpSPL基因與其高度同源,并且能夠特異性結合BpMADS5基因的啟動子,因此也具有調節花發育的功能[46-47]。SPL的下游基因LFY能夠調控植物花分生組織的形成,維持花分生組織的正常功能,調控花的早期啟動,防止花發育發生逆轉并且控制開花時間[48-49]。王艷艷等[39]發現,與野生型相比,牡丹PsSPL基因的轉基因擬南芥植株中AtLFY基因的表達量顯著上調,推測PsSPL基因主要通過促進下游AtLFY基因的表達促使植株提前開花。
5.1.4 參與植物不同發育時期的轉變。
植物的生命周期一般可分為營養生長和生殖生長2個階段,其中營養生長又可以分為幼年和成年2個時期[8]。過表達擬南芥AtSPL3、4、5、9、10中任意1個基因均可促使其葉片遠軸面產生表皮毛,進而邁入成年期[50]。另外,在擬南芥的生長發育過程中,miR156的表達量隨年齡的增長而降低,其靶基因AtSPL9和AtSPL15的表達量則隨年齡增長而升高,進而正調控下游miR172表達量逐漸升高,從而促使擬南芥發生從幼年期到成年期的轉變[50]。此外, SPL也可以通過調控植株開花可促使植株完成進入生殖生長階段的轉變。擬南芥AtSPL3、AtSPL4、AtSPL5能夠調控植株的開花時間和成花轉變,月季(Rosa chinensis)RcSPL3在聚類分析中與AtSPL3、AtSPL4、AtSPL5處于同一亞組,且在花蕾時期大量表達,因此汪先菊等[51-52]
推測其可能與月季開花時間的調控有關。
5.1.5 維持植物育性。
有性生殖受多種遺傳因素和環境因素的影響,大、小孢子的發生以及雌雄配子的發育對于植物的有性生殖十分重要[53]。首先,擬南芥AtSPL8基因能夠影響大孢子母細胞進入減數分裂的過程,其缺失突變體會導致大孢子母細胞不能形成性母細胞并隨之逐漸退化;其次,AtSPL8基因也能夠影響小孢子囊壁的形成致使其花粉囊發育異常,從而導致植株育性降低。白樺各器官中均有BplSPL8基因表達,且大多數器官中的表達量均顯著低于雄花序[54]。同時,與野生型相比,白樺雄花序發育異常的自然突變體BplSPL8基因的表達量顯著降低,雄花序、花藥及雄配子體的發育明顯延后,雄花序著生部位及小孢子發育明顯異常,花粉敗育,不能散粉[55]。擬南芥三突變體spl8spl9spl15、spl8spl2spl9、spl8spl2spl15以及四突變體spl8spl2spl9spl15的育性比spl8更低。在番茄SlySPL8基因功能的研究中,畢金曦[56]無法獲得Slyspl8b單突變體和Slyspl8a Slyspl8b雙突變體植株,認為可能是由Slyspl8b突變致死導致。由此可見,SPL對于維持植物育性具有重要作用。
5.2 SPL轉錄因子參與植物的次生代謝
植物次生代謝是植物合成生命非必需物質并儲存次生代謝產物的過程,其次生代謝產物可分為苯丙素類、醌類、黃酮類、類萜、生物堿等七大類。研究發現,過表達銀杏(Ginkgo biloba L.)GbSBP9和GbSBP13后,苯丙素生物合成、類黃酮生物合成、植物激素信號轉導等通路有差異表達基因富集,且二者過表達的銀杏愈傷系中類黃酮含量顯著高于對照組,而轉基因擬南芥株系中類黃酮含量也顯著高于野生型[57]。花青素和黃酮醇屬于眾多黃酮類物質中極為重要且常見的2種,增強和降低miR156的表達活性能夠分別促進擬南芥花青素和黃酮醇的物質積累[58]。擬南芥AtSPL8與花青素的生物合成有關,丹參(Salvia miltiorrhiza Bge.)SmSPL12在聚類分析中與其同屬一組,因此,張林甦[59]認為,SmSPL12也有可能參與花青素等次生代謝產物的生成過程。DRF是花青素合成基因,轉錄復合體MYB-BHLH-WD40可以促進其表達,miR156 與SPL9轉錄因子結合,能夠降低SPL9與TT8競爭性結合PAP1的作用,促進 MYB-BHLH-WD40轉錄復合體的形成,從而促進花青素的合成[58,60-61]。在長春花[Catharanthus roseus (L.)G.Don]中,過表達CrSPL9能夠使環烯醚萜途徑中的關鍵酶——馬錢子苷甲基轉移酶基因(CrLAMT)在葉片中的表達量上調[62]。綜上所述,SPL可能在參與植物次生代謝的生理過程中發揮重要作用。
