沿降雨梯度的檸條錦雞兒種子萌發的跨代可塑性研究
2023-12-04 06:48:14袁家梁趙麗敏李秧秧
草地學報 2023年11期
關鍵詞:模型
袁家梁, 趙麗敏, 李秧秧
(1.西北農林科技大學林學院, 陜西 楊凌 712100; 2.內蒙古卓資縣林業和草原局, 內蒙古 卓資 012300; 3.西北農林科技大學水土保持研究所, 黃土高原土壤侵蝕與旱地農業國家重點實驗室, 陜西 楊凌 712100)
跨代可塑性(Transgenerational plasticity)是指當父母代所經歷的環境影響他們后代的表型[1]。跨代可塑性可能增加或降低后代的適應性,適應性跨代可塑性可能在植物適應不同類型壓力和環境中起著十分重要的作用[2-3]。目前關于跨代可塑性的研究主要集中在環境信號(高溫、干旱、土壤壓實、鹽與養分脅迫、重金屬、高CO2、病蟲害等)與跨代可塑性強弱的關系、跨代可塑性的生態和進化學意義及跨代可塑性產生的機制等方面[1-2]。
干旱是影響植物跨代可塑性的一個重要環境信號,關于干旱對植物跨代可塑性的影響已有較多研究,但大多數研究集中在植物苗期,而對于種子萌發期的跨代可塑性研究相對較少。種子萌發是植物生活史的一個重要階段,決定著植物的建成和幼苗的生長存活。水分是影響種子萌發的關鍵因素,關于種子萌發對干旱的敏感性可通過水勢模型(Hydrotime model)來度量[4-5]。水勢模型涉及種群萌發的閾值水勢(ψb(50),MPa)、水勢常數(θH,MPa d)和標準差(σψb,MPa)三個參數,其中ψb(50)是種子萌發的最低水勢,表征了種子萌發過程對干旱的敏感性;θH是種子為萌發需要積累的高于閾值水勢的MPa d,反映了種子萌發的快慢;σψb表示種子萌發的整齊度[5]。該模型已經被廣泛用來研究雜草、草本植物、牧草、灌木等植物的萌發特性[6]。
種子萌發期的跨代可塑性與物種和親本遭受的環境梯度有關。智利的一種野生西紅柿種子和美國的長芒莧(Amaranthuspalmeri),當母系遭受水分虧缺時,其后代能在低的水勢下萌發[7-8]。澳大利亞的一種入侵植物美洲海灘芥(Cakileedentula),其在亞熱帶的萌發水勢閾值ψb(50)亦高于溫帶[9]。Zhang等[10]比較了低溫多雨的冷涼生境和高溫干旱的溫暖生境不同針茅屬植物種子萌發的水勢模型參數,發現兩種生境針茅屬植物間的水勢模型參數無差異。南美巴塔哥尼亞高原北部一種高羊茅(Festucapallescens)沿降雨梯度上9個種群的閾值水勢ψb(50)并無顯著差異,但水勢常數θH不同,在分布梯度的兩端種群具有高的水勢常數θH,意味著在不是很適宜條件下種子萌發可能被阻止或延遲[11]。
