大豆轉錄因子基因GmbHLH130克隆及在干旱脅迫中的功能分析
2023-12-13 15:22:35張斌
江蘇農業學報 2023年7期
關鍵詞:大豆
張斌


摘要: 為了探究大豆GmbHLH130基因在植物干旱脅迫中的調控功能,利用生物信息學分析GmbHLH130與其他物種bHLH家族成員的系統進化關系,檢測GmbHLH130基因及其啟動子對干旱脅迫的響應,對GmbHLH130蛋白的亞細胞定位和轉錄激活活性進行分析,最后初步評估GmbHLH130基因過表達擬南芥的耐旱性。結果顯示,在進化樹中GmbHLH130與擬南芥AtbHLH122進化關系最近;GmbHLH130基因受干旱誘導上調表達;GmbHLH130基因啟動子也受干旱誘導激活下游報告基因;煙草葉片瞬時表達分析結果表明GmbHLH130蛋白定位于細胞核,并且具有轉錄激活活性。此外,GmbHLH130基因過表達擬南芥在干旱處理下的綠色子葉率和根長均顯著大于野生型。本研究結果初步證明了大豆GmbHLH130基因在增強植物耐旱方面的功能,為后續探究其參與耐旱性調控的分子機制提供了理論依據。
關鍵詞: 大豆;GmbHLH130;干旱脅迫;耐旱性
中圖分類號: S565.1?? 文獻標識碼: A?? 文章編號: 1000-4440(2023)07-1441-08
Cloning and functional analysis of soybean transcription factor GmbHLH130 gene under drought stress
ZHANG? Bin
(Hunan University of Science and Engineering/Hunan Provincial Engineering Research Center for Ginkgo biloba, Yongzhou? 425199, China)
Abstract: To explore the regulatory function of soybean GmbHLH130 gene in plant drought stress, bioinformatics was used to analyze the phylogenetic relationship between GmbHLH130 protein and bHLH members of other species. The response of? GmbHLH130 and its promoter to drought stress was detected, and the subcellular localization and transcriptional activation of GmbHLH130 protein were analyzed. Finally, the drought tolerance of? Arabidopsis which overexpressed? GmbHLH130 was preliminarily evaluated. The results showed that GmbHLH130 protein was closest to AtbHLH122 protein in the phylogenetic tree. The expression level of? GmbHLH130 