5.3 SPL轉錄因子參與植物的信號轉導
5.3.1 參與光信號轉導。
光照長度和光照時間在植物的整個生命周期中占有十分重要的作用,植物主要是通過光受體感受二者的變化,從而完成植株的光形態建成過程。SPLs基因的轉錄活性受光受體調控,進而影響植株的生長發育。在甜橙[Citrus sinensis (L.) Osbeck]的15個CsSPLs中,每個CsSPL啟動子中均存在光啟動元件[63];在山羊草(Synclisia scabrida)AetSPL1-AetSPL18中,含有TCC-motif、LAMP-element、G-box和GT1-motif等多個光響應元件[64];在水稻SPLs順式作用元件中,也含有多個與調控光周期變化有關的順式作用元件[65],說明SPL轉錄因子參與植株的光信號轉導。
5.3.2 參與激素信號轉導。
赤霉素可以調控植物從種子萌發到花器官發育的整個生長發育歷程。spl8突變體擬南芥植株會表現出花絲變短,萼片表皮毛數量減少,大、小孢子發育異常,育性下降,利用AtSPL8特異性啟動子過表達AtSPL8能夠使spl8突變體植株完全恢復育性[66]。在赤霉素的處理下,AcSPL3在菠蘿[Ananas comosus (Linn.) Merr.]組培苗中的表達量明顯升高,GbSBP1、GbSBP9、GbSBP13在銀杏中的表達量明顯下降[57,67]。以上結果表明,SPLs可能參與赤霉素的信號轉導。
脫落酸能夠抑制細胞分裂,調節植株在逆境中的生長。SPL轉錄因子可能在脫落酸的信號轉導中發揮重要作用。在脫落酸處理下,龍眼(Dimocarpus longan Lour.)DlSPL1、DlSPL5、DlSPL7、DlSPL13基因表達量均顯著下調,且DlSPL1、DlSPL5、DlSPL7中均含有脫落酸響應元件[68]。
茉莉酸甲酯(MeJA)能夠提高植物的抗逆能力。在MeJA處理下,銀杏GbSBP1、GbSBP9、GbSBP13,龍眼DlSPL1、DlSPL7、DlSPL13顯著下調表達,龍眼DlSPL5顯著上調表達,且GbSBP1/9/13、DlSPL5/7/13均含有MeJA響應元件,DlSPL1不含有MeJA響應元件,說明SPLs參與MeJA信號轉導途徑,且調控關系復雜[57,68]。
5.3.3 參與溫度信號轉導。
在溫度敏感型植物的開花過程中,SPL轉錄因子具有重要作用[69]。在高溫處理下,擬南芥spl1-1 spl2-1雙突變體植株表現出超敏感表型[70]。在低溫處理下,桂花OfSPL1A、5、6、10、13的基因表達量顯著上調,miR156-1、2的表達量顯著下調,說明在低溫處理下桂花可能是通過抑制miR156的表達,進而促進OfSPLs進行轉錄,以加快花芽分化的進程[21]。
5.4 SPL轉錄因子參與植物的脅迫應答
植物生長在復雜多變的自然環境中,影響其正常生長發育的環境條件數不勝數,有效應對不利因素,積極進行脅迫應答,是植物能夠生存繁衍的必要條件。已有研究證明,SPL是植物進行脅迫應答的關鍵調控因子。在銀杏的非生物脅迫處理中,GbSBP1、GbSBP9、GbSBP13能夠積極響應鹽脅迫、高溫脅迫、低溫脅迫,且表達模式均相似,且GbSBP1、GbSBP9在干旱脅迫下表達趨勢相同[57]。對水曲柳(Fraxinus mandshurica Rupr.)FmSPL2轉基因煙草植株進行NaCl處理,結果發現轉基因植株耐受性比野生型明顯增強[71]。Aliakbari 等[72]證明,含有干旱脅迫相關順式作用元件能夠使該基因在早期的逆境脅迫響應中發揮重要作用。曹華麒[73]使用3%PEG-400模擬干旱環境,對受到干旱脅迫的藜麥(Chenopodium quinoa Willd.)幼苗進行分析,發現CqSPL1、CqSPL12-1 的表達量增加了 50%,推測二者在干旱脅迫中發揮重要作用。