檸條(CaraganakorshinskiiKom.)是我國北方干旱半干旱地區的主要多年生灌木,因其耐旱、耐寒、耐貧瘠和耐風沙,有極強的生命力和適應性,且生物量高、營養豐富、適口性好,被廣泛用做牲畜的優質飼草飼料。關于其種子萌發特性已有一些研究[12-13],但其種子萌發過程是否具有跨代可塑性并不清楚。研究檸條種子萌發過程的跨代可塑性對于揭示檸條對干旱的適應性機制和檸條合理引種有一定的指導價值。為此,我們采集了不同降雨梯度上的檸條種子,利用水勢模型研究了其種子萌發過程對干旱的敏感性,試圖說明檸條在萌發過程中是否產生適應性跨代可塑性。假設:不同種源種子對水分有效性的響應不同,干旱地區來源種子不僅具有更低的萌發水勢閾值,且具有高的標準差,從而使部分種子進入休眠而在水分來臨時萌發,以適應干旱地區的多變低水環境。
1 材料與方法
1.1 種子收集及萌發試驗
沿降雨梯度從黃土高原、鄂爾多斯高原及青藏高原四個采樣點采集檸條成熟種子,四個采樣點分別位于陜西省安塞縣、陜西省神木市、內蒙古涼城縣、青海省共和縣,其多年平均降雨量從549 mm下降至246 mm。于2021年7—8月種子成熟時進行隨機采收,采樣地基本情況及種子特征詳見表1。采集后的種子放置室溫儲藏8個月供萌發試驗所用。
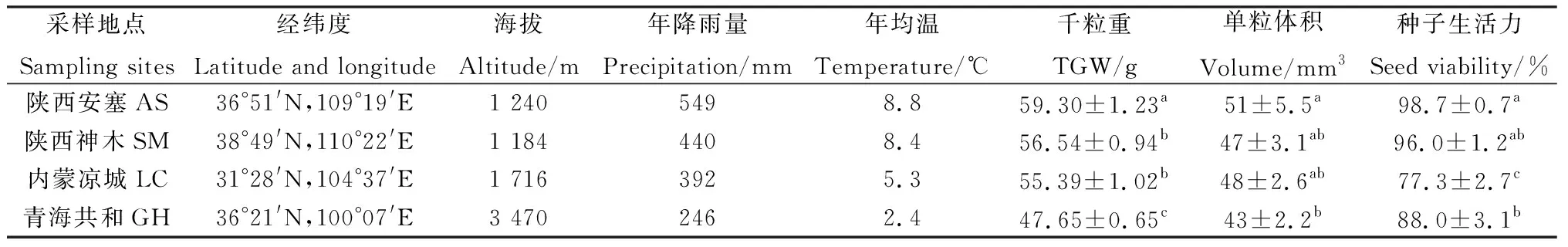
表1 采樣地及種子基本特征Table 1 Basic traits of seed-collecting sites and seeds
為防止種子霉變,試驗前使用1%次氯酸鈉溶液對種子表面消毒1.5 min后用蒸餾水反復沖洗。由于種子萌發時的溫度會影響其水時間模型參數,且檸條種子萌發的最適溫度為15℃~20℃[13]和30℃[15],故本研究在3個溫度梯度(20℃、25℃、30℃)和5個水勢梯度(0,-0.25,-0.4,-0.6,-0.8 MPa)下進行種子萌發試驗。不同水勢梯度溶液使用PEG6000配制[16],所有PEG溶液在使用前用蒸汽壓滲透計(VAPRO 5520,Wescor,Logan,USA)進行校正。每個水勢梯度4個重復,每個重復25粒種子。種子被放置在9 cm直徑的培養皿中,以雙層濾紙作為萌發基質,使用7 ml不同濃度PEG溶液潤濕濾紙,為了防止霉變及在發芽試驗期間保持培養基水勢恒定,每48小時更換一次濾紙及PEG溶液,培養皿使用封口膜進行密封,以防止水分流失。萌發實驗在黑暗的培養箱中進行,當胚根突破種皮2 mm時認為已經萌發,將其取出并計數,監測頻率視處理而定。當連續至少7天未觀測到種子萌發時,終止試驗。試驗分三批次(溫度重復)進行。種子累積發芽率為萌發試驗結束后累積萌發的種子數占所用種子總數的百分比。萌發速率(t50)為達到最大發芽率的50%所需的時間。