gene was up-regulated under drought treatment. The promoter of? GmbHLH130 gene was also induced by drought stress to activate downstream reporter gene. Transient expression of? GmbHLH130 in tobacco leaves indicated that GmbHLH130 was localized in the nucleus and had transcriptional activation activity. In addition, the green cotyledon percentage and root length of ?GmbHLH130 overexpressed? Arabidopsis were all significantly higher than those of wide type under drought treatment. The results of the study preliminarily proved the function of soybean? GmbHLH130 gene in enhancing plant drought tolerance, and provided theoretical basis for further investigation of its molecular mechanism involved in drought tolerance regulation.
Key words: soybean;GmbHLH130;drought stress;drought tolerance
植物在生長發育過程中經常會受到多種不利環境因素的影響,如高鹽、干旱、高溫和低溫等,其中干旱脅迫是導致農作物減產的重要原因之一[1]。植物在長期的進化過程中形成了多種應對干旱脅迫的方式,如增加葉片蠟質、氣孔關閉等生理響應,關鍵基因的響應引起植物體內滲透調節物質、抗氧化酶活性增加等分子調控[2]。轉錄因子在植物干旱脅迫應答過程中發揮重要作用。目前已知能夠參與植物干旱脅迫的轉錄因子家族包括:MYB[3]、bHLH[4]、bZIP[5]、WRKY[6]和AP2/ERF[7]等。轉錄因子發揮作用的分子機制是其結合到脅迫相關基因啟動子的特定順式作用元件上,進而激活或抑制下游基因的表達,最終防止或減少逆境脅迫對植物的傷害。bHLH家族是被子植物中第二大轉錄因子家族,其廣泛分布于所有的真核生物中。bHLH家族蛋白質能夠結合到基因啟動子區的E-box或G-box元件上調控基因表達[8]。bHLH轉錄因子在植物干旱脅迫響應中的功能及分子機制已有較多報道。例如,過表達擬南芥AtbHLH112基因有利于降低植株體內活性氧水平和提高脯氨酸含量,增強轉基因擬南芥植株對干旱脅迫的耐受性[4]。擬南芥AtbHLH122在干旱和滲透脅迫響應過程中均發揮重要作用,過表達AtbHLH122基因的轉基因擬南芥耐旱性和耐鹽性均明顯增強[9]。bHLH轉錄因子AmDEL則可通過介導黃酮類物質的大量累積增強植物耐旱性[10]。密羅木bHLH家族基因MfbHLH38[11]、日本百脈根LjbHLH34[12]等基因均能正調節植物的耐旱性。轉錄因子bHLH家族成員在植物耐旱性調控過程中的保守功能使其在作物耐旱性育種改良中具有重大價值。