銅離子對植物的光合作用、木質化程度、花粉和胚珠的發育以及抗病能力具有重要意義,是植物生長發育所必需的一種微量元素。miR398可以通過下游靶基因CSD調控植株維持正常生命活動所需的銅離子含量,以此來應對銅離子脅迫反應[74]。研究證明,缺乏銅離子時,AtSPL7可以結合到miR398基因的啟動子上,促進其進行轉錄;AtSPL7也可以結合miR397、miR408、miR857,誘導其表達,使體內銅離子得以重新分配[8]。
6 展望
SPL作為植物體內一類重要的轉錄因子,調控植物生長發育的多個方面。目前,雖已在金魚草、小麥、擬南芥、水稻等模式植物中鑒定出大量SPL基因,并對其基因結構、生物學功能、表達模式等方面進行了部分解析,但仍然有大量植物尚未完成SPL基因家族的鑒定,且其表達調控的許多機制也尚不明晰,如番茄SlySPL8基因維持植株育性的作用機制,以及擬南芥AtSPL8參與赤霉素信號轉導的作用機制等,相關研究有待進一步深入探索。
參考文獻
[1] HUIJSER P,KLEIN J,LNNIG W E,et al.Bracteomania,an inflorescence anomaly,is caused by the loss of function of the MADS-box gene squamosa in Antirrhinum majus[J].EMBO J,1992,11(4):1239-1249.
[2] KLEIN J,SAEDLER H,HUIJSER P.A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA[J].Mol Gen Genet,1996,250(1):7-16.
[3] CARDON G,HHMANN S,KLEIN J,et al.Molecular characterisation of the Arabidopsis SBP-box genes[J].Gene,1999,237(1):91-104.
[4] BRIGGS M R,KADONAGA J T,BELL S P,et al.Purification and biochemical characterization of the promoter-specific transcription factor Sp1[J].Science,1986,234(4772):47-52.
[5] 陳曉博.參與番茄花柄離區發育的轉錄因子SPL3的基因功能研究[D].北京:中國農業科學院,2010:55.
[6] YAMASAKI K,KIGAWA T,INOUE M,et al.A novel Zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors[J].J Mol Biol,2004,337(1):49-63.
[7] BIRKENBIHL R P,JACH G,SAEDLER H,et al.Functional dissection of the plant-specific SBP-domain:Overlap of the DNA-binding and nuclear localization domains[J].J Mol Biol,2005,352(3):585-596.
[8] 李明,李長生,趙傳志,等.植物SPL轉錄因子研究進展[J].植物學報,2013,48(1):107-116.
[9] 陳文文,吳懷通,陳贏男.SPL家族基因復制及功能分化分析[J].南京林業大學學報(自然科學版),2020,44(5):55-66.