1.2 水勢模型參數計算
為進一步了解水勢降低對種子萌發的影響,Bradford[4]在Gummerson[17]工作的基礎上提出了水勢模型:
θH=[ψ-ψb(g)]tg
(1)
式中ψ表示種子所處環境水勢(MPa),ψb(g)表示水勢閾值(MPa),即給定種子萌發百分比g所能夠萌發的最低水勢,tg為達到萌發百分比g所需要的時間(d)。θH(MPad)為水勢常數,表示種子吸脹到胚根出現所需的累積水勢時間,通常認為θH是一個常數。將公式1進行簡單轉換,可以得到公式2:
(2)
公式2表明1/tg是ψ的線性函數,其斜率為1/θH,截距為ψb(g)。因為θH為常數,所以對于任意g其斜率相等,ψb(g)隨著g變化,對于一般物種而言,ψb(g)在種群的分布為正態分布。為了對不同g下的ψb(g)進行估計,將公式1轉換得到公式3:
ψb(g)=ψ-θH/tg
(3)
將觀測的發芽時間數據在概率尺度上線性化并對ψb(g)進行回歸,可以得到下述公式4:
(4)
自此,便可獲得水勢模型的三個參數:θH、ψb(50)及σψb。
1.3 數據分析
使用雙因素方差分析比較不同種源、萌發溫度及其相互作用對檸條種子萌發特征的影響,若差異顯著使用LSD法進行多重比較。種子萌發曲線使用drcSeedGerm包中的時間-事件模型進行擬合,使用drmte函數分別對每一溫度下的水勢進行參數擬合[18]。所有數據處理均在R(ver. 4.2.0)中進行。
2 結果與分析
2.1 不同檸條種源沿降雨梯度的種子萌發特征
沿從南到北降雨梯度,檸條種子千粒重、單粒體積及種子生活力均呈現下降趨勢,內蒙涼城檸條種子的千粒重、單粒體積及種子生活力為陜西安塞檸條種子的93.4%,94.1%和78.3%,其千粒重和種子生活力顯著下降(P<0.05);青海共和檸條種子的千粒重、單粒體積及種子生活力僅為陜西安塞檸條種子的80.3%,84.3%和89.1%,三者均顯著低于陜西安塞種子(P<0.05)(表1)。