大豆是中國重要的糧油作物,是食用油、植物蛋白和動物飼料的關鍵原料,但是其生產也受到干旱脅迫的影響。盡管在大豆耐旱機理研究和品種選育方面已有較多報道[13-14],但至今仍無法闡明大豆應對干旱脅迫的分子調控網絡。研究結果表明,大豆中有308個bHLH家族成員,其中GmORG3蛋白能夠通過減少根對鎘的吸收和向地上部的轉運增強轉基因大豆的耐鎘性[15],GmPIB1能夠結合到活性氧(ROS)產生關鍵酶編碼基因GmSPOD1啟動子上的E-box元件抑制其表達,減少ROS的產生,進而增強對大豆疫霉菌的抗性[16]。此外,GmbHLH57 和GmbHLH300在大豆鐵穩態維持方面發揮重要作用[17]。然而,大多數大豆bHLH轉錄因子家族基因的生物學功能,尤其是在干旱脅迫應答方面的功能和分子機制尚不清楚。本研究前期從大豆中篩選出1個表達受干旱誘導顯著上調的bHLH家族基因GmbHLH130,結合生物信息學技術、亞細胞定位、轉錄激活活性分析及轉基因擬南芥相關試驗初步探究了其在植物干旱脅迫中的功能。本研究結果將為進一步解析大豆干旱脅迫應答機制和抗旱品種的分子育種提供理論依據。
1 材料與方法
1.1 系統進化樹構建
下載已報道的參與植物非生物脅迫(包括干旱脅迫、鹽脅迫)調控的bHLH家族成員的氨基酸序列,包括MfbHLH38(MT383747)、GmbHLH300(NP_001354033)、OsbHLH148(AK071734)、VvbHLH1(XP_002270239)、AtbHLH17(At2g46510)、FtbHLH3(KU296217)、ThbHLH1(KM101094)、GmbHLH57(XP_003540203)、OsbHLH035(AK106292)、AtbHLH122(At1g51140)、AtbHLH68(At4g29100)、OsbHLH068(Os04g0631600)、AtbHLH112(AT1G61660)。將整理好的氨基酸序列與GmbHLH130(LOC100812802)序列使用Clustal X軟件進行多重比對,利用MAGA-X軟件再次分析氨基酸多重比對結果,然后利用鄰接法(Neighbor-joining,NJ)繪制系統進化樹,參數設置:泊松模型,Bootstrap=1 000。
1.2 大豆種植及模擬干旱處理
試驗用栽培大豆品種為Williams 82。將大豆種子在水中浸泡6 h后均勻鋪在潤濕的濾紙上萌發,待長出胚根(約1.0~1.5 cm)后移至石英砂中,用1/2 Hoagland營養液澆灌,在生長室進行培養,生長環境:溫度25 ℃,空氣相對濕度60%~70%,長日照(晝16 h/夜8 h)。待幼苗的第一對真葉展開后,挑選株高一致的植株放置在黑暗條件下繼續培養24 h。用含有20% 聚乙二醇(PEG)的1/2 Hoagland營養液(干旱處理)澆灌,分別在0 h、3 h、6 h、12 h和24 h取大豆植株的根和葉,液氮速凍后放于超低溫冰箱保存,每個時間點、每個組織均包括3組生物學重復。
1.3 總RNA提取和基因表達水平檢測