[10] ERIKSSON M,MOSELEY J L,TOTTEY S,et al.Genetic dissection of nutritional copper signaling in chlamydomonas distinguishes regulatory and target genes[J].Genetics,2004,168(2):795-807.
[11] 田晶,趙雪媛,謝隆聖,等.SPL轉錄因子調控植物花發育及其分子機制研究進展[J].南京林業大學學報(自然科學版),2018,42(3):159-166.
[12] 王維維,單玲玲,芮道晨.玉米中SBP-box基因家族的生物信息學分析[J].基因組學與應用生物學,2017,36(1):294-298.
[13] HOU H M,LI J,GAO M,et al.Genomic organization,phylogenetic comparison and differential expression of the SBP- box family genes in grape[J].PLoS One,2013,8(3):1-15.
[14] HULTQUIST J F,DORWEILER J E.Feminized tassels of maize mop1 and ts1 mutants exhibit altered levels of miR156 and specific SBP-box genes[J].Planta,2008,229(1):99-113.
[15] 張磊.SPL轉錄因子研究進展[J] .農業與技術,2022,42(8):25-27.
[16] 李卓雨.植物基因克隆研究進程分析[J].種子科技,2022,40(3):10-12,18.
[17] 曾東琳,唐為玲,曾家晶,等.芥藍SPL基因家族鑒定及表達分析[J].西北植物學報,2022,42(3):361-370.
[18] 任子政,邱安然,常琳,等.森林草莓FvSPL2基因克隆、過表達載體構建及表達分析[J/OL].分子植物育種,2021-12-29[2022-06-25].https://kns.cnki.net/kcms/detail/46.1068.S.20211229.0934.002.html.
[19] 張紅雨,梁新華,石晶.枸杞LbSPL6基因的克隆及表達分析[J].西北植物學報,2021,41(3):377-385.
[20] 王俊文,石晶.枸杞Lb-SPL12基因的克隆及表達分析[J].基因組學與應用生物學,2021,40(2):766-774.
[21] 吳鴻飛,周敏舒,朱守闊,等.桂花OfSPLs基因克隆及其在不同溫度下花芽分化時期的表達分析[J].農業生物技術學報,2020,28(8):1390-1399.
[22] RHOADES M W,REINHART B J,LIM L P,et al.Prediction of plant microRNA targets[J].Cell,2002,110(4):513-520.
[23] HE J,XU M L,WILLMANN M R,et al.Threshold-dependent repression of SPL gene expression by miR156/miR157 controls vegetative phase change in Arabidopsis thaliana[J].PLoS Genet,2018,14(4):1-28.
[24] 汪淑雯,楊愛怡,王華森,等.黃瓜miR156/157-SPL途徑基因鑒定和表達分析[J].園藝學報,2021,48(11):2227-2238.
[25] SUN Z X,SU C,YUN J X,et al.Genetic improvement of the shoot architecture and yield in soya bean plants via the manipulation of GmmiR156b[J].Plant Biotechnol J,2019,17(1):50-62.
[26] SALINAS M,XING S P,HHMANN S,et al.Genomic organization,phylogenetic comparison and differential expression of the SBP-box family of transcription factors in tomato[J].Planta,2012,235(6):1171-1184.
[27] 羅紅玉.馬鈴薯miR156靶向作用SPL基因調控側根發育的機理研究[D].蘭州:甘肅農業大學,2020:40.
[28] 肖秋生.荔枝SPL介導不同成熟度末次梢成花的作用機制[D].廣州:華南農業大學,2019:78-79.
[29] WANG S K,LI S,LIU Q,et al.The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality[J].Nat Genet,2015,47(8):949-954.
[30] KROPAT J,TOTTEY S,BIRKENBIHL R P,et al.A regulator of nutritional copper signaling in Chlamydomonas is an SBP domain protein that recognizes the GTAC core of copper response element[J].Proc Natl Acad Sci,2005,102(51):18730-18735.
[31] 吳艷,侯智紅,程群,等.SPL轉錄因子的研究進展[J].大豆科學,2019,38(2):304-310.