20℃下無水分脅迫時,陜西安塞檸條種子的累積發芽率為93%,而青海共和的檸條種子累積發芽率僅為77%,30℃下陜西安塞和青海共和種子累積發芽率則分別為73%和56%(圖1),進一步證明了檸條種子質量隨從南到北降雨梯度上的降低。隨種子萌發過程中水分脅迫程度增加,4個種源檸條種子累積發芽率明顯降低,且萌發速率(t50)變慢(圖1)。20℃、25℃和30℃下,4個種源的平均累積發芽率分別為58%±3%、74%±1%和32%±4%,且萌發速率分別為3.52±0.23 d、3.52±0.19 d及4.99±0.37 d(圖1),表明25℃是檸條種子萌發的最適溫度。
2.2 種源和溫度對檸條種子萌發的水勢模型參數的影響
種源對水勢常數θH有顯著影響(P<0.05),對閾值水勢ψb(50)的影響接近顯著(P=0.062),但對標準差σψb無顯著影響。溫度極顯著影響水勢模型的3個參數(P<0.01),但種源和溫度的交互作用對水勢模型的3個參數無顯著影響(表2)。溫度對水勢模型3個參數的影響大于種源的影響。

表2 種源和溫度對水勢模型影響的雙因素方差分析Table 2 Two-way ANOVA analysis of seed sources and temperature on the hydrotime model parameters
陜西安塞的水勢常數θH顯著低于其他3個地點,陜西安塞和神木的水勢閾值ψb(50)表現出低于內蒙涼城和青海共和的趨勢,且安塞和神木的水勢閾值ψb(50)顯著低于青海共和,4個種源的標準差σψ b無顯著差異(圖2)。隨溫度增加,水勢常數θH顯著降低,30℃下的θH顯著低于20℃(P<0.05);水勢閾值ψb(50)則為先降低后升高,三個溫度下均有顯著差異(P<0.05);標準差σψb也表現出先降低后升高,30℃下的σψ b顯著高于其他兩個溫度(圖3)。在種子萌發的最適溫度(25℃),其萌發的水勢閾值ψb(50)和標準差σψ b最低。
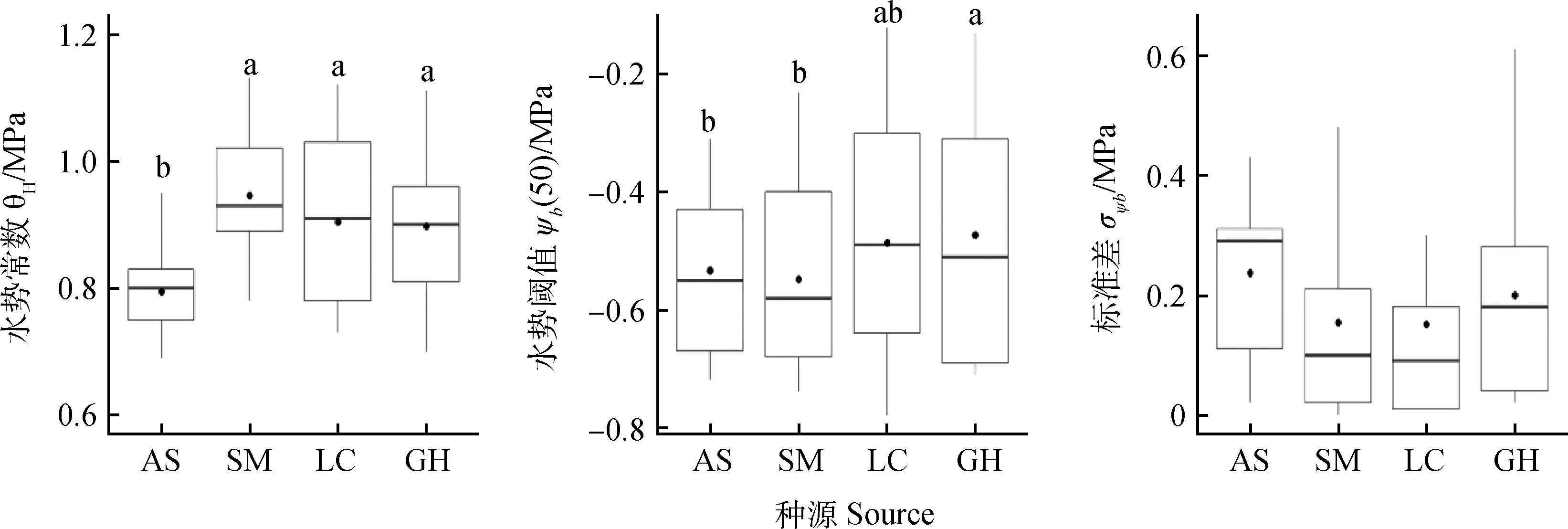
圖2 不同種源檸條種子萌發的水時間模型常數Fig.2 Hydrotime model parameters for the 4 seed sources of C. korshinskii注:不同小寫字母表示種源間差異顯著Note:Different lowercase letters indicate a significant difference among different seed sources at P<0.05
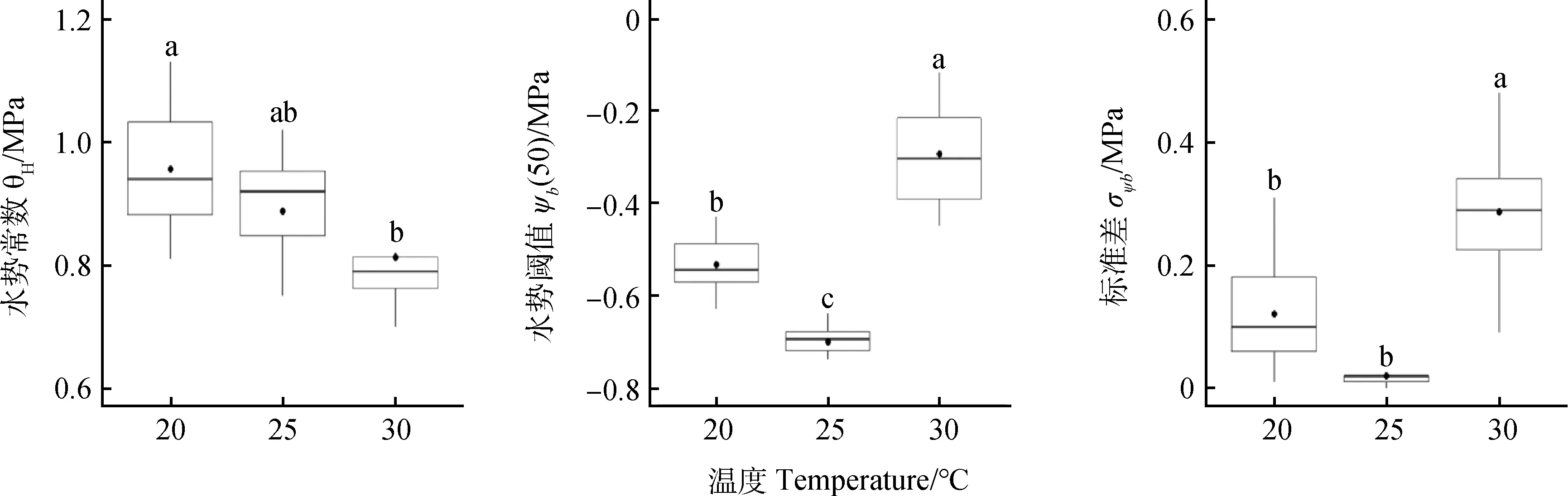