使用RNA提取試劑盒[生工生物工程(上海)股份有限公司]提取大豆根和葉的總RNA,使用Nanodrop 2000測定RNA濃度,根據反轉錄試劑盒[SuperScript III First-Strand Synthesis SuperMix,賽默飛世爾科技(中國)有限公司]的說明書操作合成cDNA。以合成的cDNA為模板進行半定量PCR分析基因的表達情況,PCR反應總體系為20.0 μl:10.0 μl Taq Mix[翌圣生物科技(上海)股份有限公司],8.0 μl去離子水,正向和反向引物各0.5 μl,cDNA模板1.0 μl,以大豆GmUBI3基因作為內參基因,本研究所用的引物見表1。
1.4 GUS(β-葡萄糖苷酸酶基因)組織化學染色
以GmbHLH130基因起始密碼子上游1 500 bp序列作為啟動子區,以大豆DNA為模板克隆啟動子片段,插入雙元表達載體PBI101獲得融合質粒PGmbHLH130::GUS。將質粒PGmbHLH130::GUS導入根癌農桿菌GV3101,利用浸花轉化法獲得轉基因擬南芥,并篩選得到T3代純合子。將在正常MS培養基上培養7 d的擬南芥幼苗分別用去離子水和20% PEG處理24 h,然后取幼苗浸沒到GUS染色液[1.44 ml 1 mol/L Na2HPO4、1.06 ml 1 mol/L NaH2PO4、 1.00 ml 100 mmol/L K4Fe(CN)6·3H2O、1.00 ml 100 mmol/L K3Fe(CN)6、500 μl 500 mmol/L乙二胺四乙酸(EDTA)、50 μl TritonX-100、20 mg 5-溴-4-氯-3-吲哚葡萄糖苷(X-Gluc),定容至50.00 ml]中,置于37 ℃,避光染色12 h。倒掉GUS染色液后,用75%乙醇脫色,最后觀察GUS染色情況。
1.5 亞細胞定位檢測
將不含終止密碼子的編碼序列(CDS)片段插入pCAMBIA1300-GFP載體,獲得重組質粒35S::GmbHLH130::GFP并導入根癌農桿菌GV3101。根據已報道的方法[18],將含有重組質粒的農桿菌菌液注射到生長旺盛的本氏煙草葉片中,以注射35S:: GFP空載體的葉片為對照,繼續培養24 h,使用激光共聚焦顯微鏡觀察煙草葉片細胞中的綠色熒光信號。
1.6 轉錄激活活性檢測
克隆GmbHLH130基因全長CDS插入酵母表達載體pGBKT7中,獲得重組質粒pGBKT7-GmbHLH130。分別將空載體pGBKT7和重組質粒pGBKT7-GmbHLH130導入酵母菌株Y2H Gold。經PCR檢測后,選取陽性單菌落接種到酵母浸出粉葡萄糖液體培養基(YPDA)中,待OD600為0.8~1.0時,抽取菌液分別滴到YPDA、SD/-TRP/-HIS和SD/-TRP/-HIS/+X-α-Gal固體培養基上,5 d后根據菌落的生長情況判斷轉錄因子的激活活性。
1.7 GmbHLH130基因過表達擬南芥表型分析
克隆GmbHLH130基因全長CDS插入雙元表達載體pFGC5941的35S啟動子下游。利用根癌農桿菌介導的浸花法轉化野生型擬南芥,將收獲的T0代種子播種在含有5 mg/L草銨膦的MS培養基上篩選抗性植株。此外,提取RNA進行半定量PCR檢測外源基因是否成功高表達,選取外源基因高表達的株系,篩選到T3代,獲得純合材料用于初步鑒定GmbHLH130基因的功能。