[32] FANG Y D,SPECTOR D L.Identification of nuclear dicing bodies containing proteins for microRNA biogenesis in living Arabidopsis plants[J].Curr Biol,2007,17(9):818-823.
[33] NODINE M D,BARTEL D P.MicroRNAs prevent precocious gene expression and enable pattern formation during plant embryogenesis[J].Genes Dev,2010,24(23):2678-2692.
[34] LONG J M,LIU C Y,FENG M Q,et al.MiR156-SPL modules regulate induction of somatic embryogenesis in citrus callus[J].J Exp Bot,2018,69(12):2979-2993.
[35] FERREIRA E S,ILVA G F,SILVA E M,DE SILVA AZEVEDO M,et al.microRNA156-targeted SPL/SBP box transcription factors regulate tomato ovary and fruit development[J].Plant J,2014,78(4):604-618.
[36] SCHMID M,UHLENHAUT N H,GODARD F,et al.Dissection of floral induction pathways using global expression analysis[J].Development,2003,130(24):6001-6012.
[37] WANG J W,SCHWAB R,CZECH B,et al.Dual effects of miR156-targeted SPL genes and CYP78A5/KLUH on plastochron length and organ size in Arabidopsis thaliana[J].Plant Cell,2008,20(5):1231-1243.
[38] SHIKATA M,KOYAMA T,MITSUDA N,et al.Arabidopsis SBP-box genes SPL10,SPL11 and SPL2 control morphological change in association with shoot maturation in the reproductive phase[J].Plant Cell Physiol,2009,50(12):2133-2145.
[39] 王艷艷,管世銘,蓋樹鵬,等.異源表達牡丹PsSPL基因影響擬南芥營養生長與開花時間[J].植物生理學報,2016,52(8):1207-1213.
[40] LEE J,PARK J J,KIM S L,et al.Mutations in the rice liguleless gene result in a complete loss of the auricle,ligule,and laminar joint[J].Plant Mol Biol,2007,65(4):487-499.
[41] 常英英.核桃WOX和SPL基因在不定根發生中的作用研究[D].北京:中國林業科學研究院,2020:103-104.
[42] 陳廣龍.水稻PPR和SPL基因家族及OsSPL4基因的功能分析[D].武漢:武漢大學,2019:73-75.
[43] JIAO Y Q,WANG Y H,XUE D W,et al.Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice[J].Nat Genet,2010,42(6):541-544.
[44] DAVIES B,SCHWARZ-SOMMER Z.Control of floral organ identity by homeotic MADS-box transcription factors[J].Results Probl Cell Differ,1994,20:235-258.
[45] JACK T.Molecular and genetic mechanisms of floral control[J].Plant Cell,2004,16(S1):S1-S17.
[46] GUO A Y,ZHU Q H,GU X C,et al.Genome-wide identification and evolutionary analysis of the plant specific SBP-box transcription factor family[J].Gene,2008,418(1/2):1-8.
[47] LNNENP M,JNNEN I,HLTT-VUORI M,et al.A new SBP-box gene BpSPL1 in silver birch (Betula pendula)[J].Physiol Plant,2004,120(3):491-500.
[48] MOYROUD E,KUSTERS E,MONNIAUX M,et al.LEAFY blossoms[J].Trends Plant Sci,2010,15(6):346-352.
[49] WEIGEL D,ALVAREZ J,SMYTH D R,et al.LEAFY controls floral meristem identity in Arabidopsis[J].Cell,1992,69(5):843-859.
[50] WU G,PARK M Y,CONWAY S R,et al.The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis[J].Cell,2009,138(4):750-759.
[51] 汪先菊,張紹宇,徐澤,等.桃 SPL 基因家族成員鑒定及其基因表達與果實發育的相關性分析[J].農業生物技術學報,2020,28(9):1521-1534.
[52] 張宇鑫,李惠,張夢,等.月季基因組 SPL轉錄因子鑒定及表達特征分析[J].湖北農業科學,2021,60(21):146-151.