圖3 不同萌發溫度下檸條種子萌發的水時間模型參數Fig.3 Hydrotime model parameters for Caragana korshinskii seeds under different germination temperatures注:小寫字母表示不同溫度間的差異顯著(P<0.05)Note:Different lowercase letters indicate a significant difference between different temperatures at P<0.05
2.3 水勢模型參數與種子質量、種源地氣象因子間的相關關系
水勢模型參數和種子特征,如種子千粒重、種子單粒體積、種子生活力間并無顯著相關性,與年降雨量間亦無顯著相關性,但與年均溫存在接近顯著的負相關關系(P=0.052)(表3,圖4),年均溫度越高,種子萌發的水勢閾值越低,種子萌發過程越耐旱。
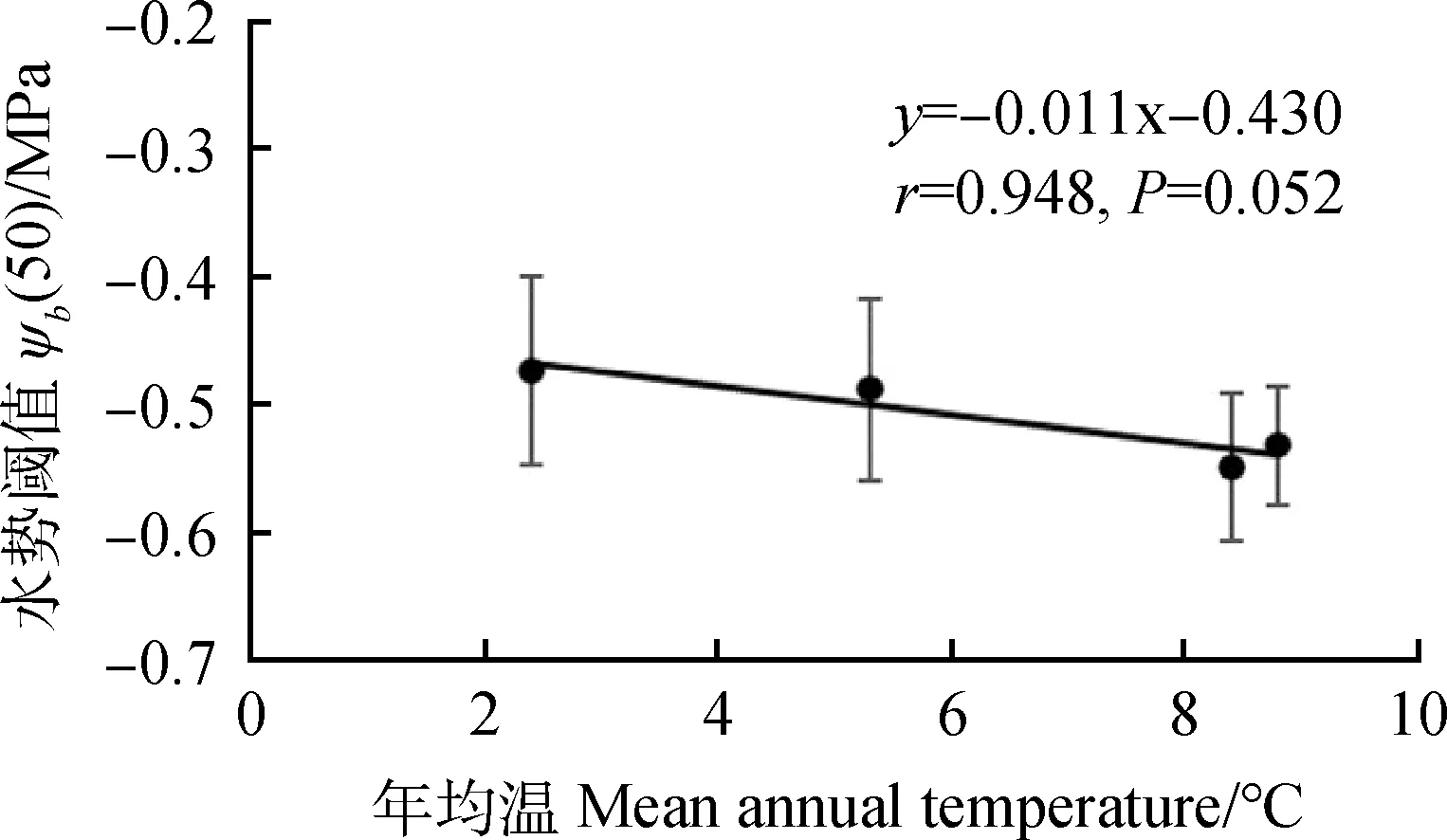
圖4 水勢閾值ψb(50)與年均溫間相關關系Fig.4 Pearson correlation between average annual temperature and water potential threshold ψb(50)

表3 水勢模型參數與種子性狀及氣候條件的Pearson相關性Table 3 Pearson correlation between hydrotime model parameters and seed traits,climate variables
3 討論
3.1 降雨梯度上檸條種源對水勢模型參數的影響
本研究中,陜西安塞種源的水勢常數θH顯著低于其他3個地點,陜西安塞和神木的水勢閾值ψb(50)顯著低于青海共和,4個種源的標準差σψb無顯著差異(圖2)。López等[11]研究了巴塔哥尼亞高原北部一種高羊茅9個種群沿降雨梯度上種子萌發的水勢模型參數變化,發現在分布梯度的兩端種群具有高的水勢常數θH,且種群間ψb(50)和σψb無顯著差異,他們認為微環境對植物生理行為的影響更大,而宏觀降雨尺度的影響相對較小。本研究中,水勢常數θH和年均降雨量及年均溫度的相關性并不顯著(表3),說明安塞種子萌發的水勢常數低主要可能與微環境有關,安塞種子通過快速萌發來適應其立地條件。
種子萌發過程中的水勢閾值ψb(50)反映了種子萌發過程對干旱的敏感性。Lai等[19]研究認為內蒙古高原檸條錦雞兒、中間錦雞兒與小葉錦雞種子萌發的耐旱程度隨其生境降雨量的增加而降低。澳大利亞的一種入侵植物美洲海灘芥,其在亞熱帶的萌發水勢閾值ψb(50)亦高于溫帶[9],Zhang等[10]比較了低溫多雨的冷涼生境和高溫干旱的溫暖生境不同針茅屬植物種子萌發的水勢模型參數,發現兩種生境針茅屬植物間的水勢模型參數無差異。