取同時期收獲的野生型(Col-0,WT)和轉基因擬南芥種子于75%乙醇中滅菌10 min,在超凈臺中吹干種子,然后均勻播種于MS培養基和20% PEG處理的MS培養基中,在4 ℃冰箱中春化3 d后移至生長室正常培養7 d,然后觀察不同材料的長勢并統計綠色子葉率(葉片能正常生長呈現綠色的植株數占播種下去的種子總數比例)。此外,將表面消毒的擬南芥種子播種并經過4 ℃春化后,在生長室垂直放置培養,7 d后觀察并測定不同材料的根長。
野生型擬南芥和過表達株系(L1和L2)的種子經75%乙醇表面消毒后,均勻播種于濕潤的營養土和蛭石混合(1∶1)基質上,于4 ℃春化3 d后轉移至植物培養室,使用自來水澆灌,保證每盆每次澆水量相同。3周后取長勢一致的擬南芥用于干旱處理,使其自然干旱,觀察野生型和過表達株系的表型變化。
2 結果與分析
2.1 大豆GmbHLH130的系統進化樹和氨基酸序列分析
根據已知報道,本研究整理了參與非生物脅迫調控的bHLH家族成員的氨基酸序列,包括MfbHLH38、GmbHLH300、OsbHLH148、VvbHLH1、AtbHLH17、FtbHLH3、ThbHLH1、GmbHLH57、OsbHLH035、AtbHLH122、AtbHLH68、OsbHLH068、AtbHLH112,與GmbHLH130構建系統進化樹。結果顯示,所選的bHLH家族成員分布在2個不同分支,GmbHLH130與擬南芥AtbHLH112、AtbHLH122、AtbHLH68以及水稻OsbHLH068在同一分支,并且GmbHLH130與AtbHLH122進化關系最近(圖1A)。進一步對比氨基酸序列發現,GmbHLH130與AtbHLH122序列相似度為48.18%,但C端的bHLH結構域十分保守(圖1B)。
2.2 大豆GmbHLH130基因對干旱脅迫的響應
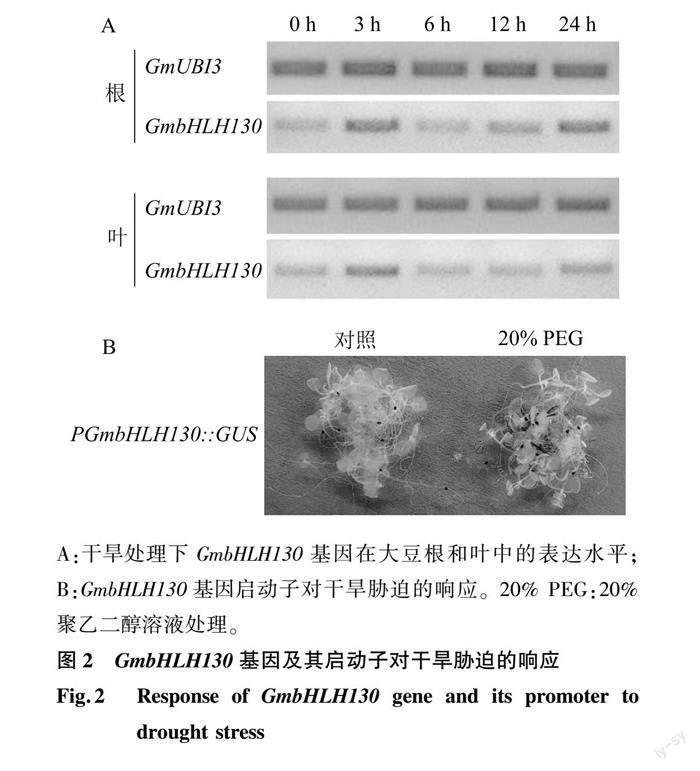
利用PEG模擬干旱脅迫,檢測GmbHLH130基因在大豆幼苗中的表達模式。結果顯示,干旱處理3 h時,GmbHLH130基因表達水平在根和葉中均明顯增加;但隨著處理時間增加,表達水平又恢復至與干旱處理前無明顯差異(6 h和12 h),干旱處理24 h時表達水平又增強(圖2A)。進一步克隆了GmbHLH130基因起始密碼子上游1 500 bp的啟動子片段,構建GUS融合載體,并獲得相應的轉基因擬南芥植株PGmbHLH130::GUS。在MS培養基上生長2周的擬南芥PGmbHLH130::GUS被隨機分為2組(各10株),分別用去離子水和20% PEG溶液處理,GUS染色結果顯示對照組未觀察到藍色,而處理組觀察到明顯的藍色,說明GmbHLH130基因啟動子受干旱脅迫誘導激活β-葡萄糖苷酸酶基因的表達,生成的β-葡萄糖苷酸酶與染色液中的底物反應生成藍色物質(圖2B)。
2.3 GmbHLH130蛋白的亞細胞定位
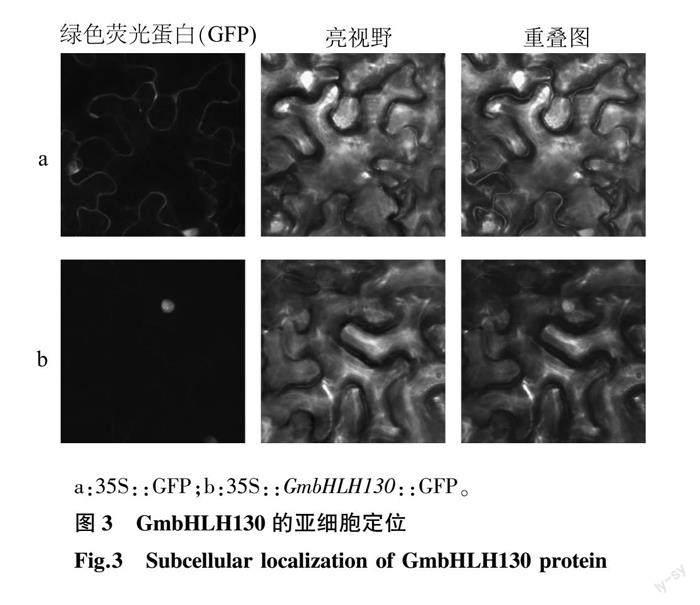
普遍認為轉錄因子蛋白定位于細胞核內發揮轉錄調控作用[18]。為了研究大豆轉錄因子蛋白GmbHLH130的亞細胞定位,將GmbHLH130基因除終止密碼子外的CDS插入pCAMBIA1300-GFP載體,構建35S::GmbHLH130::eGFP融合表達質粒。通過根癌農桿菌介導的方法在煙草葉片中進行瞬時表達,以轉化pCAMBIA1300-GFP空載體(35S:: eGFP)的葉片為對照。結果顯示,35S:: eGFP轉化的煙草葉片細胞核和細胞膜上均觀察到較強的綠色熒光,而僅在35S::GmbHLH130::eGFP融合質粒注射的葉片細胞核中觀察到較強的綠色熒光信號,說明轉錄因子GmbHLH130蛋白定位在細胞核中(圖3)。