[53] XING S P,SALINAS M,GARCIA-MOLINA A,et al.SPL8 and miR156-targeted SPL genes redundantly regulate Arabidopsis gynoecium differential patterning[J].Plant J,2013,75(4):566-577.
[54] 官民曉,劉雪梅,張妍,等.白樺SPL8轉錄因子基因的分離及轉錄表達分析[J].南京林業大學學報(自然科學版),2013,37(3):17-22.
[55] 劉雪梅,周菲,邢磊,等.白樺雄花突變體及其細胞學特征[J].東北林業大學學報,2010,38(6):1-3,14.
[56] 畢金曦.番茄SPL8基因的功能研究[D].太原:山西大學,2021:42-43.
[57] 萇棒.銀杏SBP-box基因家族生物信息學分析及GbSBP1/9/13功能研究[D].揚州:揚州大學,2021:104-105.
[58] GOU J Y,FELIPPES F F,LIU C J,et al.Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor[J].Plant Cell,2011,23(4):1512-1522.
[59] 張林甦.丹參次生代謝相關基因SmKSL3及SmSPLs的克隆與分析[D].貴陽:貴州大學,2015:101-105.
[60] CUI L G,SHAN J X,SHI M,et al.The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants[J].Plant J,2014,80(6):1108-1117.
[61] YU N,CAI W J,WANG S C,et al.Temporal control of trichome distribution by microRNA156-targeted SPL genes in Arabidopsis thaliana[J].Plant Cell,2010,22(7):2322-2335.
[62] 劉天怡.miR156及轉錄因子SPL在長春花次級代謝中的功能研究[D].大連:大連工業大學,2018:48-49.
[63] 楊杰,陳蓉,胡文娟,等.甜橙SPL基因家族的鑒定及其在成花誘導中的表達分析[J].西北植物學報,2022,42(1):48-56.
[64] 楊紫貽,米福貴,唐芳,等.山羊草SPL轉錄因子基因家族分析[J].中國草地學報,2021,43(10):9-17.
[65] 鐘華.基于連鎖和關聯分析定位水稻籽粒性狀相關QTL及稻屬SPL基因家族分析[D].武漢:武漢大學,2019:75.
[66] MURRAY F,KALLA R,JACOBSEN J,et al.A role for HvGAMYB in anther development[J].Plant J,2003,33(3):481-491.
[67] 趙淑燕.菠蘿AcSPL3基因的克隆及功能分析[D].海口:海南大學,2020:31.
[68] 路保順,朱永靜,張舒婷,等.龍眼SPL基因家族全基因組鑒定及表達分析[J].中國農業科學,2020,53(20):4259-4270.
[69] JAE J K,JEONG H L,KIM W H,et al.The microRNA156-SQUAMOSA PROMOTER BINDING PROTEIN-LIKE3 module regulates ambient temperature-responsive flowering via FLOWERING LOCUS T in Arabidopsis[J].Plant Physiol,2012,159(1):461-478.
[70] CHAO L M,LIU Y Q,CHEN D Y,et al.Arabidopsis transcription factors SPL1 and SPL12 confer plant thermotolerance at reproductive stage[J].Mol Plant,2017,10(5):735-748.
[71] 馬銘浩.水曲柳SPL家族基因克隆及FmSPL2功能分析[D].哈爾濱:東北林業大學,2020:21-23.
[72] ALIAKBARI M,RAZI H.Isolation of Brassica napus MYC2 gene and analysis of its expression in response to water deficit stress[J].Mol Biol Res Commun,2013,2(3):63-71.
[73] 曹華麒.藜麥SPL家族基因的分離和分子特性分析[D].太原:山西大學,2021:29.
[74] ABDEL-GHANY S E,PILON M.MicroRNA-mediated systemic down-regulation of copper protein expression in response to low copper availability in Arabidopsis[J].J Biol Chem,2008,283(23):15932-15945.