但在本研究中,年降雨量高的陜西安塞和神木種子萌發的水勢閾值ψb(50)顯著低于干旱的青海共和。不同植物種子萌發對水分的響應并不相同,這可能與其生境對種子萌發的選擇壓力有關。隨降雨量降低,檸條的繁殖方式從有性繁殖向無性繁殖轉變[20],可能不需要通過降低種子萌發過程的水勢閾值ψb(50)來增加萌發過程中的抗旱性,因為相比于無性繁殖,跨代產生可塑性的成本可能更高[1,21]。我們的結果并不支持干旱地區具有更低的萌發水勢閾值這一假設,這與關于動植物跨代可塑性的薈萃分析表明:一年生植物表現出更強的跨代可塑性,而多年生植物幾乎沒有跨代可塑性[22]一致。干旱地區的檸條也并未通過種子萌發過程中高的標準差來適應其多變低水環境。
3.2 萌發溫度對水勢模型參數的影響
隨萌發溫度從20℃增加到25℃,水勢閾值ψb(50)顯著降低,但當萌發溫度從25℃增加至30℃,水勢閾值ψb(50)和標準差σψb顯著增加,30℃的水勢常數θH顯著低于20℃(圖3)。因此,相對于最適萌發溫度,溫度降低或升高均導致水勢閾值ψb(50)增大。這與在紅花[23]、狗尾草屬物種[24]、馬鈴薯[25]、埃塞俄比亞芥[26]、鷹嘴豆[27]、波斯甜瓜[28]等的研究結果一致。低溫下較高的水勢常數θH與水勢閾值ψb(50)可能主要與低溫延遲了種子萌發過程有關。低溫導致種子充分吸水滯后,同時降低了種子的代謝活力,使種子無法在低水勢下突破種皮,從而延遲或阻止了胚根伸出。種子在較低溫度下對干旱脅迫的耐受力低,也為其在早春較低的溫度條件下的種子萌發提供了保障,這些幸存于土壤種子庫的休眠種子一旦遇有降水即可開始萌發,種子在春季低溫期間的提早萌發可保證幼苗根系充分發育的時間,增強幼苗對夏季降雨間隔期內強烈干旱的耐受能力,進而增大幼苗存活和建立的機會。溫度增加(30℃)通過降低萌發過程的水勢常數θH,從而加快種子的萌發速率。較高的水勢閾值ψb(50)表明檸條種子萌發在高溫下對干旱脅迫的敏感性增加,遇到少量水即可萌發,從而提高了其在夏季干熱生境的適合度,是其在溫度相對較高的環境中成功定居和種群更新的重要保障。最適溫度25℃下,檸條種子的水勢閾值ψb(50)和標準差σψb最低,表明在該溫度下檸條種子萌發最為耐旱且萌發最為整齊,溫度降低或者增加均導致種子萌發不整齊。
3.3 水勢模型參數與種子性狀和氣候條件的關系