2.4 GmbHLH130蛋白具有轉錄激活活性
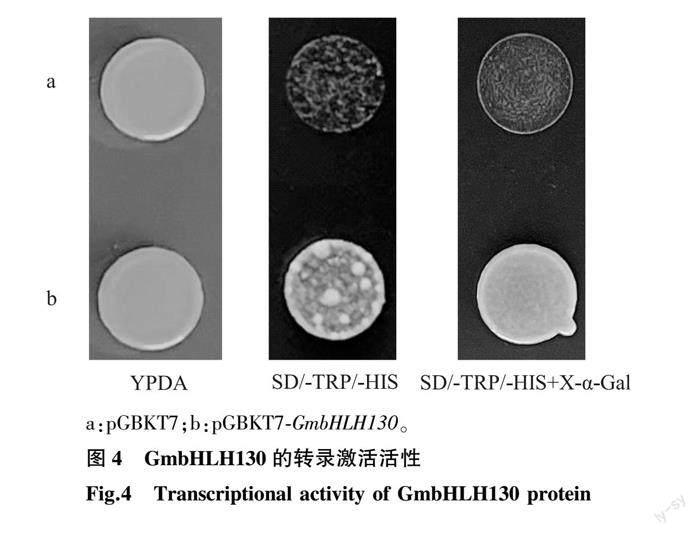
利用酵母雙雜交系統驗證GmbHLH130轉錄因子的轉錄激活活性。結果顯示,含有pGBKT7空載體或pGBKT7-GmbHLH130重組質粒的Y2H酵母菌株都能在YPDA培養基上生長。而在SD/-TRP/-HIS營養缺陷培養基上,導入pGBKT7空載體的酵母菌株不能生長,但含pGBKT7-GmbHLH130的酵母可在SD/-TRP/-HIS培養基上生長,且菌斑在含X-α-Gal的SD/-TRP/-HIS培養基上變藍,說明轉錄因子GmbHLH130能夠激活Y2HGold菌株中HIS、MEL1基因的表達,具有轉錄激活活性(圖4)。
2.5 GmbHLH130基因過表達擬南芥的獲得及耐旱性分析
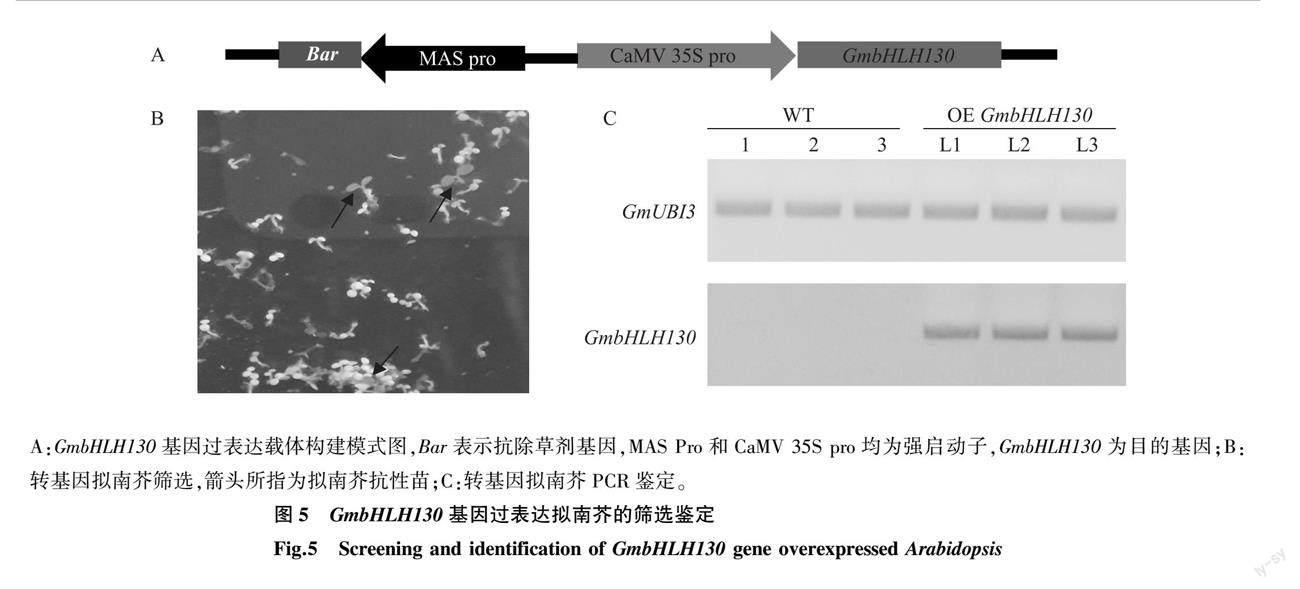
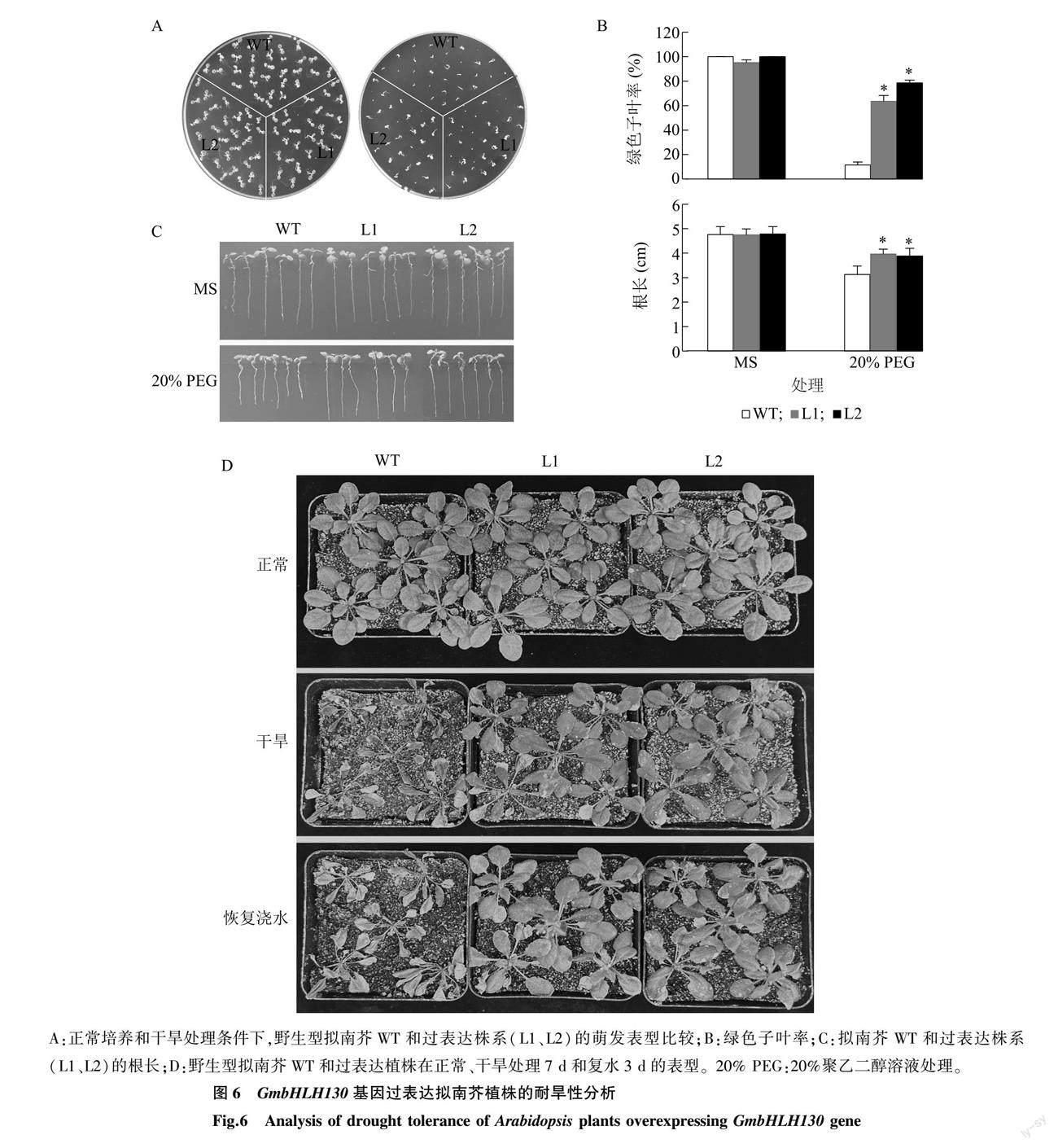
為了驗證大豆GmbHLH130基因在植物干旱脅迫響應過程中的功能,本研究構建了該基因的過表達載體(圖5A);通過根癌農桿菌介導的浸花法轉化,并在含有草銨膦的MS培養基上篩選獲得抗性苗(圖5B);進一步提取擬南芥野生型和抗性苗的總RNA,半定量PCR檢測結果顯示,外源基因GmbHLH130在擬南芥抗性苗中成功表達(圖5C),說明獲得的遺傳材料可靠,可用于后續試驗。
挑選同時期收獲的擬南芥種子,包括野生型(WT)、轉基因擬南芥T3代純合子(2個獨立株系L1和L2),均勻播種于正常MS培養基和20% PEG處理的MS培養基上。培養7 d后,觀察擬南芥的生長狀況,發現正常培養基上WT和轉基因擬南芥都能正常生長且子葉幾乎都能展開;而在20% PEG處理的MS培養基上野生型種子雖然能夠萌發,但是子葉幾乎無法正常展開,即受到嚴重的抑制,過表達株系L1和L2的綠色子葉率則顯著高于WT(圖6A、圖6B)。進一步將在正常MS培養基上萌發2 d的WT、過表達株系L1和L2移至MS培養基和20% PEG處理的培養基上,然后將培養基垂直放置培養7 d,結果顯示正常條件下上述材料的根長無明顯差異;而在20% PEG處理條件下,L1和L2的根長顯著大于WT(圖6C)。在盆栽條件下對比過表達GmbHLH130基因植株與WT的耐旱性,發現正常澆水條件下,生長表型無明顯差異;而自然干旱導致WT植株枯萎死亡,過表達植株生長雖然受到抑制,但仍然存活;恢復澆水后,WT植株無法復活,而過表達植株葉片變綠、能夠繼續生長(圖6D)。以上結果說明GmbHLH130基因過表達導致轉基因擬南芥植株耐旱性增強。
3 討論
研究結果表明,植物bHLH家族基因在植物非生物脅迫逆境響應方面發揮重要作用[4]。甘藍型油菜BnbHLH122基因在高溫、低溫、干旱、高鹽和滲透脅迫下均顯著上調,證明BnbHLH122基因在植株抵御逆境脅迫過程中發揮調節作用[19]。沈方圓等[20]利用全基因組和轉錄組聯合分析在歐洲山楊中鑒定出4個潛在的bHLH抗旱蛋白。本研究通過構建不同植物bHLH家族成員的系統進化樹,發現大豆GmbHLH130與擬南芥AtbHLH112、AtbHLH122、AtbHLH68及水稻OsbHLH068在同一分支,并且和AtbHLH122進化關系最近。對基因家族或同源基因進行系統進化分析時,一般認為屬于同一亞家族或同一進化分支的成員同源性較高,可能具有類似的生物學功能[21]。鑒于擬南芥AtbHLH122在增強植物耐旱性方面的功能[9],推測進化關系較近的大豆GmbHLH130在干旱脅迫響應和適應過程中可能發揮特定功能。本研究結果顯示,20%PEG模擬處理的大豆根和葉中GmbHLH130基因的表達水平都顯著上調,且在根中上調更明顯,但是隨著處理時間延長基因表達出現上下波動,這種干旱誘導表達特性可能與其對干旱脅迫的適應有關[1]。此外,克隆GmbHLH130基因啟動子,構建啟動子融合GUS報告基因的轉基因擬南芥PGmbHLH130::GUS,通過模擬干旱處理和GUS染色顯示啟動子對干旱脅迫的響應,這一結果也進一步證明GmbHLH130基因參與干旱脅迫調控過程。通常認為轉錄因子是核定位蛋白,在細胞核中結合到目標基因的特定順式作用元件調控基因表達[2]。本研究結果顯示,大豆GmbHLH130也是核定位轉錄因子,在Y2H酵母細胞中也能表現明顯的轉錄激活活性,表明GmbHLH130作為激活下游靶基因表達的轉錄因子發揮作用。
有報道稱外源基因在組成型啟動子控制下的表達會對轉基因擬南芥的正常生長、發育過程產生不利影響[22]。本研究通過農桿菌浸花轉化法獲得了GmbHLH130基因過表達擬南芥植株。比較擬南芥野生型WT和GmbHLH130過表達株系(L1和L2)在正常培養基和栽培基質上的生長表型,發現過表達植株和WT表型無明顯差異,說明大豆基因的異源表達未對擬南芥的生長發育產生不利影響。而在干旱條件下,所有材料的生長均受到抑制,綠色子葉率降低,但是過表達植株的綠色子葉率明顯高于WT;此外,模擬干旱條件下,過表達株系的根長也顯著大于WT;盆栽自然干旱條件下過表達植株也表現出較WT植株更強的耐旱性。研究結果證明,GmbHLH130基因過表達在不影響植株正常生長過程的前提下增強了擬南芥的耐旱性,這對大豆等作物耐旱性分子育種具有重要意義。本研究篩選到1個受干旱誘導表達上調的bHLH轉錄因子家族基因GmbHLH130,通過系統進化、亞細胞定位、轉錄激活活性以及干旱處理的轉基因擬南芥表型分析,初步證明了GmbHLH130可增強轉基因植株的耐旱性。但是轉錄因子GmbHLH130參與植物干旱脅迫響應和適應的分子機制仍需進一步研究。
參考文獻:
[1] LIANG Y F, MA F, LI B Y, et al. A bHLH transcription factor, SlbHLH96, promotes drought tolerance in tomato[J]. Horticulture Research, 2022, 9: 198.
[2] LIANG B B, WAN S G, MA Q L, et al. A novel bHLH transcription factor PtrbHLH66 from trifoliate orange positively regulates plant drought tolerance by mediating root growth and ROS scavenging[J]. International Journal of Molecular Sciences, 2022, 23(23): 15053.