本研究中,盡管在從南到北的降雨梯度上,檸條種子質量顯著降低,但種子萌發的水勢模型參數與種子質量性狀(千粒重、單粒體積和種子生活力)均無顯著相關性(表3),意味著種子性狀不是影響檸條不同種源水勢模型參數的主要因素。對新熱帶森林的演替早期種而言,隨種子質量增加,水勢閾值ψb(50)降低,意味著大種子萌發過程的抗旱性更強,因而更易生長在光照和溫度高的大林窗中[29]。Arene等[30]的Meta分析發現對一年生植物,種子質量和臨界水勢ψb(50)呈負相關,即種子質量越大,臨界水勢越低,而對多年生植物,種子質量和臨界水勢ψb(50)無關。多年生植物可能依賴于相對長的壽命和適應性(如存儲器官、大的根系、休眠芽或極端抗旱性),使其能在不利季節存活[31],這些適應性降低了種子萌發階段的相對重要性,所以導致種子大小和臨界水勢無關。檸條為多年生植物,其相對強的抗旱性[32-33]和在降雨量低的地區主要依賴于無性繁殖的繁殖方式可能是其種子質量和臨界水勢無相關性的重要原因。
水勢模型參數中,僅水勢閾值ψb(50)和年均溫呈接近顯著負相關,年均溫越高,閾值水勢ψb(50)越低。袁禎等[15]也發現分布在半濕潤地區的紅花錦雞兒萌發最耐旱,而分布在干旱荒漠區的多刺錦雞兒、荒漠錦雞兒和粉刺錦雞兒其種子萌發對水勢脅迫最為敏感,與本結果相似,但與物種間發現的萌發基礎溫度越低,萌發的水勢閾值越低不一致[6]。在降雨量少、年均溫低的地區,種子萌發后幼苗極易遭遇干旱而導致閃苗,低的年均溫導致萌發的水勢閾值增加,從而使萌發發生在濕潤條件下,因而高水勢閾值是一個風險降低的特征,可能是這類地區檸條種子萌發過程的重要適應性對策。年均溫較高的安塞和神木盡管萌發溫度高可能增加萌發水勢閾值,但其種子在7月成熟,此時蒸發強烈,因而降低水勢閾值ψb(50)從而加快種子萌發可能是其適應性選擇結果,萌發時水勢閾值與萌發季節水分條件的關系可能更大,此外,土壤和立地條件等也可能影響水勢閾值,對此值得更進一步研究。
4 結論
沿降雨梯度上種源對種子萌發的水勢常數θH和水勢閾值ψb(50)均有影響。降雨量高的陜西安塞種源的水勢常數θH顯著低于其他3個種源,且其水勢閾值ψb(50)顯著低于干旱的青海共和種源。干旱地區檸條種子并不具有更低的萌發水勢閾值和高的標準差,表明沿降雨梯度上不同種源檸條種子萌發過程中并不具有適應性跨代可塑性。不同種源檸條種子萌發的水勢閾值ψb(50)主要決定于年均溫而非種子性狀,年均溫相對高地區的檸條種子(陜西安塞和神木)主要通過降低水勢閾值ψb(50)從而加快種子萌發來適應其生境,而年均溫低的地區(青海共和)則通過增加水勢閾值來適應其生境。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