[3] WANG X P, NIU Y L, ZHENG Y. Multiple functions of MYB transcription factors in abiotic stress responses[J]. International Journal of Molecular Sciences, 2021, 22(11): 6125.
[4] LIU Y J, JI X Y, NIE X G, et al. Arabidopsis AtbHLH112 regulates the expression of genes involved in abiotic stress tolerance by binding to their E-box and GCG-box motifs[J]. The New Phytologist, 2015, 207(3): 692-709.
[5] JOO H, BAEK W, LIM C W, et al. Post-translational modifications of bZIP transcription factors in abscisic acid signaling and drought responses[J]. Current Genomics, 2021, 22(1): 4-15.
[6] KHOSO M A, HUSSAIN A, RITONGA F N, et al. WRKY transcription factors (TFs): molecular switches to regulate drought, temperature, and salinity stresses in plants[J]. Frontiers in Plant Science, 2022, 13: 1039329.
[7] DEBBARMA J, SARKI Y N, SAIKIA B, et al. Ethylene response factor (ERF) family proteins in abiotic stresses and CRISPR-Cas9 genome editing of ERFs for multiple abiotic stress tolerance in crop plants: a review[J]. Molecular Biotechnology, 2019, 61(2): 153-172.
[8] BABITHA K C, RAMU S V, PRUTHVI V, et al. Co-expression of AtbHLH17 and AtWRKY28 confers resistance to abiotic stress in Arabidopsis[J]. Transgenic Research, 2013, 22(2): 327-341.
[9] LIU W W, TAI H H, LI SS, et al. bHLH122 is important for drought and osmotic stress resistance in Arabidopsis and in the repression of ABA catabolism[J]. The New phytologist, 2014, 201(4): 1192-1204.
[10]WANG F B, ZHU H, KONG W L, et al. The Antirrhinum AmDEL gene enhances flavonoids accumulation and salt and drought tolerance in transgenic Arabidopsis[J]. Planta, 2016, 244(1): 59-73.
[11]QIU J R, HUANG Z, XIANG X Y, et al. MfbHLH38, a Myrothamnus flabellifolia bHLH transcription factor, confers tolerance to drought and salinity stresses in Arabidopsis[J]. BMC Plant Biology, 2020, 20(1): 542.
[12]劉 福,陳 誠,張凱旋,等. 日本百脈根LjbHLH34基因克隆及耐旱功能鑒定[J]. 草業學報, 2023, 32(1): 178-191.
[13]VALLIYODAN B, YE H, SONG L, et al. Genetic diversity and genomic strategies for improving drought and waterlogging tolerance in soybeans[J]. Journal of Experimental Botany, 2017, 68(8): 1835-1849.
[14]李文濱,宋春曉,萇興超,等.干旱脅迫下20個大豆品種抗旱性評價[J].東北農業大學學報, 2019, 50(4): 1-10.
[15]XU Z L, LIU X Q, HE X L, et al. The soybean basic Helix-Loop-Helix transcription factor ORG3-like enhances cadmium tolerance via increased iron and reduced cadmium uptake and transport from roots to shoots[J]. Frontiers in Plant Science, 2017, 8:1098.
[16]CHENG Q, DONG L D, GAO T J, et al. The bHLH transcription factor GmPIB1 facilitates resistance to Phytophthora sojae in Glycine max[J]. Journal of Experimental Botany, 2018, 69(10): 2527-2541.
[17]LI L, GAO W W, PENG Q, et al. Two soybean bHLH factors regulate response to iron deficiency[J]. Journal of Integrative Plant Biology, 2018, 60(7): 608-622.
[18]KU Y S, NI M, MUNOZ N B, et al. ABAS1 from soybean is a 1R-subtype MYB transcriptional repressor that enhances ABA sensitivity[J]. Journal of Experimental Botany, 2020, 71(10): 2970-2981.
[19]唐佳佳,萬云寶,王茂林.甘藍型油菜BnbHLH122基因的克隆、表達模式及脅迫響應分析[J]. 四川大學學報(自然科學版), 2021, 58(2): 180-188.
[20]沈方圓,王嵐春,李 校. 歐洲山楊bHLH轉錄因子家族全基因組分析[J]. 四川大學學報(自然科學版), 2021, 58(3): 179-187.
[21]CHEN Y, ZHU P P, WU S Y. Identification and expression analysis of GRAS transcription factors in the wild relative of sweet potato Ipomoea trifida[J]. BMC Genomics, 2019, 20(1): 911.
[22]LI P, ZHANG B, SU T, et al. BrLAS, a GRAS transcription factor from Brassica rapa, is involved in drought stress tolerance in transgenic Arabidopsis[J]. Frontiers in Plant Science, 2018, 9:1792.
(責任編輯:陳海霞)
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
